










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.25 no.1 Ciudad de México ene./abr. 2015
Artículos
Carbono orgánico nuevo frente a la Península de Baja California: series de tiempo y climatología
New organic carbon in front of the Baja California Peninsula: time series and climatology
Benigno Hernández de la Torre1, Gilberto Gaxiola Castro2, Saúl Álvarez Borrego3, Artemio Gallegos García4 y Raúl Aguirre Gómez5
1 Instituto Nacional de Ecología y Cambio Climático (INECC). Periférico Sur 5000, Colonia Insurgentes-Cuicuilco, Delegación Coyoacán, C.P. 04530, México, D.F.
2 Departamento de Oceanografía Biológica. División de Oceanología. Centro de Investigación Científica y de Educación Superior de Ensenada, CICESE, Carretera Ensenada-Tijuana, No. 3918, Zona Playitas, C.P. 22860, Ensenada, Baja California, México.
3 Departamento de Ecología. División de Oceanología. Centro de Investigación Científica y de Educación Superior de Ensenada, CICESE, Carretera Ensenada-Tijuana, No. 3918, Zona Playitas, C.P. 22860, Ensenada, Baja California, México.
4 Instituto de Ciencias del Mar y Limnología, UNAM, Circuito Exterior, Ciudad Universitaria, C.P. 04510, México, D.F.
5 Instituto de Geografía, UNAM, Circuito Exterior, CU, C.P. 04510, México, D.F. bhernan99@gmail.com
Recibido: 30 mayo 2014.
Aceptado: 17 julio 2014.
Resumen
Se presenta la serie de tiempo de la producción fitoplanctónica nueva (Pnueva) frente a la Península de Baja California de 1970 a 2008, calculada por modelos estadísticos de la relación temperatura-nitrato en diez grupos de estaciones costeras y oceánicas de las Líneas 90, 107, 120, 137 y 157 de la red CalCOFI-IMECOCAL. La Pnueva se calculó a partir de la razón-f y la producción primaria total (Ptotal). Espacialmente las zonas más productivas fueron las oceánicas de las Líneas 90, 120 y 157. La Pnueva en las Líneas 107 y 137 presentó valores bajos y se comportó como un parte-aguas entre las regiones norte y sur. Estacionalmente la Pnueva fue alta en la transición primavera-verano; disminuyó en el verano y el otoño, con aumento en invierno. La Pnueva se relacionó con el índice de surgencias costeras y se concluye que para un índice de surgencias costeras menor a 200 m3/s/100 m, la Pnueva aumenta y el máximo se produce en el mismo período que las surgencias costeras. Para un índice superior a 200 m3/s/100 m la Pnueva disminuye y ambas series se desfasan temporalmente.
Palabras claves: Océano Pacífico, Península de Baja California, producción nueva, surgencias costeras.
Abstract
Time series of new phytoplankton production (Pnew) off Baja California from 1970 to 2008 computed using statistical models of temperature-nitrate relationship in ten groups of coastal and oceanic stations (Lines 90, 107, 120, 137 and 157) of the CalCOFI-IMECOCAL network, are presented. Pnew was calculated from the f-ratio and total primary production (Ptotal). Spatially, the most productive areas of the region were Lines 90, 120 and 157. Pnew of lines 107 and 137 had relatively low values. These two lines marked the border between the northern and southern areas of the studied region. Seasonally, Pnew was high in the spring-summer transition, decreased in summer and autumn, and increased in winter. Pnew was related to coastal upwelling index, and we conclude that for an index of less than 200 m3/s/100 m, Pnew increases and its maximum occurs in phase with the coastal upwelling index. For an upwelling index greater than 200 m3/s/100 m Pnew decreases and both series are out of phase.
Keywords: Baja California Peninsula, Coastal upwelling, new primary production, Pacific Ocean.
INTRODUCCIÓN
El transporte vertical de nitratos hacia la zona eufótica (ZE) es el factor principal en la regulación de la biomasa y producción del fitoplancton en las aguas costeras de la Corriente de California (CC), donde la distribución vertical y la profundidad del inicio de la nutriclina están correlacionadas con la razón de producción (Eppley et al., 1979). Parte de esta fracción de producción fotosintética juega un rol importante en la captación del CO2 de la atmósfera hacia el océano, por lo que su estudio en diversas zonas como la CC es de reconocida importancia debido al aumento continuado del CO2 atmosférico en los últimos años a escala mundial (Gao et al., 2012; Li et al., 2012; Reay et al., 2008).
