











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
El transporte de las fracciones inorgánicas disueltas del nitrógeno (NID: NO-2 + NO-3 + NH+4) y el fósforo (PID: ortofosfatos) del reservorio profundo a la zona eufótica, es quizás el principal factor modulador de la producción primaria para la mayoría de los ecosistemas marinos (Tyrrell, 1999; Deutsch & Weber, 2012).
En los estratos superficiales y subsuperficiales, estos nutrientes tienden generalmente a consumirse por la generación de biomasa por parte de los organismos fotosintéticos (e.g., Eppley & Renger, 1986). Una vez en forma de materia orgánica, parte de ésta es transferida a través de las redes tróficas, mientras que la porción restante es exportada al fondo marino donde se recicla en la columna de agua o queda secuestrada en el sedimento (Sarmiento & Gruber, 2006). De acuerdo al cociente de Redfield, la proporción con la cual estos elementos se fijan vía fotosíntesis y posteriormente se remineralizan de la materia orgánica particulada, mantiene un valor promedio (106:16:1 para C:N:P, respectivamente) relativamente similar a la estequiometría de las aguas profundas del océano moderno (Redfield, 1958).
No obstante, la gran variabilidad de este cociente entre regiones e incluso entre estratos de la columna de agua, surge como resultado del balance entre las rutas dominantes de ingreso y remoción de nitrógeno a los océanos (Codispoti, 1989). La fuente principal de estos flujos radica en la gran diversidad fisiológica de los microorganismos marinos, cuya intensa actividad depende substancialmente de los requerimientos metabólicos y las condiciones oceanográficas (Azam & Malfatti, 2006; Ulloa et al., 2012).
En la columna de agua de la Cuenca Alfonso, Bahía de la Paz, los cambios en el contenido de NID, PID y otros micronutrientes, ocurren principalmente por la incidencia estacional de mecanismos de fertilización asociados a la dinámica física del océano (e.g., estabilidad de la columna de agua, ingreso de masas de agua y procesos de submesoescala), así como por la presencia de fenómenos atmosféricos (transporte eólico, tormentas y huracanes) y la captación por parte de diversos grupos de microorganismos (Villegas-Aguilera, 2009; Silverberg et al., 2007; Martínez-López et al., 2012). Sin embargo, como en otras regiones del Golfo de California y el Pacífico Oriental Subtropical, en el área de la cuenca se presentan fuertes desviaciones en los cocientes NID:PID (<<16) respecto a la estequiometría elemental de Redfield (Villegas-Aguilera, 2009).
Un estudio sobre el flujo y la composición isotópica del nitrógeno en la materia orgánica particulada en trampas de sedimentación en Cuenca Alfonso, sugiere la incidencia y estacionalidad de los principales procesos que regulan positiva (fijación biológica de nitrógeno) y negativamente (desnitrificación) el inventario del nitrógeno, así como un predominio de esta última ruta (Aguíñiga et al., 2010). A pesar de que ninguno de estos procesos han sido medidos directamente, las condiciones deficientes de oxígeno (<1.4 ml l-1) en estratos por debajo de los ~50 m de profundidad (Monreal et al., 2001), sugieren que el desbalance de NID podría ser resultado de estos procesos de remoción. Sin embargo, no se descarta la posibilidad de que el desbalance en los cocientes N:P sean además consecuencia de un superávit en fósforo debido a su descomposición preferencial en la materia orgánica (Vitousek & Howarth 1991; Vitousek et al., 1997; Monteiro & Follows, 2012), al aporte continental y la naturaleza intrínseca del componente geológico de la zona (Daesslé et al., 2004; Ruttenberg, 2003), la intervención antropogénica de los reservorios sedimentarios (Correl, 1998), o a la advección de masas de aguas con características particulares de su región de origen (Codispoti & Richards, 1976; Rafter et al., 2012).
