











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
La comunidad de peces comprende un gran número de individuos y biomasa en las lagunas costeras y estuarios (Yáñez-Arancibia, 1985); cuyas larvas y juveniles, especialmente abundantes, se benefician de su alta productividad que los provee de recursos alimentarios y a la vez estos sitios con sus múltiples tipos de hábitats son utilizados como refugio (Cowan et al., 2013).
Las primeras etapas del desarrollo larval de los peces es el periodo más susceptible a los factores bióticos y abióticos (Lasker, 1981), lo que llega a ocasionar variaciones en el reclutamiento de las poblaciones de los adultos (Miller & Kendall, 2009).
Los peces que habitan ambientes estuarinos son tolerantes a cambios drásticos de temperatura, salinidad, concentración de oxígeno y niveles de turbidez, así como a cambios estacionales en éstas y otras condiciones físicas, tales como la apertura y cierre de las bocas de estos sistemas (Able & Fahay, 2010) durante los períodos de lluvias y secas. La mayoría de los peces estuarinos pueden tolerar fluctuaciones ambientales, pero en cada especie depende de su adaptación fisiológica que influye en su distribución dentro de estos ecosistemas (Blaber, 1997).
Las lagunas costeras han cobrado notable importancia no sólo por el papel que juegan en los ciclos biológicos de numerosos peces marinos o dulceacuícolas, sino también por las grandes alteraciones y por el vertimiento diario de múltiples contaminantes (Reséndez-Medina & Kobelkowsky-Díaz, 1991).
La Laguna Madre de Tamaulipas junto con la Laguna Madre de Texas, constituyen el sistema hipersalino más grande del mundo; esta área posee una gran riqueza y diversidad biológica debido a su ubicación, ya que se encuentra en dos regiones biogeográficas: la Neártica y la Neotropical; la Laguna Madre de Tamaulipas recibe agua dulce de dos regiones hidrológicas: la cuenca del Río Bravo y la cuenca de los ríos San Fernando y Soto La Marina, y tiene la influencia de dos provincias marinas: la Carolineana y la Caribeña (CONANP, 2012); la Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) de México la considera la Región Marina Prioritaria Nº 44, como un sitio prioritario para la conservación biológica (Arriaga-Cabrera et al., 1998); en 2004 fue catalogada por la Convención RAMSAR sobre humedales, como Humedal de Importancia Internacional registrado como sitio número 1362; mientras que el Ejecutivo Federal Mexicano la declaró Área Natural Protegida, con el carácter de área de protección de flora y fauna (SEMARNAT, 2005).
Sobre peces óseos adultos de la Laguna Madre, Tamaulipas, se tienen los trabajos de: Hildebrand (1958) quien realizó el primer listado y registra 58 especies, Barba-Macías (1999) señaló 33 especies, Leija-Tristán et al. (2000) encuentran 121 y finalmente Raz-Guzman y Huidobro (2002) identificaron 83 especies; mientras que en la Laguna Madre de Texas se han registrado 94 especies (McKee, 2008), sin embargo es probable que el número de especies presentes en el sistema pudiera ser mayor, toda vez que se trata de un cuerpo de agua con una gran heterogeneidad ambiental espacial y temporal. Por otro lado sobre las fases larvarias, Tolan et al. (1997) analizaron la estructura de la comunidad ictioplanctónica en la parte estadounidense; para la región mexicana de la laguna, existe el trabajo de Ocaña-Luna y Sánchez-Ramírez (1998) sobre la alimentación de larvas de esciénidos; por lo que los objetivos de este trabajo son: determinar la composición ictioplanctónica de la Laguna Madre de Tamaulipas, caracterizar los patrones temporales de abundancia y diversidad en la comunidad, determinar las especies dominantes, establecer la relación de su abundancia larvaria con la salinidad y temperatura del agua superficial y definir las asociaciones de larvas de peces en este sistema.
Materiales y métodos
La Laguna Madre se localiza al norte del Estado de Tamaulipas, entre los paralelos 23° 47' y 25° 57' N y los meridianos 97° 09' y 97° 52' O. La limitan al norte la desembocadura del Río Bravo y al sur la del Río Soto la Marina. El área de estudio se encuentra al sur de la laguna, limitándose al norte con la Bahía de Catán y al sur con la Enramada, en dicha región se localizan la Boca de Catán y La Boca el Caballo, entre los paralelos 24° 14' y 24° 34' N y los meridianos y 97° 41' y 97° 52' W (Fig. 1).