La producción del carbono orgánico nuevo (o Pnueva) es la fracción de la producción primaria que se genera a partir de una fuente de nitratos alóctonos (Dugdale y Goering, 1967) presentes por debajo de la termoclina estacional y transportada hacia las capas superficiales del océano. La Pnueva es considerada como la porción de la producción fitoplanctónica que es exportada, por debajo de la ZE, como materia orgánica particulada sin que el ecosistema pelágico se colapse (Eppley, 1989).
Estudios de Pnueva en el Océano Pacífico nororiental han sido realizados por Eppley y Peterson (1979), Eppley (1992) y por Kudela y Chavez (2000), acerca de la producción fitoplanctónica y su relación con la concentración de los nitratos. Otros autores, como Hayward y Venrick (1998), combinaron la relación entre factores físicos (profundidad de la termoclina) y la entrada de nitratos hacia la ZE en la CC. En aguas mexicanas se han llevado a cabo estudios de Pnueva en el Golfo de California y Golfo de México (Hidalgo & Álvarez-Borrego, 2004; Hidalgo et al., 2005).
Las observaciones de los océanos mantenidas a largo plazo (series de tiempo) son necesarias para mejorar la comprensión y predicción de los cambios en los ecosistemas, con la finalidad de lograr un manejo adecuado de los recursos marinos y costeros. La utilización de los índices climáticos como el "El Niño" o "La Niña", en relación a los cambios del ecosistema pelágico del océano, es una herramienta que se puede utilizar para explicar y predecir efectos de los cambios ambientales en una región específica. En este trabajo se presenta la variabilidad interanual del carbono orgánico nuevo desde enero de 1970 hasta diciembre del 2008 en áreas costeras y oceánicas frente a la Península de Baja California, con el objetivo de determinar los procesos o perturbaciones hidroclimáticas que han incidido en su variación interanual y las causas probables de los cambios sobre escalas diversas.
MATERIALES Y MÉTODOS
El área de estudio comprende la región nororiental del Océano Pacífico frente a la Península de Baja California (Fig.1). Se utilizó la red de estaciones del programa CalCOFI-IMECOCAL (www.calcofi.org; http://imecocal.cicese.mx/) y en particular las estaciones hidrográficas de las Líneas 90, 107, 120, 137 y 157. Las posiciones de las estaciones analizadas se agruparon en costeras y oceánicas, de acuerdo al criterio de Lynn y Simpson (1987) (Fig. 1).
Temperatura Superficial del Mar (TSM). La TSM media mensual de septiembre de 1969 a diciembre de 2008, estimada en 1° x 1° de latitud y longitud, proviene del sitio Web: http://www.pfeg.noaa.gov/products/PFEL/observed/GTS_surface/sfc_temp.html. La climatología (30 años de datos enero de 1960 a diciembre de 1990) de la TSM se obtuvo por R. Mendelssohn, PFEG-NOAA (comunicación personal). Para la confección de los modelos de la relación temperatura-nitratos (NO3, μM) por mejor ajuste, se utilizaron los datos de CalCOFI-IMECOCAL para las líneas anteriormente señaladas. Estos datos se estandarizaron para las profundidades de 0, 10, 20, 30, 50, 100, 150 y 200 m.
Razón-f. Esta razón es un estimado cuantitativo entre la Pnueva y la producción primaria total (Ptotal) (f=Pnueva/Ptotal) y varía entre 0 y 1 (Dugdale & Goering, 1967). Con base en los datos reportados por Eppley et al. (1979) para el SCB (33°N; 119°W) y el modelo empírico propuesto por Harrison et al. (1987) para la misma zona, se calculó la razón-f con la ecuación: razón-f = f max[1-e-(mNO3/fmax)], donde fmax es el valor máximo esperado y m es la pendiente inicial de la curva (NO3 versus f). Partiendo de que los valores altos de NO3 cercanos a la superficie se presentan con mayor frecuencia en las estaciones costeras, se tomó fmax = 0.67 y m = 13.7 (Tabla II del modelo empírico de Harrison et al., 1987); mientras que en las estaciones oceánicas se presenta una disminución progresiva de NO3 a medida que se alejan de la costa, se tomó fmax = 0.61 y m = 10.5 (Tabla II del modelo empírico de Harrison et al., 1987). Para obtener la razón-f intermedia en la ZE se calculó un promedio ponderado por luz a cada profundidad por medio del coeficiente de atenuación de la luz difusa (Kd) y la ley de Lambert-Beer. La información sobre el promedio de Kd [Kd = 4.6/(Z1%)] se obtuvo a partir de los modelos propuestos por Millán-Nuñez et al. (1996) y de los datos derivados de los cruceros IMECOCAL, donde se estimaron los valores promedios de la profundidad de la ZE (Z1%) a partir de la posición del máximo profundo de clorofila en la columna de agua y valores de Kd in-situ, respectivamente. Se calcularon valores promedios de Kd para la época fría (invierno-primavera) y para la época cálida (verano-otoño) para cada estación hidrográfica. A partir de las consideraciones anteriores se calculó el promedio ponderado de la razón-f en la ZE.