A falta de datos experimentales que cuantifiquen las vías de ingreso o pérdida de nitrógeno al océano, la aplicación de trazadores biogeoquímicos, tales como el N*, ha sido una herramienta relativamente útil y sencilla para conocer desviaciones significativas del comportamiento conservativo de los nutrientes de importancia biológica en diferentes escalas espaciales y temporales. En la literatura, la mayoría de las estimaciones del N* se han realizado para hacer balances a gran escala, empleando constantes deducidas a partir de los promedios globales y/o regionales de NID y PID obtenidos de las bases datos (e.g., GEOCECS, 1978; World Ocean Atlas, García et al., 2013; HOT, Karl & Lukas, 1996; Gruber & Sarmiento, 1997; Castro et al., 2001; Deutsch et al., 2001). En el presente trabajo, proponemos el uso de este trazador, adaptando algunas de las constantes de la ecuación propuesta por Gruber & Sarmiento (1997) a los valores de nutrientes en el agua de mar compilados durante el monitoreo de la Estación-Cuenca Alfonso (24° 29' N, 110° 36' W) entre diciembre de 2005 - diciembre de 2012. De esta manera, por primera vez se examina la variabilidad estacional del trazador cuasi-conservativo N* (Gruber & Sarmiento, 1997) en el sitio de Cuenca Alfonso entre febrero de 2012 y marzo de 2013, con el propósito de identificar desde una perspectiva local si hay déficit de nitrógeno y, con ello, la incidencia de los procesos dominantes de remoción e ingreso que son deducidos a partir de la proporción entre el NID y PID (Broecker & Peng, 1982; Gruber, 2008). Los resultados obtenidos proporcionan bases para comprender la importancia de algunas de las rutas que retroalimentan el inventario del nitrógeno y su relación con el ciclo de producción estacional, así como su vinculación con la dinámica física del sitio.
Área de estudio. La Cuenca Alfonso se encuentra ubicada en la zona norte de la Bahía de La Paz, frente a la costa suroriental de la Península de Baja California (Fig. 1). La región presenta un sistema de vientos estacionales paralelos a la costa, generalmente más intensos del noroeste en invierno, que se revierten con procedencia sureste en verano (Pares-Sierra et al., 2003). Durante la temporada fría, se presenta conjuntamente la influencia de los vientos locales o brizas, los cuales cambian de dirección y velocidad en el transcurso del día en consistencia con los gradientes de presión atmosférica entre el mar y el continente (Zaytsev et al., 2010). En el verano y parte del otoño, el clima deja de tener condiciones predominantemente secas, debido a que la incidencia de altas temperaturas superficiales del mar y del continente favorecen el arribo de humedad y la llegada de ciclones y tormentas tropicales que traen el mayor volumen de precipitación (>~80 %) para la zona (CONAGUA, 2013).
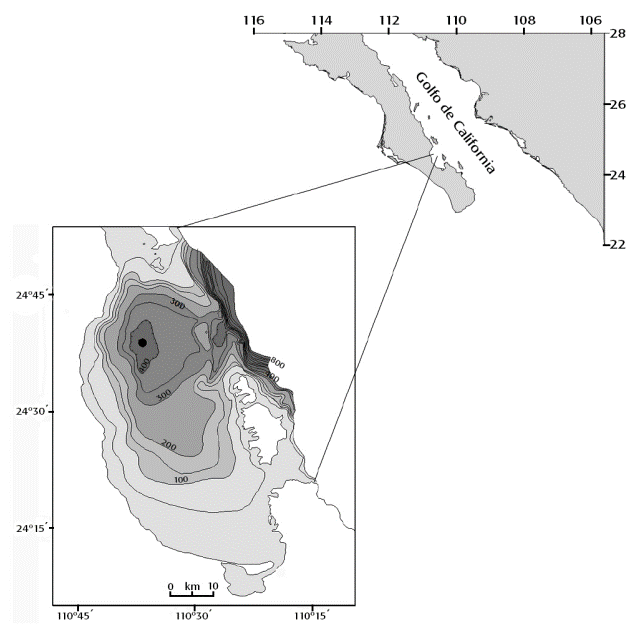
Figura 1 Ubicación de la Cuenca Alfonso, Bahía de La Paz, Golfo de California, México. La figura muestra la batimetría de la zona (Nava-Sánchez et al., 2001) y la posición de la Estación-Cuenca Alfonso (círculo negro).