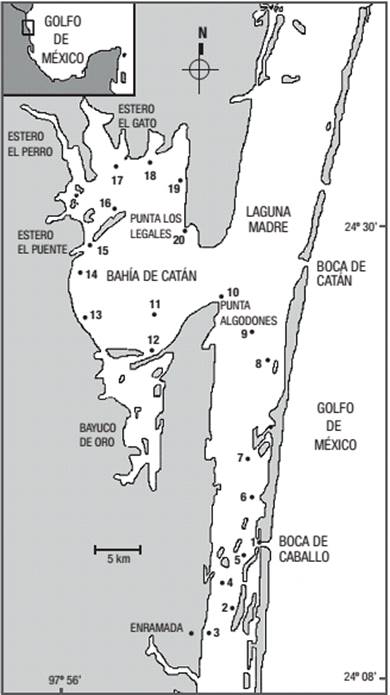
Figura 1: Área de estudio y ubicación de estaciones de muestreo en la Laguna Madre, Tamaulipas (1997-1998).
Esta zona presenta variaciones muy amplias de salinidad de 21-51, por lo que de acuerdo con el Sistema de Venecia (1958), se observan condiciones euhalinas en octubre (35-38), poli-euhalinas en enero (21-36) y eu-hiperhalinas en mayo y julio (33-46 y 36-51, respectivamente); en este último mes la laguna alcanzó la mayor temperatura de todo el ciclo anual de 29.0 a 31.0 ºC, mientras que durante enero ocurrió la menor temperatura del agua (18.0 a 22.0 ºC), provocada por los intensos vientos del norte propios de ésta época (Ocaña-Luna et al., 2008). La laguna presenta salinidades superiores a las del mar la mayor parte del año; amplias variaciones han sido mencionadas con anterioridad por Hildebrand (1958), quien registró en la zona norte del sistema valores de 117 unidades en el mes de julio; así mismo Hildebrand (1969) menciona valores bajos de 9 en noviembre por efecto de fenómenos meteorológicos como los huracanes. Tunnell (2002) señala que la hipersalinidad de la Laguna Madre de Texas tiende a aumentar debido a que el volumen de agua en el sistema ha descendido en los últimos años por el desvió de agua del Río Grande (Río Bravo del Norte) para la irrigación de tierras destinadas a la agricultura, como también lo menciona Edwards y Contreras-Balderas (1991) para la Laguna Madre de Tamaulipas.
Para el presente estudio se estableció una red de 20 estaciones de monitoreo en el área, las cuales cubrieron zonas importantes como: bahía, bocas de comunicación con el mar y desembocaduras de arroyos (Fig. 1). En cada una de las estaciones se tomaron muestras de zooplancton durante los meses de octubre de 1997 (final de la época de lluvias), enero (época de nortes), mayo (época de estiaje) y julio de 1998 (inicio de la época de lluvias), con una red estándar con malla de 505 µm y boca de 50 cm de diámetro, a la cual se le colocó un flujómetro para medir el volumen de agua filtrada; los arrastres se realizaron en la capa superficial de la columna de agua aproximadamente en los primeros 50 cm, de manera circular durante cinco minutos. Las 80 muestras recolectadas fueron fijadas con formalina al 4% neutralizada con borato de sodio. Se tomaron de manera simultánea datos de salinidad y temperatura (°C) del agua superficial con un refractómetro marca "Atago" y un termómetro de mercurio, respectivamente.
De cada muestra se separo el total de larvas de peces y se conservaron en alcohol al 70%; posteriormente se cuantificaron e identificaron al más bajo taxón posible. La abundancia fue expresada en términos de densidad, para lo cual el número de larvas se estandarizó como número de organismos/100 m3.
Las especies identificadas en el sistema fueron ubicadas en las siguientes categorías, considerando la clasificación propuesta por Sanvicente-Añorve et al. (2011) la cual sin embargo fue modificada en algunos casos: a) Residentes Permanentes en la Laguna (RPL) correspondiente a especies que llevan a cabo todo su ciclo de vida dentro de las lagunas; b) especies marinas que ocupan la laguna como área de crianza (MN), aquellas que usualmente viven en el mar pero las larvas y/o juveniles hace un uso extensivo de los ambientes lagunares; c) especies marinas en tránsito (MT) correspondiente a aquellas que usualmente viven en el mar y una pequeña porción de la población usan la laguna y d) Catádromas (C), aquellas especies que viven principalmente en agua dulce y migran al mar a desovar.