Producción primaria total (Ptotal). El valor de la Ptotal se calculó con el modelo empírico propuesto por Smith y Eppley (1982) para la SCB: Ptotal = e (-3.78 - 0.372 (ΔT) + 0.227 (D)), donde ΔT es la anomalía de la temperatura superficial (°C) definida como el valor del dato menos la media climatológica mensual de los años analizados; (D) es la duración del día expresado en horas-luz para cada mes en función de la latitud (http://aa.usno.navy.mil/AA/data). Pnueva (gC m-2 d-1) se obtuvo del producto de la razón-f y la Ptotal según Dugdale y Goering (1967).
Surgencias Costeras. Para relacionar la Pnueva con las surgencias costeras (m3/s/100 m de línea de costa) se obtuvieron los valores medios mensuales del índice de surgencias costeras (Fig. 1) según el sitio Web:
http://www.pfeg.noaa.gov/products/PFEL/modeled/indices/upwelling/upwelling.html
en cuatro sitios frente a la Península de Baja California: Línea 90 costera (33° N y 119° O); Línea 107 costera (30° N y 119° O); Línea 120 costera (27° N y 116° O) y Línea 137 costera (24° N y 113° O).
Los fenómenos y eventos de "El Niño" y "La Niña" se obtuvieron según el sitio Web: http://www.cpc.ncep.noaa.gov/products/precip/CWlink/MJO/enso.shtml
Las series de tiempo fueron suavizadas mediante un filtro paso-baja de promedios móviles (12 meses) para sustraer la estacionalidad.
RESULTADOS
Relación Temperatura-Nitratos. La relación entre la temperatura y nitratos de las cinco líneas de estudio (Fig. 2) se utilizaron con el fin de estimar la concentración de nutrientes en cada zona. Existe una correspondencia directa entre las dos variables en la columna de agua, tanto para las estaciones costeras como las oceánicas. Sin embargo, la relación temperatura-nitratos no fue la misma por región, ya que el intercepto se fue ampliando a medida que la latitud disminuyó representadas por las líneas costeras y las oceánicas, lo que implica que a temperatura de 22 °C aún hay nitratos pero en menor concentración. El intercepto del eje de la temperatura aumentó hacia el sur como se muestra en la Línea 90, donde corta en 15.0 y 14.5 °C en las estaciones costera y oceánica, respectivamente; en la Línea 107 a 16.0 y 15.5 °C; en la Línea 120 a 17.7 y 18.0 °C; en la Línea 137 a 18.4 y 18.2 °C y en la Línea 157 a 21.6 y 22.5 °C. Los coeficientes de determinación de las relaciones lineales explican entre el 80 y 90% de la varianza total.