Un rasgo importante en la columna de agua de esta zona, es la existencia de una capa deficiente de oxígeno (<1.4 ml l-1) en profundidades mayores a 50 m (Monreal et al., 2001), la cual es aparentemente una consecuencia de los altos niveles de producción primaria (Villegas-Aguilera, 2009) y la subsecuente oxidación de materia orgánica, así como por las características topográficas (profundidad máxima de 415 m; umbral a 275 m) que dificultan la ventilación en el fondo de la cuenca (Nava-Sánchez et al., 2001).
Materiales y métodos
El monitoreo en la cuenca se llevó a cabo mensualmente entre febrero de 2012 y marzo de 2013, exceptuando enero de 2013. La caracterización termohalina de los primeros 300 m de la columna de agua se efectuó mediante lances de CTD (SBE 19 Seacat profiler, SEABIRD). La recolección de muestras se realizó en ocho profundidades (cuatro en los niveles de 100, 33, 3 y 0.1% de la luz incidente en superficie, I0 de la zona eufótica, y las cuatro restantes a 150, 200, 250 y 300 m con una botella Niskin (modelo 1010, General Oceanics).
A partir de los datos de CTD se calculó la profundidad de la capa de mezcla (PCM) con base en el valor de la capa isotérmica (Δθ) de 0.5 °C (Kara et al., 2000). Los nutrientes analizados para este estudio fueron PID como ortofosfatos y NID, correspondiente a la sumatoria de las fracciones disueltas de nitritos, nitratos y amonio. Las muestras de agua fueron prefiltradas a través de filtros precombustionados GF/F y almacenadas en congelación a -20°C hasta su posterior análisis. La determinación de amonio se realizó el mismo día de la colecta en el Laboratorio de Fitoplancton del CICIMAR - IPN, de acuerdo a la técnica colorimétrica de Solórzano (1969), utilizando un espectrofotómetro Perkin Elmer modelo LAMBDA 25 UV/Vis. El resto de los nutrientes se analizaron según las técnicas colorimétricas propuestas por Strickland & Parsons (1972) en el Laboratorio de Eutrofización Costera del CIIDIR - IPN, Unidad Sinaloa, utilizando un espectrofotómetro UV/Vis Jenway, modelo 6705.
A partir de la distribución espacio-temporal de las concentraciones de PID y NID se estimó el trazador biogeoquímico N* de acuerdo a la ecuación de Gruber & Sarmiento (1997):
N* = (NID - (ΔN/ΔP × PID) + Kte1) × Kte2
En donde: ΔN/ΔP (13.16), corresponde al cociente NID:PID, resultado de los procesos de fotosíntesis y de remineralización de la materia orgánica, que fue ponderado a partir de las concentraciones de nutrientes compiladas de los datos históricos para la Cuenca Alfonso (Fig. 2); Kte1 (9.40), es una constante que representa el exceso de PID en relación a la covariación ideal entre el valor promedio de PID (1.81 μmol kg-1) y el de NID (14.41 μmol kg-1), y que se modificó de la ecuación original de acuerdo a los valores definidos a partir de los datos históricos compilados para la Cuenca Alfonso (Fig. 2); y Kte2 (0.87), es la constante que indica la relación molar entre el nitrato utilizado para oxidar, bajo condiciones disóxicas, la materia orgánica y el nitrógeno molecular (N2) producido, según el balance estequiométrico de la ecuación química de la desnitrificación propuesta por Anderson (1995).
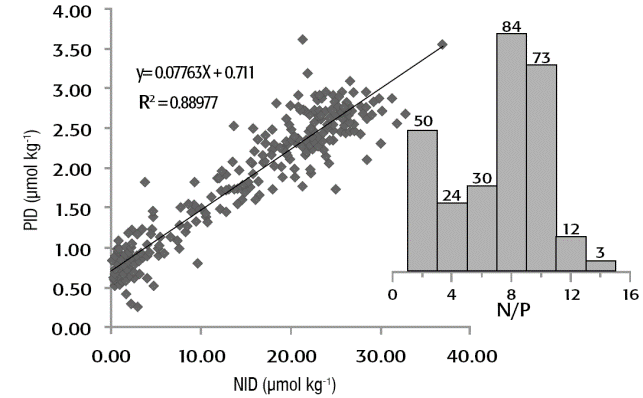
Figura 2 Datos de concentración de fósforo inorgánico disuelto (PID) versus nitrógeno inorgánico disuelto (NID) de la Bahía de La Paz, México entre 2005-2012 (Martínez-López, datos no publicados). El inverso de la pendiente de la ecuación de la regresión lineal corresponde al cociente ΔN/ΔP, y el punto de corte a la abscisa y indica el PID remanente en ausencia de NID. Las barras representan el histograma de frecuencia de los valores del cociente N/P.