Para analizar la similitud de la comunidad ictioplanctónica de la Laguna Madre, Tamaulipas con otros sistemas lagunares cercanos geográficamente como Laguna Madre de Texas (Holt et al., 1990; Tolan et al., 1997; Tolan & Newstead, 2003, 2004 y 2005), Laguna de Tamiahua (Sanvicente-Añorve et al., 2011) y Laguna de Tampamachoco (Ocaña-Luna & Sánchez-Ramírez, 2003), se calculó el coeficiente de similitud de Sörensen (Sörensen, 1948).
Donde:
CCs = coeficiente de similitud
c= número de especies comunes entre ambas comunidades
s1 = número de especies de la comunidad 1
s2 = número de especies de la comunidad 2.
Para analizar las diferencias en la abundancia promedio de larvas de peces entre los meses analizados, estas abundancias fueron transformadas como ln (x+1); posteriormente se realizó un análisis de varianza (ANOVA) y finalmente una prueba de rangos múltiples de Tukey (Sokal & Rohlf, 1995).
Mediante el empleo del coeficiente de correlación de Rangos de Spearman (Zar, 2010) se obtuvo la relación entre la densidad de larvas de las tres especies más abundantes en el sistema, con respecto a la temperatura (ºC) y salinidad.
Se estimaron los siguientes parámetros ecológicos: riqueza (S) como número de especies. La diversidad de especies dentro de la comunidad fue calculada con el índice de Shannon-Wiener (Shannon & Weaver, 1963).
Donde:
H´ = índice de diversidad de especies de Shannon-Wiener
pi = proporción de la especie i
s= número de especies en la muestra.
La dominancia de las especies en la comunidad se obtuvo con el índice de Simpson (Simpson, 1949).
Donde:
D = índice de dominancia
pi = proporción de la especie i en la comunidad
s = número de especies en la muestra.
Para determinar las especies dominantes de peces en etapa larvaria se aplicó el Índice de Valor Biológico propuesto por Sanders (1960).
Donde:
IVB= Índice de valor biológico, i= cada una de las especie
j= cada una de las épocas de recolecta. En este índice el valor de importancia para cada especie se asigna en función de su abundancia en cada época, expresada como puntajes, por lo que las especies se ordenan con base en la constancia espacio-temporal de sus abundancias (Loya-Salinas & Escofet, 1990).
Se determinó la formación de grupos de especies para lo cual se aplicó el índice de similitud de Bray-Curtis (Bray & Curtis, 1957), previo a la aplicación de este índice los datos fueron transformados de la siguiente manera: ln (x+1).
Donde:
BC = índice de similitud de Bray-Curtis
Xij y Xik = diversidad de la especie i en las muestras j y k
s= número de especies Hammer
Finalmente a partir de la matriz de similitud se elaboró un dendrograma utilizando el método de grupos pareados no ponderados (UPGMA), esto se llevó a cabo utilizando de programa PAST versión 2.01 (Hammer et al., 2001).
Resultados
Se recolectaron un total de 8,480 larvas de peces, de las cuales se identificaron 39 especies pertenecientes a 21 familias, Opisthonema oglinum (Lesueur, 1818), Microgobius thalassinus (Jordan & Gilbert, 1883), Gobiesox strumosus Cope, 1870, Citharichthys arctifrons Goode, 1880, Lupinoblennius nicholsi (Tavolga, 1954), Ctenogobius shufeldti (Jordan & Eigenmann, 1887) y Acanthostracion quadricornis (Linnaeus, 1758), se consideran nuevos registros para la Laguna Madre, Tamaulipas; ya que anteriormente no habían sido recolectadas ni como larva ni como adulto, además las últimas cuatro especies, tampoco tienen registros previos en la Laguna Madre de Texas.
Las familias mejor representadas fueron Gobiidae y Sciaenidae con ocho y seis especies, respectivamente. En relación a la riqueza específica, en mayo se presentó el mayor número de especies (24), mientras que en octubre el menor (17) (Tabla 1).