Anomalías de Temperatura. El área de estudio se caracterizó por anomalías negativas en los primeros años de la década de los setentas. Un período de anomalías positivas de aproximadamente 22 años (entre 1977 y 1998) se presentó en las estaciones costeras y oceánicas de las líneas más norteñas. Se muestra igualmente, un intervalo de anomalías negativas a partir de 1999, pero menos pronunciado que en los años setenta (Fig. 3a). Los máximos de anomalías de temperatura en las estaciones oceánicas fueron menores que las costeras. Las anomalías de mayor amplitud se concentraron en la zona de la Línea 120 costeras, disminuyendo hacia el norte y hacia el sur. La Línea 107 oceánica presentó los valores más altos en el último periodo de anomalías negativas de temperatura. La Línea 157 oceánica presentó valores negativos durante más tiempo que el resto de las líneas. En general, la señal de anomalías negativas entre 1985 y 1989 fue más intensa al sur y disminuyó hacia el norte. De acuerdo con esta tendencia, el régimen frío en la región frente a Baja California terminó aproximadamente entre 1976 y 1977, originando un cambio hacia anomalías positivas de la TSM. Los valores extremos de anomalías positivas se presentaron en los años 1982-84, 1992-93 y 1997-98, con mayor impacto en las áreas costeras debido al predominio de eventos "El Niño". Durante este período largo de anomalías positivas se registraron algunos años con anomalías negativas, como lo fueron 1985, 1989 y 1995, relacionadas principalmente con la presencia de eventos hidrometeorológicos fuertes en la región, originados por cambios bruscos en el gradiente de presiones atmosféricas por la presencia de eventos "La Niña". En la climatología de TSM, los valores máximos ocurren entre agosto y octubre, mientras que los mínimos entre enero y abril. La Línea 90 oceánica mostró el valor mínimo de 14.48 °C y el máximo de 18.09 °C. La estación oceánica de la Línea 157 mostró la temperatura mínima más alta de la región (21.62 °C), mientras que la posición costera tuvo la temperatura máxima de 27.09 °C (Fig. 3b). En general, se presentó una diferencia latitudinal de 13.41 °C entre Punta Dana, California y Cabo San Lucas, Baja California Sur (Fig. 1 y 3b).
Anomalías de Ptotal. La serie de las anomalías de Ptotal de la Línea 90 presentó un período de valores positivos altos en los primeros años de la década de los setenta, disminuyendo a partir de 1976-77 y con un cambio hacia anomalías negativas en 1982-83 (Fig. 4a). En algunas zonas la tendencia negativa llegó hasta 1986, como es el caso de la Línea 120 oceánica. Entre 1985 y 1989 la Ptotal se recuperó hasta alcanzar anomalías positivas, aunque no tan altas como en la década de los 70s, pero sí suficientes para obtener el tercer máximo importante en la estación costera de la Línea 157. En la década de los 90s las anomalías de Ptotal fueron negativas, producto de dos eventos cálidos diferentes: uno largo en el periodo 1992-1994, otro muy fuerte (1997-1998) y en medio un evento frío anómalo en 1995, que permitió la recuperación de la productividad a niveles medios, principalmente en las estaciones costeras. A partir de 1999, los períodos de anomalías positivas de producción primaria total estuvieron relacionados con eventos fríos (1999-2000 y 2007-2008) con tendencia a la recuperación. En la climatología de Ptotal, los máximos costeros de Ptotal se obtuvieron de abril a agosto, mientras que en las estaciones oceánicas fueron de mayo a agosto en las estaciones de la Línea 120 y en las costeras de las Líneas 137 y 157 llegaron a septiembre (Fig. 4b). Las estaciones oceánicas de las Líneas 137 y 157 tuvieron comportamientos diferentes en los valores máximos, con la primera de mayo a agosto, mientras que la más sureña sólo tuvo tres meses (mayo-julio) con valores máximos. Los valores mínimos se observaron de noviembre a febrero. El máximo valor medio de la Ptotal fue en junio en la Línea 157 oceánica (0.74 gC m-2 d-1) y el mínimo se registró en julio en la Línea 120 costera (0.46 gC m-2 d-1).
Razón-f. Las estaciones de la Línea 90 presentaron valores altos de f en toda la serie, seguida de la 120 y 157 oceánica. Los valores altos de f se presentaron en los primeros años de los 70s; sin embargo, es marcado el descenso hasta 1984 (Fig. 5a). La razón-f se recuperó en casi todas las estaciones a partir de 1994. La climatología de la razón-f (1970-2008) en general presenta valores altos en la época fría (meses DEFMAM) y bajos en la época cálida (JJASON). Los valores medios de razón-f se incrementaron con la latitud durante la época fría. De la Línea 120 oceánica hacia el norte y hacia el sur los valores decayeron en la época fría. El norte, centro y sur del área de estudio presentaron valores altos de razón-f para el área oceánica (Fig. 5b).