Resultados
PCM. El rango de profundidades para la capa de mezcla fluctuó entre ~11 y 97 m, con valores menores entre abril y octubre (~11 - 27 m) y máximos durante los primeros meses del 2013 (~96 - 97 m; Fig. 3).
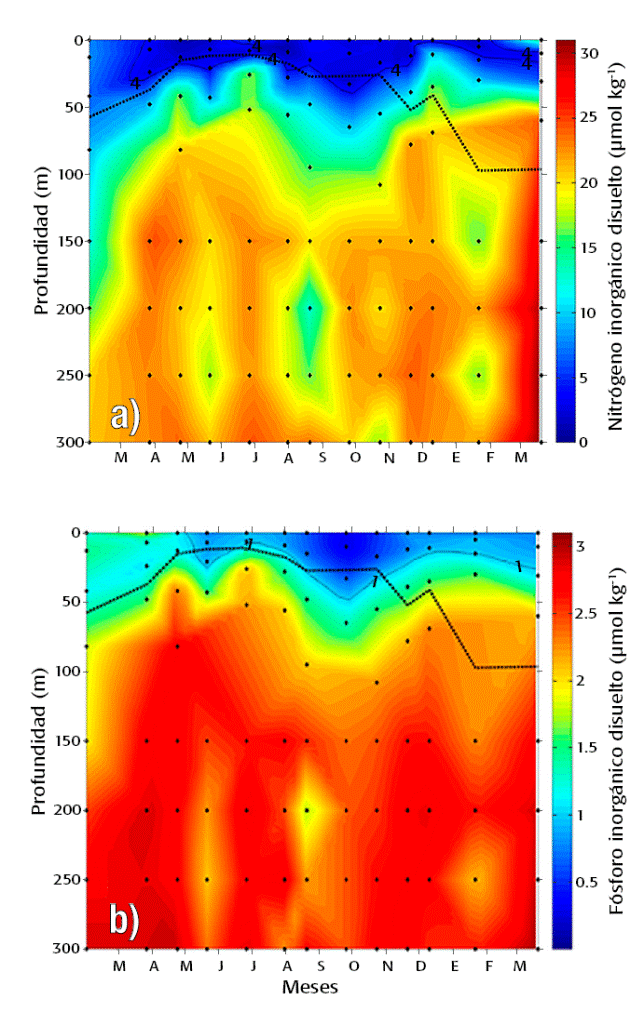
Figuras 3a-b Distribución temporal y vertical de concentraciones de NID (a) y PID (b) en los primeros 300 m de profundidad de la Cuenca Alfonso, Bahía de La Paz, México. Las líneas continuas en color negro representan las isolíneas de concentración de 4 y 1 μmol kg-1 para NID y PID, respectivamente. La línea discontinua representa la profundidad de la capa de mezcla (PCM).
Distribución del NID y PID. Ambos nutrientes presentaron una distribución similar en el tiempo y en la columna de agua de la Cuenca Alfonso (Fig. 3a-b ). En general, los datos muestran una tendencia a incrementar las concentraciones con la profundidad, así como dos incrementos en subsuperficie, debidos a la activación de un remolino ciclónico entre abril - julio y entre noviembre - diciembre como resultado de la mezcla intensa dada por vientos del Noroeste.