Tabla 1: Abundancia promedio mensual (no. de organismos/100 m3), abundancia relativa (%) de larvas de peces, IVB= Índice de Valor Biológico y riqueza específica. Categorías ecológicas. Laguna Madre, Tamaulipas, México. 1997-1998. Taxas ordenados por IVB
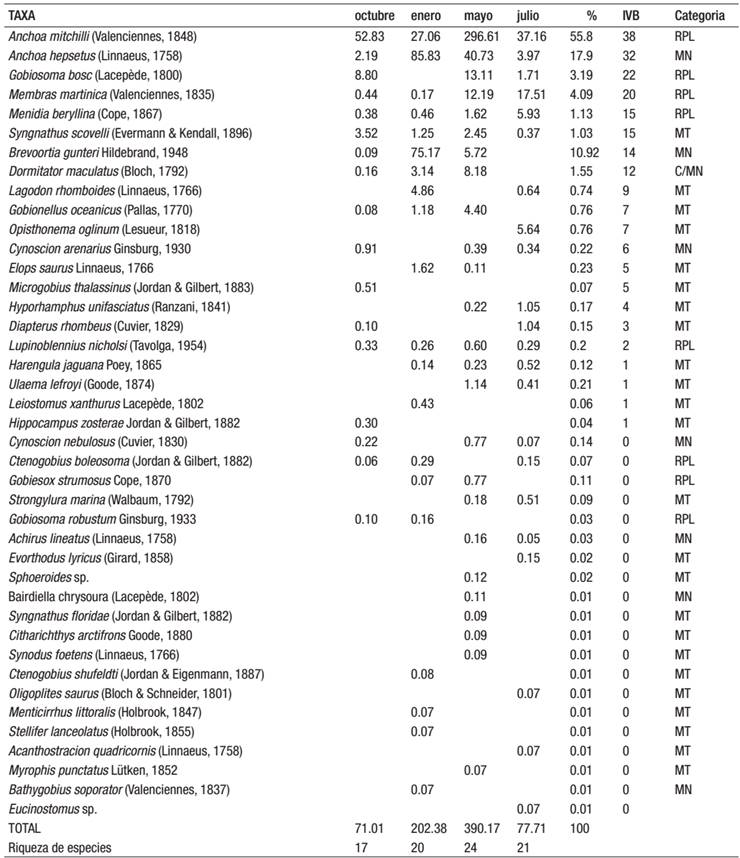
Residentes Permanentes en la Laguna (RPL)= especies que llevan a cabo todo su ciclo de vida dentro de las lagunas; especies marinas que ocupan la laguna como área de crianza (MN)= especies que usualmente viven en el mar pero las larvas y/o juveniles hace un uso extensivo de los ambientes lagunares; marina en tránsito (MT)= especies que usualmente viven en el mar y una pequeña porción de la población usan la laguna; Catádroma (C)= especies que viven principalmente en agua dulce y migran al mar a desovar.
El 80% de las especies que conforman la comunidad ictioplanctónica pertenecen a las categorías de marina en tránsito (MT) y especies marinas que ocupan la laguna como área de crianza (MN) (Tabla 1).
Este sistema presenta una mayor similitud en cuanto a la composición específica con la Laguna Madre, Texas, compartiendo 27 especies y con la Laguna de Tampamachoco, Veracruz 23 (similitud= 0.66 y 0.68, respectivamente), mientras que con la Laguna de Tamiahua, Veracruz comparte 21 especies (similitud= 0.59) (Tabla 2).
Tabla 2: Especies de larvas de peces presentes en lagunas costeras de Texas, Estados Unidos; Tamaulipas y Veracruz, México. Taxa ordenado alfabeticamente.

TEX= Laguna Madre de Texas (1Holt et al., 1990; 2Tolan et al., 1997; 3Tolan & Newstead, 2003; 4Tolan & Newstead, 2004; 5Tolan & Newstead, 2005)
TAMA= Laguna Madre de Tamaulipas (este trabajo)
TAMI= Laguna de Tamiahua, Veracruz (Sanvicente-Añorve et al., 2011)
TAMPA= Laguna de Tampamachoco (Ocaña-Luna & Sánchez-Ramírez & 2003)
La mayor abundancia de larvas se presentó en mayo, mientras que en octubre la menor, y el análisis de varianza detectó diferencias en las abundancias de las larvas entre estos dos meses (F3, 76= 4.995, p < 0.005). Las larvas del género Anchoa, constituyen más del 70% de la abundancia total en la Laguna Madre de Tamaulipas. Aanchoa hepsetus (Linnaeus, 1758) y A. mitchilli (Valenciennes, 1848) estuvieron presentes durante todo el año; esta última especie es considerada como residente permanente en la laguna (RPL) y presentó la mayor abundancia en mayo (periodo cálido) cuando ocurre su principal época de desove; representando el 55.8% de la abundancia promedio anual; mientras que A. hepsetus el 17.9%, y es clasificada como una especie marina, cuyas larvas y/o juveniles penetran a los sistemas lagunares para protegerse y alimentarse (MN), tuvo las mayores abundancias en enero (época fría) justo cuando ocurrió un pico en su desove (Tabla 1).