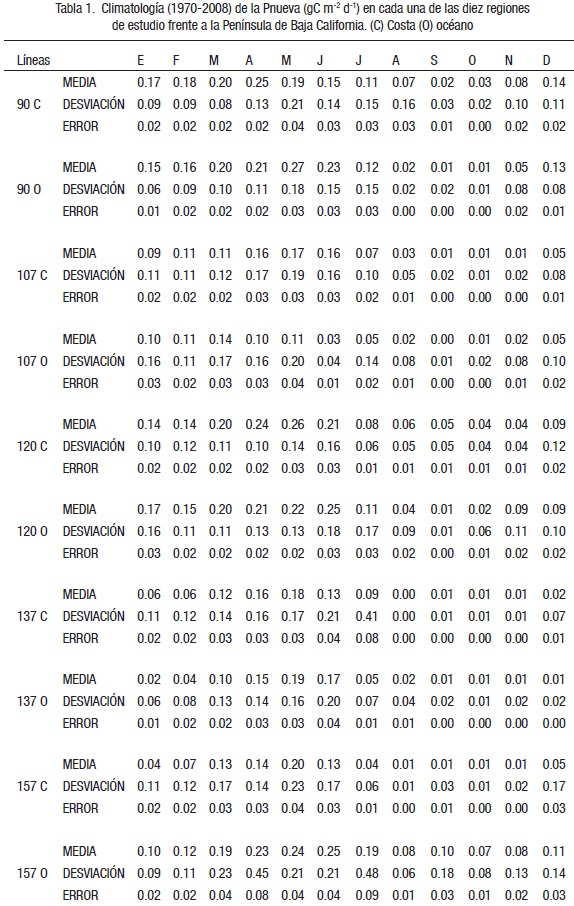
Anomalías de Pnueva. La serie de Pnueva presentó valores de anomalías positivas en los primeros años de la década de los setenta, con una ligera inflexión hacia valores negativos durante el evento cálido de 1972-1973 (Fig. 6a). En 1975 se observaron anomalías positivas en las estaciones oceánicas y en las costeras. Las anomalías presentaron valores negativos a partir del evento cálido de 1976-1977, las que continuaron debido a eventos similares durante 1982-1984 y 1986-1988. En 1979-1980 las anomalías presentaron signos positivos en las estaciones costeras, no así en las oceánicas. La serie muestra una disminución de las anomalías positivas hacia el norte en 1985 y 1989 en las costeras, mientras que las oceánicas no presentaron valores positivos (Fig. 6a). Sólo la Línea 157 oceánica mostró un ligero incremento en las anomalías. Una señal positiva se manifestó desde 1990 hasta el primer semestre de 1992, con la disminución durante el evento cálido (1992-1993 y 1994). La señal positiva de 1995 se reflejó con intensidad sólo en las localidades costeras de las Líneas 90 y 107. El evento cálido de 1997-1998 afectó la región de estudio, inclusive con los valores más bajos de anomalías negativas. A mediados de 1998 comenzó el incremento en las anomalías de Pnueva, continuó durante el evento frío de 1999-2000 principalmente en las oceánicas de sur a norte y disminuyó en las costeras de las Líneas 120 a la 90. No se presentaron anomalías positivas en la Línea 157 costera desde 1989. Un evento cálido de bajas proporciones (El Niño-débil) afectó toda la zona, con una disminución ligera de los valores de Pnueva durante 2002-2003. Después de este último evento, la zona recuperó la producción nueva con anomalías positivas a causa de la presencia de eventos fríos en la región sur de la CC que han afectado la mayoría de las estaciones, excepto la más sureña de la Línea 157. Para la climatología de la Pnueva (1970-2008) la primavera es la época de mayor producción nueva, aunque en algunas temporadas se adelantaron en marzo y otros se retrasaron a junio, como las oceánicas de las Líneas 107 y 120, respectivamente (Fig. 6b). En mayo se presentaron los valores medios mayores de la Línea 120 y abril en las costeras de las Líneas 90 y 107. Diciembre presentó valores altos de Pnueva en las Líneas 90, 120 y 157, mientras que los de las líneas intermedias como la 107 y 137 fueron bajos. El valor medio más alto se observó en la estación oceánica de la Línea 90, mientras que el más bajo en la 107 oceánica (Fig. 6b). Destacan los valores bajos en otoño. Las áreas consideradas con muy baja producción nueva en la época otoñal son las estaciones costeras de las Líneas 107, 137 y 157, así como la 137 oceánica (Fig. 6b). La Tabla 1 muestra los estadígrafos de la climatología (valores medios, desviación estándar y error) de la Pnueva.