En la zona eufótica (100 - 0.1% I0) y durante el periodo analizado, los intervalos de concentración oscilaron entre 0.84 - 24.29 μmol kg-1 y 0.26 - 2.77 μmol kg-1 para el NID y PID, respectivamente. Por debajo de la zona eufótica (150 - 300 m) la variación fue substancialmente menor, tanto para el NID (12.43 - 31.20 μmol kg-1) como para el PID (1.72 - 3.05 μmol kg-1). Particularmente, en mayo y septiembre ambos nutrientes presentaron una marcada reducción de los valores promedio (para el periodo de estudio) en la zona eufótica. Esta reducción representó, respectivamente, el 52 y 53% para NID, y 79 y 49% para el PID (Fig. 3). Por debajo del estrato eufótico (150 - 300 m), las concentraciones promedio menores de NID se observaron en febrero y agosto (81 y 83% del promedio para este estrato, respectivamente), mientras que para el PID fueron similarmente bajos en mayo y agosto (~88%). Los máximos valores promedio se registraron en marzo de 2013 para NID y PID, los cuales correspondieron al 140 y 111% del valor promedio de la zona afótica, respectivamente.
Distribución del trazador N*. El intervalo de valores de N* ajustado para la Cuenca Alfonso oscilaron entre -9.4 y 12.2 μmol kg-1, con un claro predominio de valores negativos entre -8 a 0 que agruparon el ~62% de todo los datos, mientras que un 34% correspondió a los valores positivos. En la distribución vertical, los valores negativos del N* asociados al déficit de nitrógeno (vía desnitrificación) constituyeron el ~40% y ~85% de los datos de la zona eufótica en el estrato entre 150 - 300 m, respectivamente (Fig. 4a-b).
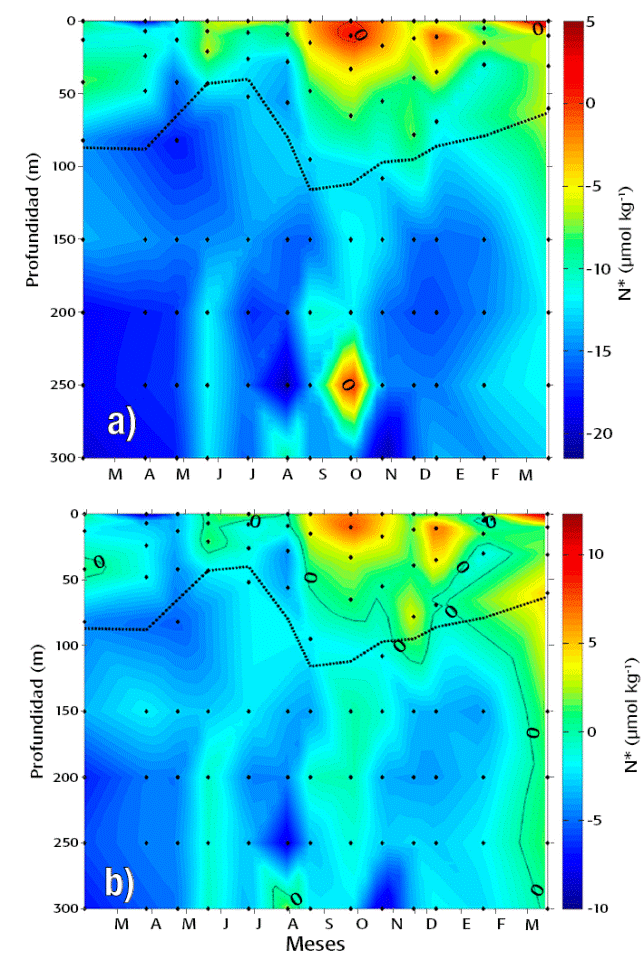
Figuras 4a-b Distribución temporal y vertical del parámetro N* en los primeros 300 m de profundidad de Cuenca Alfonso, Bahía de La Paz, México. (a) Valores estimados a partir de las constantes ajustadas del cociente remineralización (ΔN/ΔP) y del exceso de PID de la covariación ideal con el NID (Kte1) para el sitio. (b) Valores estimados con las constantes originales de la ecuación propuesta por Gruber & Sarmiento (1997). Los valores positivos indican exceso de NID con respecto al PID. La línea discontinua representa la oxipleta de 1.41 ml l-1.