Otras especies también abundantes en la laguna son: Brevoortia gunteri Hildebrand, 1948 (10.9%), especie marina que ocupan la laguna como área de crianza (MN); Membras martinica (Valenciennes, 1835) (4.1%) y Gobiosoma bosc (Lacepède, 1800) (3.2%), ambas residentes permanentes en la laguna (RPL) y Dormitator maculatus (Bloch, 1792) (1.55%), cuyos adultos viven principalmente en agua dulce, migran al mar a desovar y sus larvas y/o juveniles hacen un uso extensivo de los ambientes lagunares (C/MN) (Tabla 1).
Se presentó una correlación positiva entre la abundancia de larvas de A. mitchilli con la temperatura y salinidad (rs= 0.376 y rs=0.467, respectivamente), esta especie es considerada euritérmica y eurihalina; mientras que A. hepsetus presentó una correlación negativa con la temperatura (rs= - 0.247), debido probablemente a que su principal período reproductivo ocurre en el invierno; finalmente en B. gunteri se observó una correlación negativa tanto con la temperatura (rs= -0.567) como con la salinidad (rs= -0.436), ya que al igual que A. hepsetus presentan un pico de desove en el período frío (Tabla 3).
Tabla 3: Correlación de la abundancia de larvas de peces (no. de organismos/100 m3) con respecto a la temperatura (°C) y salinidad del agua superficial de la Laguna Madre, Tamaulipas.

* correlación significativa; rs= correlación de rangos de Sperman; α= nivel de significancia
El índice de diversidad de Shannon-Wiener presentó valores bajos tanto en octubre (1.40 bits/ind) como en mayo (1.02 bits/ind) Figura 2. cuando A. mitchilli presentó alta abundancia; la mayor diversidad se observó en julio (2.23 bits/ind) cuando la riqueza de especies fue alta.

Figura 2: Parámetros ecológicos de la comunidad ictioplanctónica de la Laguna Madre, Tamaulipas. México. 1997-1998.
La dominancia presentó valores altos en octubre y mayo (0.57 y 0.59, respectivamente), y tuvo un comportamiento inverso a la diversidad, debido a las altas densidades que presentó A. mitchilli (Fig. 2).
De acuerdo con el IVB, fueron diez las especies dominantes que caracterizaron a la comunidad y en conjunto representan el 97.82% de la abundancia total (Tabla 1).
La asociación de especies de larvas de peces definidas por el índice de Bray-Curtis, arrojó ocho grupos (Fig. 3): Grupo I (dos especies, 4.9%).- formado por especies con mayor abundancia en invierno, de las cuales Lagodon rhomboides (Linnaeus, 1766) pertenecen al grupo dominante (IVB= 9); Grupo II (tres especies, 7.3%).- especies exclusivas o con mayor abundancia en invierno; Grupo III (7 especies, 17.1%).- especies exclusivas o con bajas densidades en el mes de mayo; Grupo IV (diez especies, 24.4%).- especies típico-lagunares o que dependen de este sistema en alguna etapa de su vida, el cual está conformado por las especies dominantes de acuerdo con el Índice de Valor Biológico: A. mitchilli (RPL), A. hepsetus (MN), G. bosc (RPL), M. martinica (RPL), Syngnathus scovelli (Evermann & Kendall, 1896) (MT), Menidia beryllina (Cope, 1867) (RPL); B. gunteri (MN), D. maculatus (C/MN), Gobionellus oceanicus (Pallas, 1770) (MT) y O. oglinum (MT), y Grupo V (nueve especies, 22.0%).- con bajas densidades, la mayoría de ellas ausentes en enero; Grupo VI (cuatro especies, 9.8%).- formado por especies de las familias Gobiidae y Sciaenidae, exclusivas de invierno.- Grupo VII (dos especies, 4.9%). formado por exclusivas de octubre con bajas densidades y Grupo VIII (cuatro especies, 9.8%) especies exclusivas del mes más cálido (julio).