Relación Pnueva-Índice de Surgencias Costeras (Pnueva-ISC). En abril se presentó el máximo de Pnueva (0.25 ± 0.02, gC m-2 d-1) para la Línea 90 costera (Fig. 7b), asociada con un valor de ISC de aproximadamente 200 m3/s/100m (Fig. 7a) para el mismo mes. Sin embargo, mayo y junio presentaron máximos de ISC en el intervalo entre 275 y 300 m3/s/100 m, lo que corresponde a una Pnueva baja. La Línea 107 costera presenta niveles de ISC cercanos a 200 m3/s/100 m en abril, mayo y junio (Fig. 7c), coincidiendo con los tres meses de máxima Pnueva (0.160.17 ± 0.03, gC m-2 d-1) para la misma estación costera (Fig. 7d). Las estaciones costeras de las Líneas 120 y 137 mostraron una variación similar en los primeros seis meses, tanto del ISC (Fig. 7e, g) como de Pnueva (Fig. 7 f, h): el primero presentó valores por debajo de 200 m3/s/100m, mientras que el segundo 0.25 (±0.03) gC m-2 d-1 y 0.18 (±0.03) gC m-2 d-1, respectivamente. El análisis de correlación cruzada entre Pnueva y el ISC en las cuatro líneas de estudio mostró que las estaciones costeras de las Líneas 120 y 137 indican máxima productividad sin desfase en meses con respecto al ISC, mientras que las Líneas 90 y 107 presentan desfase de tres meses con respecto del máximo de Pnueva (Tabla 2).
DISCUSIÓN
Para que ocurra Pnueva se requiere de luz suficiente para el crecimiento del fitoplancton, aguas ricas en nutrientes y relativamente bajas temperaturas en superficie, que por lo general ocurre en zonas de surgencias costeras (Eppley et al., 1979, Bakun & Parrish, 1980, 1991). El promedio de duración de un evento de intensificación de surgencias costeras es de 7-10 días (Zimmerman et al., 1987) y el tiempo que le toma al fitoplancton incorporar los nutrientes (nitratos) y efectuar la fotosíntesis varía, ya que las aguas recién afloradas tienen baja disponibilidad de biomasa, lo cual reduce la capacidad para la asimilación instantánea de nutrientes y su posterior crecimiento (Eppley et al., 1979). De acuerdo con Lara-Lara et al. (1980) y Millan-Nuñez et al. (1982) el fitoplancton toma unos seis días para utilizar los nitratos después de un evento de intensificación de surgencia costera. Por esta razón, el máximo de biomasa del fitoplancton está temporalmente desacoplado de las concentraciones máximas de nitratos en el agua recién aflorada (Haury & Shulenberger, 1998). Por ello se requiere de un estudio continuo sobre este aspecto, donde al menos la escala de muestreo sea diaria. Sin embargo, tratándose de ecosistemas marinos y conociendo los procesos que en ellos se llevan a cabo, se pueden interpretar algunos signos que pueden dar una respuesta de la relación Pnueva-ISC a una escala mensual.
Los resultados de este estudio demuestran que con un ISC promedio de 200 m3/s/100m de línea de costa, o menor, la Pnueva aumenta y en la escala de promedios mensuales no tiene desfase con ISC, e inversamente para un ISC mayor de los 200 m3/s/100m de línea de costa.
A partir de este resultado se puede construir una ventana ambiental óptima (Lasker, 1978; Cury & Roy, 1989) que indique que un ISC por arriba de 200 m3/s/100 mno contribuye al máximo simultáneo de Pnueva, ya que un aumento en la velocidad del viento produce mayor turbulencia, que hace que la capa de mezcla se haga más profunda y el fitoplancton no se fotoaclimate a una irradiancia particular en la ZE, lo que coadyuva a la disminución de la razón-f. Estudios de afloramientos costeros y sus efectos sobre la producción biológica, indican que en los sistemas de corrientes de las costas oeste de los continentes, la producción óptima en las pesquerías ocurre dentro de un límite de velocidad del viento que no excede los 5-7m s-1 (Schwing & Mendelssohn, 1997). Aun así, en este estudio se muestra una tendencia al incremento de la razón-f a partir del año 2000 en la región de estudio.
La tendencia espacio-temporal de la Pnueva observada en el área de estudio está relacionada con los cambios a gran escala del esfuerzo del viento. A finales de invierno e inicio de primavera, cuando los vientos se incrementan, el índice de afloramiento se eleva y sube la termoclina a niveles superficiales (Parrish et al., 1981; Husby & Nelson, 1982). Esto trae consigo un acarreamiento hacia la superficie de nitratos subsuperficiales, aumentando así los valores de razón-f y Pnueva (Eppley et al., 1979).
Los resultados de la serie de tiempo y climatología de la TSM expuestos anteriormente, parecen indicar un enfriamiento del océano frente a la Península de Baja California hasta el 2008, en concordancia con los resultados obtenidos por Belkin (2009). En este trabajo los NO3 aparecen relacionado con la temperatura en una relación causa-efecto y se muestran en secuencia latitudinal desde la superficie hasta los 200m.