Temporalmente, la incidencia de valores positivos indicadores de exceso de nitrógeno vía fijación biológica de N2 fueron evidentes desde mayo hasta el final del estudio en la zona eufótica (Fig. 4a). Los principales incrementos se presentaron en la zona eufótica entre agosto - octubre, diciembre y marzo de 2013, siendo este último el mes que registró también la mayor expansión vertical (300 m) de valores positivos seguido por noviembre (78 m) y septiembre (65 m). Los menores valores promedio de la zona eufótica (-1.37 a -4.25 μmol kg-1) se detectaron en febrero - abril y julio. Durante estos mismos meses, incluyendo junio, se registraron también las mayores deficiencias de nitrógeno (-3.41 a -5.57 μmol kg-1) en las profundidades entre 150 - 300 m (Fig. 4a). Con excepción de marzo de 2013, los valores positivos en el estrato por debajo de 150 m fueron detectados únicamente en julio a 300 m (2.11 μmol kg-1).
Discusión
La aplicación del trazador N* propuesta por Gruber & Sarmiento (1997) para balances globales y a nivel de cuencas oceánicas, ha ofrecido un punto de partida para el entendimiento de los flujos preponderantes que modulan el inventario del nitrógeno. Estas rutas antagónicas son principalmente la desnitrificación (respiración anaeróbia de la materia orgánica a base de nitrato, con producción de N2 o N2O) y la fijación de N2 (conversión de N2 a nitrógeno biodisponible - NH4+). Según Codispoti (1989), ambos procesos han mantenido el balance global en escalas de ~107 años, aunque el primero eventualmente ha excedido (20 - 30%) la fijación de N2 por periodos de hasta varios miles de años. Así mismo, en escalas espaciales la existencia de diferencias contrastantes entre las señales de estos dos procesos de regulación de las regiones y cuencas oceánicas, ha constituido una característica inherente a la respuesta biológica frente a las condiciones oceanográficas, atmosféricas y, recientemente, a la actividad humana (Gruber, 2004).
Algunos de los enunciados de modelos sobre el presupuesto actual del nitrógeno y la distribución de las principales rutas han sido derivados de la combinación del N* y otros trazadores similares, así como del empleo de indicadores indirectos (e.g., δ15N en material disuelto y particulado, cocientes N2:Ar). Sus resultados han sido posteriormente corroborados con observaciones directas de las poblaciones de microorganismos asociados a los procesos metabólicos (e.g., microscopía, FISH, técnicas moleculares) y mediante determinación, con diversas técnicas, de las tasas fisiológicas en sistemas experimentales y/o in situ (e.g. pareamiento isotópico, asimilación de 15N, reducción de acetileno), lo cual ha brindado una buena correspondencia y complementariedad entre dichas estimaciones (Altabet et al., 1999; Deutsch et al., 2001; Coles et al., 2004; Coles & Hood, 2007; Monteiro, 2009; DeVries et al., 2013). La introducción del sistema de circulación termohalina a estos modelos, por ejemplo, ha permitido catalogar al Atlántico Norte como sumidero de nitrógeno hacia el océano, mientras que a las cuencas del Pacífico e Índico se consideran como regiones de remoción de este elemento hacia la atmósfera (Gruber, 2004).
En México, las evaluaciones preliminares de estos procesos biogeoquímicos a través de aproximación como el N*, han sido pobremente implementados debido a la dificultad para mantener monitoreos continuos y a la escasez de series de tiempo de variables fisicoquímicas. En este sentido, Cuenca Alfonso es un caso interesante, además de por sus características de ser un sistema marginal y altamente influenciado por la costa, para estudiar el ciclo del nitrógeno, porque presenta una condición deficiente de oxígeno en la columna de agua y se cuenta con una serie de tiempo. En este estudio, las modificaciones a la ecuación del N* de algunas de las constantes que definen las propiedades semiconservativas de los reservorios de PID y NID, incluyen el criterio de tomar todas las fracciones disueltas del NID, y no sólo el nitrato o el nitrito + nitrato, en consideración de que en estos ambientes costeros, el amonio eventualmente puede tener una contribución significativa y de que además es potencialmente captado por diversos grupos del fitoplancton para la construcción de biomasa (Nixon, 1981; Dortch, 1990; Howarth et al., 2000; Escobedo-Urías, 2010). En este caso, las concentraciones de amonio aportaron en sus concentraciones altos hasta ~80% del NID, ocurriendo los mínimos porcentajes en profundidad y los mayores en superficie, sugiriendo una activa remineralización. El intervalo de valores de NID y PID observados para el periodo de este estudio es similar a las concentraciones observadas en perfiles verticales de la boca del Golfo de California (Bustos-Serrano & Castro-Valdez, 2006) y el Pacífico Nororiental Tropical (Sutka et al., 2004), no obstante que, por su ubicación dentro de la región sur del Golfo de California y los niveles de clorofila a, se ha considerado como oligotrófica en superficie (Álvarez-Borrego & Lara-Lara, 1991). Por otra parte, el ajuste de los valores de las constantes ΔN/ΔP y Kte1 a la ecuación original con la serie histórica de datos, permite identificar las anomalías del cociente de las concentraciones de nutrientes relativas a la condición local promedio multianual de la cuenca. Esto confiere una mayor definición en la identificación del proceso dominante, cuya influencia sobre la concentración y proporción de nutrientes indica un exceso de nitrógeno nuevo o su remoción respecto a la relación local promedio con el fósforo. No obstante, en el sentido estricto del fundamento del N*, los valores negativos/positivos de N* no aseguran totalmente la exclusión de uno u otro proceso de retroalimentación, solamente las desviaciones al comportamiento conservativo son significativas en la identificación de zonas o periodos en los que la desnitrificación o la fijación de nitrógeno son predominantes (Deutsch et al. 2001).