Discusión
El número de especies que conforman la comunidad ictioplanctónica en este sistema es ligeramente menor (39), que las señaladas por Holt et al. (1990), Tolan et al. (1997), Tolan y Newstead (2003, 2004, 2005) para la Laguna Madre de Texas (43), de ellas 16 especies no se recolectaron en la Laguna Madre en Tamaulipas, mientras que 12 especies de este sistema no se han registrado para la Laguna Madre en Texas; esta diferencia probablemente se deba a que el esfuerzo de muestreo no ha sido el mismo, pero también a que en la recolecta realizada en el sistema de la Laguna Madre de Texas, ocurrieron una gran cantidad de organismos juveniles cuyos adultos no necesariamente desovan en la laguna. Por lo anterior se tiene que la comunidad ictioplanctónica de la Laguna Madre (Texas-Tamaulipas) está conformada por 55 especies.
Al comparar la riqueza específica con otras lagunas costeras geográficamente cercanas y hacia el sur, como la Laguna de Tamiahua y la Laguna de Tampamachoco en el Estado de Veracruz (Sanvicente-Añorve et al., 2011; Ocaña-Luna & Sánchez-Ramírez, 2003) registraron 32 y 29 especies, respectivamente, seguramente esta riqueza mayor es un reflejo de una mayor área y heterogeneidad de hábitats en Laguna Madre.
Por otro lado la Laguna Madre de Tamaulipas comparada con la Laguna Madre de Texas y las Lagunas de Tamiahua y Tampamachoco en Veracruz, presenta una similitud de más del 50% con los sistemas lagunares de Tamiahua y Tampamachoco, lo cual se debe a su ubicación biogeográfica en: sur de la provincia Carolineana, norte de la provincia Caribeña y zona de transición entre ellas, donde confluye la fauna ictiológica de ambas regiones. Dicha fauna se ve influenciada por cambios climáticos (subtropical y tropical) y tambien por la circulación oceánica de la zona descrita por Zavala-Hidalgo et al. (2003), quienes mencionan que existe una circulación estacional: que va de Louisiana y Texas a Tamaulipas y Veracruz de septiembre a marzo, la cual se revierte de mayo a agosto. En esta región se observa un remplazo gradual de especies que tienen su límite de distribución hasta las costas de Texas como Adinia xenica (Jordan & Gilbert, 1882) y otras que se encuentran en latitudes al norte o al sur en el Golfo de México como: Dorosoma cepedianum (Lesueur, 1818) hasta el Río Pánuco, Tamaulipas; Fundulus grandis Baird & Girard, 1853 y Sciaenops ocellatus (Linnaeus, 1766), hasta el Estado de Veracruz (McEachran & Fechhelm, 1998 y 2005) o como Menidia peninsulae (Goode & Bean, 1879) (Castro-Aguirre, 1978) y B. gunteri hasta la Laguna de Términos, Campeche, México (Reséndez-Medina, 1979).
En relación a la abundancia del ictioplancton en la Laguna Madre, Tamaulipas, el género Anchoa constituye más del 70% de la densidad total y también en la Laguna Madre de Texas, como lo señalan Tolan et al. (1997). Los engráulidos en general son considerados dominantes a lo largo de la costa Atlántica de los Estados Unidos y Golfo de México, ya que alcanzan el 50% del total de la captura fuera de las costas de Alabama (Hernandez et al., 2010) y hasta el 85% en Carolina del Norte (Hettler & Chester, 1990). En Alabama se han registrado cuatro especies del género Anchoa, entre las que se encuentran A. mitchilli y A. hepsetus, esta última considerada como una de las más abundantes en la región (Hernandez et al., 2010), ambas especies se presentaron de forma abundante en la Laguna Madre durante el presente estudio.
Anchoa mitchilli, es considerada una especie dominante en sistemas estuarino-lagunares, particularmente en la porción norte el Golfo de México en su fase adulta por Reis y Dean (1981), Vougliotis et al. (1987) y Griffth y Bechler (1995) , mientras que en el sur Castillo-Rivera et al. (1994) la registraron en la Laguna de Pueblo Viejo, Veracruz, en su fase de huevo Sánchez-Ramírez y Ocaña-Luna (2002) en el mismo sistema donde alcanza el 97.7% del total de huevos recolectados, por otro lado Ocaña-Luna y Sánchez-Ramírez (2003) mencionan también que las larvas de esta especie se presentan en gran abundancia en la Laguna de Tampamachoco, Veracruz.