Los resultados de la variabilidad interanual de 1970 a 2008 muestran que los eventos "El Niño" afectaron la Pnueva disminuyéndola, principalmente desde 1976-77 hasta 1997-98, siendo las estaciones costeras las más vulnerables a esta señal. En la serie analizada las anomalías (positivas-negativas) representadas por máximos en la TSM, Ptotal, razón-f y Pnueva que se transmiten de sur a norte o viceversa, pueden estar relacionados con ondas barotrópicas o intensos gradientes de presión entre la baja presión de las Aleutianas y las altas presiones sobre el continente, con vientos paralelos a la costa que originan surgencias costeras por el efecto del transporte de Ekman, frente a la Península de Baja California. Parés-Sierra y O'Brien (1989) demostraron que gran parte de la energía contenida en las bajas frecuencias de registros de nivel del mar en las costas de California y Baja California, es introducida al océano en latitudes bajas, pero transmitida hacia el polo en forma de ondas costeras de Kelvin (Ripa, 1997). Este modo contribuye de manera significativa a la energía total de la zona relacionada con fenómenos de escala interanual (Parés-Sierra, 1991).
Con los resultados de la variabilidad interanual de la Pnueva en la CC (1970-2008), se puede afirmar que existe una tendencia a la recuperación de la Pnueva frente a Baja California. A partir de lo anterior, se puede dar respuesta a interrogantes sobre fluctuaciones pesqueras tanto costeras como oceánicas, considerando que la Pnueva del fitoplancton es el principal sustento de la red trófica marina (Hernández et al., 2003). Los valores de Pnueva representan la capacidad de carga del carbono orgánico a consumir en el océano y por tanto la cantidad tope que se puede extraer sin descompensar el ecosistema epipelágico. Los resultados mostrados exponen una aproximación cuantitativa de la fertilidad en áreas costeras y oceánicas frente a la Península de Baja California y brindan información necesaria en función del cambio climático marino.
REFERENCIAS
Bakun, A. & R. H. Parrish. 1980. Environmental inputs to fishery population models for Eastern Boundary Currents Regions. IOC, Workshop Report 28: 67 104. [ Links ]
Bakun, A. & R. H. Parrish. 1991. Comparative studies of coastal fish reproductive habitats: the anchovy (Engraulis anchoita) of the southwestern Atlantic. ICES. Journal of Marine Science 48: 343 361. [ Links ]
Belkin, I. M. 2009. Rapid warming of LME. Progress in Oceanography 81: 207-213. [ Links ]
Cury, P. & C. Roy. 1989. Optimal environmental window and pelagic fish recruitment success in upwelling areas. Canadian Journal of Fisheries and Aquatic Science 46: 670-680. [ Links ]
Dugdale, R. C. & J. J. Goering. 1967. Uptake of new and regenerate forms of nitrogen in primary productivity. Limnology Oceanography 12: 196-206. [ Links ]
Eppley, R. w., Renger, E. H. & w. G. Harrison. 1979. Nitrate and phytoplankton production in Southern California coastal waters. Limnology Oceanography 24: 483-494. [ Links ]
Eppley, R. W. & B. J. Peterson. 1979. Particulate organic matter flux and planktonic new production in the deep ocean. Nature 282: 677-680. [ Links ]
Eppley, R. W. 1989. New Production: History, Methods, Problems. In: Berger, W. H., V. S. Smetacek y G. Wefer (Eds.). Productivity of the Ocean: Present and Past. Wiley, New York: pp. 85-97 [ Links ]
Eppley, R. W. 1992. Chlorophyll, photosynthesis and new production in the Southern California Bight. Progress in Oceanography 30: 117-150. [ Links ]
Gao, K., E. W. Helbling, D. P. Háder & D. A. Hutchins. 2012. Responses of marine primary producers to interactions between ocean acidification, solar radiation, and warming. Marine Ecology Progress Series 470: 167-189. [ Links ]
Harrison, W. G., T. Platt & M. R. Lewis. 1987. "f"-ratio and its relationship to ambient nitrate concentration in coastal waters. Journal of Plankton Research 9: 235-248. [ Links ]
Haury, L. & E. Shulenberger. 1998. Surface nutrient enrichment in the California Current off Southern California: description a possible causes. Deep Sea Research II 45: 1577-1601. [ Links ]