En el caso particular del ΔN/ΔP, tradicionalmente las estimaciones se han basado en el cociente N:P de ~16, el cual coincide bien con la estequiometría promedio de la asimilación de nutrientes (fotoautotrófica) y remineralización de materia orgánica propuesto por Redfield (1958). Sin embargo, hoy en día varios estudios sobre la variabilidad de la composición elemental del fitoplancton y materia orgánica, exhiben considerables fluctuaciones regionales que dependen de las particularidades ambientales y biológicas de cada ecosistema (Kaiser & Benner, 2012; Martiny et al., 2013; Karl, 2014). En Cuenca Alfonso, la relación entre NID:PID presenta desviaciones significativas al valor propuesto por Redfield (1958) que deben ser consideradas para la implementación de trazadores geoquímicos como el N*. Mientras otras estimaciones con la ecuación original indican el total predominio de las rutas de remoción, a través de los datos obtenidos en este estudio planteamos la incidencia estacional de la fijación de N2 como un proceso conspicuo para el ingreso de nitrógeno al sistema y atenuación del desbalance NID:PID. Esto fortalece nuestra postulación sobre la adecuación de constantes propias para la identificación de las rutas predominantes bajo condiciones muy locales a partir de los datos de registros históricos.
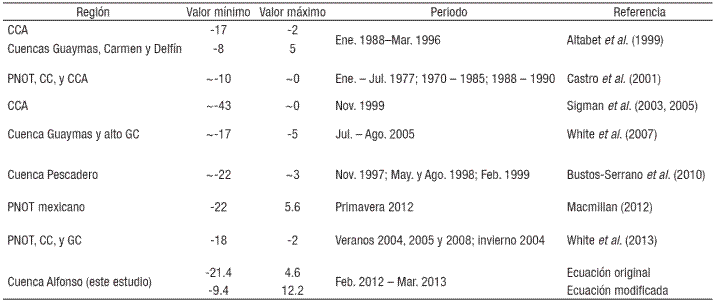
En el presente estudio, los valores de N* revelaron un patrón estacional y vertical muy similar al obtenido con la ecuación original (Spearman = 0.94, p<0.05; Fig. 4b). Sin embargo, la estimación basada en esta ecuación mostró una baja proporción de valores positivos (~3%), y un intervalo de variación mayor (-21.4 a 4.62 μmol kg-1) respecto a los valores obtenidos después de su ajuste (-9.4 a 12.2 μmol kg-1), así como mayores deficiencias de NID comparado con los reportados para el Golfo de California (-19 a -1 μmol kg-1; Altabet et al., 1999; White et al., 2007, 2013) y otras localidades del Pacífico Nororiental Tropical (-22 a 5.6 μmol kg-1; Castro et al., 2001; Macmillan, 2012; Tabla 1).
Tabla 1 Valores comparativos de N* en las regiones del Pacífico Nororiental Tropical y Golfo de California.