La distribución de A. mitchilli se extiende desde la costa atlántica de los Estados Unidos (Hoese & Moore, 1998) hasta el norte de la Península de Yucatán en México (Sanvicente-Añorve et al., 2011). Es una especie que desova durante todo el año, con un pico máximo en mayo (período cálido), lo cual también se ha observado en diferentes localidades de su distribución, como la Bahía de Chesapeake (Onley, 1983; Wang & Houde, 1995), Laguna de Pueblo Viejo, Veracruz (Sánchez-Ramírez & Ocaña-Luna, 2002), Laguna de Tamiahua, Veracruz (Gaspar-Dillanes & Sánchez-Iturbe, 1985, Flores-Coto et al., 1983) y Laguna de Términos, Campeche (Flores-Coto et al., 1988).
Las larvas de A. mitchilli alcanzan la mayor abundancia de mayo a agosto en los sistemas estuarino lagunares de la porción mexicana del Golfo de México: Pueblo Viejo, Tamiahua y Tampamachoco (Sánchez-Ramírez & Ocaña-Luna, 2002; Flores-Coto et al., 1983; Ocaña-Luna & Sánchez-Ramírez, 2003).
Otra de las especies abundantes fue A. hepsetus la cual desova en la zona costera, por ejemplo en Carolina del Norte a menos de 16 Km de la costa (Mansueti & Hardy, 1967), mientras que en sur del Golfo de México frente a las costas de Campeche, las mayores abundancias de su etapa larvaria se encuentran en el talud y borde de la plataforma continental (Hernández, 1990). Esta especie desova todo el año, con mayor intensidad en los meses de abril a junio (Santos-Valencia et al., 1998). La mayor densidad de larvas de la especie ocurre de febrero a marzo en la Laguna de Tampamachoco (Ocaña-Luna & Sánchez-Ramírez, 2003) lo cual coincide con la época de mayor abundancia de larvas en la Laguna Madre y el principal periodo de desove en la Laguna de Términos (Flores-Coto et al., 1988).
En las lagunas costeras hay especies que desarrollan todo su ciclo de vida dentro del sistema (ejemplo: A. mitchilli), mientras que otras desovan en la zona nerítica adyacente a las lagunas y penetran en ellas en algún estadio larvario o como juvenil (ejemplo: A. hepsetus y B. gunteri). Al respecto Raz-Guzman y Huidobro (2002) señalan que en la Laguna Madre, el 93% de las especies de peces adultos son marinas, de las cuales el 69.7% son eurihalinas y 23.3% son estenohalinas, mientras que el 7.0% lo componen especies que permanentemente habitan las lagunas costeras.
Brevoortia gunteri es la tercera especie más abundante en la Laguna Madre, con la mayor densidad en enero, lo cual ocurre de manera similar en la Laguna de Tamiahua (Flores-Coto et al., 1983) y Laguna de Términos (Flores-Coto, 1987), donde se encuentran con mayor abundancia en marzo y de enero a marzo, respectivamente.
Se presenta una correlación positiva entre la abundancia de larvas de A. mitchilli con la temperatura y salinidad; esta especie es considerada euritérmica y eurihalina. Houde y Zastrow (1991) mencionan que las larvas de A. mitchilli ocurren en temperaturas entre 15 y 30 °C y en el sistema estuarino se encontraron larvas hasta en 31 °C. Otra especie, A. hepsetus, presentó una correlación negativa con la temperatura, ya que tiene un desove principalmente invernal; finalmente B. gunteri presentó una correlación negativa tanto con la temperatura como con la salinidad, ya que al igual que A. hepsetus, presenta un pico de desove en el período de menor temperatura; además de que A. mitchilli y A. hepsetus toleran amplios intervalos de salinidad, desde agua dulce hasta aguas hipersalinas (Nizinski & Munroe, 2002).
Membras martinica se reproduce en Laguna Madre todo el año, al igual que en la Laguna de Tamiahua (Flores-Coto et al., 1983) y Laguna de Tampamachoco (Ocaña-Luna & Sánchez-Ramírez, 2003), con sus mayores densidades de junio a noviembre. En la Sonda del Misisipi las larvas de esta especie han sido consideradas junto con A. mitchilli en un grupo denominado nectónico costero, que ocurre en salinidades moderadamente altas (Rakocinski et al., 1996).