Hayward, T. L. & E. Venrick. 1998. Nearsurface pattern in the California Current: coupling between physical and biological structure. Deep Sea Research II 45: 1617-1638. [ Links ]
Hernández-de la Torre, B., G. Gaxiola-Castro., S. Álvarez-Borrego., J. Gómez-Valdés & S. Nájera-Martínez. 2003. Interannual variability of new production in the southern region of the California Current. Deep-Sea Research II 50: 2423-2430. [ Links ]
Hidalgo-González, R. M. & S. Álvarez-Borrego. 2004. Total and new production in the Gulf of California estimated from ocean color data from the satellite sensor SeaWIFS. Deep-Sea Research II51: 739-752. [ Links ]
Hidalgo-González, R. M., S. Álvarez-Borrego, C. Fuentes-Yaco & T. Platt. 2005. Satellite-derived total and new phytoplankton production in the Gulf of Mexico. Indian Journal of Marine Sciences 34: 408-417. [ Links ]
Husby, D. M. & C. S. Nelson. 1982. Turbulence and vertical stability in the California Current. CalCOFI Report 23: 113-129. [ Links ]
Kudela, R. M. & F. P. Chávez. 2000. Modeling the impact of the 1992 El Niño on new production in Monterey Bay, California. Deep Sea Research II 47: 1055-1076. [ Links ]
Lara-Lara, J. R., S. Álvarez-Borrego & L. F. Small. 1980. Variability and tidal exchange of ecological properties in a coastal lagoon. Journal of Estuarine and Coastal Marine Science 11: 613-637. [ Links ]
Lasker, R. 1978. The relation between oceanographic conditions and larval anchovy food in the California Current: identification of factors contributing to recruitment failure. Rapports et Proces-verbaux des Reunions. Conseil International pour l´Éxploration de la Mer 173: 212-230. [ Links ]
Li, Q. P., P. J. S. Franks, M. D. ohman & M. R. Landry. 2012. Enhanced nitrate fluxes and biological processes at a frontal zone in the Southern California Current System. Journal of Plankton Research 34: 790-801. [ Links ]
Lynn, R. J. & J. J. Simpson. 1987. The California Current System: The seasonal variability of its physical characteristics, Journal of Geophysical Research 92: 12,947-12,966. [ Links ]
Millán-Nuñez, R., S. Álvarez-Borrego & D. M. Nelson. 1982. Effects of physical phenomena on the distribution of nutrients and phytoplankton productivity in a coastal lagoon. Estuarine, Coastal and Shelf Science 15: 317-335. [ Links ]
Millán-Nuñez, R., S. Álvarez Borrego & C. C. Trees. 1996. Relationship between deep chlorophyll maximum and surface chlorophyll concentration in the California current system. CalCOFI Reports 37: 241-250. [ Links ]
Parés-Sierra, A. & J. J. O'Brien. 1989. The seasonal and interannual variability of the California Current system: A numerical model. Journal of Geophysical Research 94: 3159-3180. [ Links ]
Parés-Sierra, A. 1991. Remote and local forcing of the Rossby wave variability in the midlatitude Pacific Ocean. Geofísica Internacional 30: 121-134. [ Links ]
Parrish, R. H., C. S. Nelson & A. Bakun. 1981. Transport mechanisms and reproductive success of fishes in the California Current. Biological Oceanography 1: 175-203. [ Links ]
Reay, D. S., F. Dentener, P. Smith, J. Grace & R. A. Feely. 2008. Global nitrogen deposition and carbon sinks. Nature Geoscience 1: 430-437. [ Links ]
Ripa, P. 1997. Ondas y dinámica oceánica. In: Lavín, M. F. (Ed.). Contribuciones a la oceanografía Física en México, Monografía 3, Capítulo 3, Unión Geofísica Mexicana, Ensenada, Baja California, México, pp. 47-74. [ Links ]
Schwing, F. B. & R. Mendelssohn. 1997. Increased coastal upwelling in the California Current. Journal of Geophysical Research 102: 3421-3438. [ Links ]
Smith, P. E. & R. W. Eppley. 1982. Primary production and the anchovy population in the Southern California Bight: comparison of time series. Limnology Oceanography 27:1-17. [ Links ]
Zimmerman, R. C., J. N. Kremer & R. C. Dugdale. 1987. Acceleration of nutrient uptake by phytoplankton in a coastal upwelling ecosystem: a modeling analysis. Limnology Oceanography 32: 359-367. [ Links ]