La distribución vertical de nuestros datos es coherente con los reportes de N* previamente citados para la región, es decir, mayores valores en superficie que disminuyen abruptamente por debajo de la zona eufótica. Esto, aunado con el comportamiento de la señal isotópica (δ15N) enriquecida en el material orgánico suspendido y el nitrato (Altabet et al., 1999; Sigman et al., 2003, 2005), ha sugerido en la mayoría de los casos el impacto de la desnitrificación local en la columna de agua y/o el efecto por intercambio con masas de agua del Pacífico Nororiental Tropical (Brandes et al., 1998; Sutka et al., 2004). En Cuenca Alfonso, la disponibilidad de materia orgánica y, sobre todo, las condiciones de hipoxia necesarias para la ocurrencia de los procesos de desnitrificación en columna de agua, han sido registradas desde los ~50 m durante la incidencia de un giro ciclónico de submesoescala (Monreal et al., 2001, Hakspiel-Segura, 2014). Asimismo, la hipótesis sobre la influencia de determinadas masas de agua, podría explicar el aumento de los valores del N* en la Cuenca Alfonso durante el verano en coherencia con la llegada de Agua Superficial Tropical (Macmillan, 2012).
Por otro lado, los valores positivos de N* soportan la ocurrencia de la fijación biológica de N2. Por mucho tiempo se asumió que esta actividad, era un proceso prevaleciente en la zona eufótica, llevado a cabo fundamentalmente por cianobacterias diazótrofas estimuladas por la disponibilidad de PID y metales (hierro) en sistemas limitados por NID (e.g., Codispoti, 1989; Mills et al., 2004). Esto coincidió bien con nuestros hallazgos, sin embargo, reconocemos las limitaciones del trazador para esclarecer la posibilidad de que en Cuenca Alfonso, esta actividad pueda ejercer una retroalimentación positiva al balance de nitrógeno por debajo de la zona eufótica, de manera similar a lo que se ha sugerido recientemente para ambientes profundos (>150 m), bajo condiciones de alto contenido de nutrientes e hipoxia (Hamersley et al., 2011; Fernández et al., 2011; Bonnet et al., 2013) y que son análogos al sitio de estudio (Hakspiel-Segura & Martínez-López, 2014). La fijación de N2 es sin duda un proceso que ocurre a lo largo del Golfo de California, incluyendo otras cuencas como Carmen, Guaymas y Delfín (White et al., 2007, 2013). La estimación de sus tasas, así como el registro de la señal isotópica del material orgánico exportado, han indicado que el aporte de nitrógeno nuevo a la zona eufótica, a pesar de ser moderado y muy variable, no se considera como un mecanismo efectivo de retroalimentación positiva a las posibles pérdidas por desnitrificación (White et al., 2013). A pesar de ello, la distribución estacional de valores de N* observada en este trabajo, define con claridad un exceso de nitrógeno en superficie respecto a la proporción N:P promedio del sitio asociado a los periodos con mayor estratificación termohalina de la columna de agua y menores concentraciones de NID.
Es posible que la magnitud de estos flujos negativos de nitrógeno vía remoción por desnitrificación pueda verse incrementado estacionalmente con la disponibilidad de materia orgánica, sustrato para la actividad de microorganismos desnitificadores. Las proliferaciones fitoplanctónicas y su acoplamiento temporal con el zooplancton (De Silva-Dávila & Palomares-García, 2002; Villegas-Aguilera, 2009) favorecen altos flujos de material orgánico particulado desde finales del otoño, se mantienen altos en invierno y repuntan en mayo - junio en respuesta a la intrusión de nutrientes por la influencia de fenómenos oceanográficos, como la mezcla y la elevación de las isopicnas (Aguíñiga et al., 2010; Martínez-López et al., 2012; Silverberg et al., 2014). En este estudio la dinámica temporal reportada para el material orgánico en hundimiento recolectado en trampas de sedimentos (Aguirre-Bahena, datos no publicados; Silverberg et al., 2014), fue consistente con los menores valores del N* observados por debajo de los 150 m, sugiriendo un posible acoplamiento de la remoción de nitrógeno con los niveles de producción biológica en la zona eufótica. Será conveniente en trabajos futuros, corroborar dichos supuestos con la medición de tasas fisiológicas.