Gobiosoma bosc es una especie que presenta dependencia por las conchas de Crassostrea virginica (Gmelin, 1791) como hábitat para su anidación (Harding & Mann, 2000); en el norte del Atlántico Shenker et al. (1993) señala una elevada densidad de larvas en el estuario del Río Patuxent, Maryland, y en los estuarios de Carolina del Sur desovan de mayo a septiembre y utilizan el interior de las ostras vacías como un sustrato para el desove (Crabtree & Middaugh, 1982), mientras que en las costas de Misisipi Hendon et al. (2000), registraron las larvas de mayo a octubre, con la mayor abundancia en mayo-junio. En el sur del Golfo de México se presenta una reproducción continua a lo largo del año, con la mayor abundancia en el periodo de primavera-verano en las lagunas de Tamiahua (Flores-Coto et al., 1983), Tampamachoco (Ocaña-Luna & Sánchez-Ramírez, 2003) y de Términos (Flores-Coto, 1987).
Los adultos de D. maculatus bajan de la cuenca de los ríos para desovar cerca de las lagunas salobres y estuarios (Miller et al., 2005). Se han registrado larvas y juveniles en lagunas costeras en intervalos de tallas (Lp, mm) que van de 8.2-9.5 en las lagunas Madre, Tamiahua y Tampamachoco (Ocaña-Luna, 2000) y de 8.2-13.3 en la Laguna de Alvarado, Veracruz (Flores-Coto & Zavala-García, 1982). Los ejemplares se recolectaron en gran abundancia hacia las zonas de mayor influencia marina, como las bocas de las lagunas y estuarios, lo cual puede indicar que los desoves se efectúan en la zona marina adyacente y luego las larvas penetran a los sistemas estuarino-lagunares cuando sus tallas son mayores a los 8.0 mm.
La diversidad de la comunidad de larvas de peces en la Laguna Madre de Tamaulipas fue menor (1.02-2.23 bits/ind) que en la Laguna de Tampamachoco (1.8-3.1 bits/ind) como lo señalan Ocaña-Luna y Sánchez-Ramírez (2003), ésto posiblemente se deba a que se presenta una menor dominancia de especies en este sistema. El aumento de la diversidad de las larvas hacia el sur del Golfo de México, también se ve influenciado por el decremento en la abundancia de la especie dominante A. mitchilli, a lo largo del gradiente latitudinal (Sánchez-Ramírez & Ocaña-Luna, 2002).
La conformación de especies del grupo IV, el cual determina la estructura de la comunidad de larvas de peces de la Laguna Madre de Tamaulipas, coincide con el grupo base de la comunidad ictioplanctónica en la Laguna de Tamiahua por la presencia de B. gunteri, A. mitchilli, M. martinica y Gobiosoma bosc (Flores-Coto, 1987); y con el grupo de especies residentes y/o más abundantes (A. hepsetus, G. bosc y M. martinica) en la Laguna de Tampamachoco (Ocaña-Luna & Sánchez-Ramírez, 2003).
La Laguna Madre, Tamaulipas presenta una gran heterogeneidad ambiental por su fisiografía la cual está conformada por: ríos, esteros, arroyos, bocas de comunicación con el mar, islas, bahías, zonas con vegetación sumergida y manglar, lo cual da origen a una mayor riqueza y diversidad de peces. Una especie clave en la comunidad íctica de este sistema es A. mitchilli, cuya alta frecuencia, abundancia y biomasa, se observa en la mayor parte de su distribución, desde la Bahía de Chesapeake costa este de EUA (Wang & Houde, 1995) hasta la Laguna de Términos en Campeche (Flores-Coto et al., 1988), debido a que es eurihalina, que puede tolerar salinidades de 0.5-80 (Whitehead, 1978), de ahí que en una laguna hipersalina como la Laguna Madre, donde particularmente en la porción sur se registran salinidades hasta de 51, esta especie sea la dominante.
En la Laguna Madre de Tamaulipas, por a su condición hipersalina el 80% de las especies que conforman la comunidad ictioplanctónica pertenecen a las categorías de especies marinas en tránsito (MT) y especies que utilizan la laguna para crianza (MN), a diferencia de lo que ocurre en el sistema lagunar Indian-River, Florida, con condiciones polihalinas y marinas donde es del 66%, este tipo de ambientes permiten una mayor riqueza de especies marinas, las cuales podrían ser excluidas de sistemas que presentan condiciones oligohalinas (Gilmore, 1995). Por otro lado Edwards y Contreras-Balderas (1991) mencionan que la disminución del volumen de agua del Río Grande (Río Bravo del Norte), debido a su uso para irrigación y el incremento de la contaminación, han provocado la disminución de especies dulceacuícolas en la parte baja de la cuenca y han sido reemplazadas por especies estuarinas y marinas.

