










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.25 no.1 Ciudad de México ene./abr. 2015
Artículos
Phenological variation of Rhizophora mangle and ground water chemistry associated to changes of the precipitation
Variación fenológica de Rhizophora mangle y química del agua intersticial asociada a cambios de la precipitación
Claudia Maricusa Agraz Hernández1, Carlos Armando Chan Keb1, Silvia Iriarte-Vivar 2, Gregorio Posada Venegas 1, Beatriz Vega Serratos1 y Juan Osti Sáenz1
1 Instituto EPOMEX, Universidad Autónoma de Campeche, Av. Agustín Melgar s/n entre Juan de la Barrera y Calle 20, Col. Buenavista, A.P. 24039 San Francisco de Campeche, Campeche, México.
2 Departamento de Ecología y Recursos Naturales, Facultad de Ciencias, Universidad Nacional Autónoma de México, Circuito Exterior, Ciudad Universitaria, México D.F. 04510, México. e-mail: clmagraz@uacam.mx
Recibido: 29 julio 2013.
Aceptado: 29 abril 2014.
Abstract
Mangrove forests are vulnerable to long term changes of temperature and precipitation. In this study, we want to know if the precipitation regime significantly varies among years and seasons during a five-yr period (2006-2010) at the North of Laguna de Términos, Campeche, Mexico. In addition, we want to know how the salinity and the redox potential of the ground water of a monospecific mangrove forest of Rhizophora mangle are modified according to this variation. Finally, we describe the total litterfall, and the flowers and hypocotiles production of the studied mangrove and analyze these responses as a function of the environment variables. A repeated measures ANOVA showed that precipitation significantly varied among years and seasons (F=15.83, p<0.003; F=29.8, p<0.0001). The salinity and the redox potential were also significant effects of the same temporal variation (F=1003.25, p<0.0001; F=213.32, p<0.0001), as well as the total litterfall and the hypocotile production (F=39.35, p<0.001; F=8.74, p<0.025). Precipitation was directly proportional to the hypocotile production (r=0.43, p<0.005) and inversely proportional to the salinity ground water (r= - 0.36, p<0.02). The precipitation and the salinity play significant roles on the variation of the litterfall and the hypocotile production of the studied mangrove (R2= 0.85; p<0.05 and R2= 0.70; p<0.05). Our study suggests a resilient ability of R. mangle, which allows it to tolerate seasonal and annual changes of the precipitation.
Keywords: Litterfall, mangroves, redox potential, salinity, standardized precipitation index.
Resumen
Las comunidades de mangle son vulnerables a los cambios de precipitación y temperatura por periodos largos. El presente estudio tuvo como objetivo el conocer si el régimen de la precipitación varió significativamente entre años y temporadas durante cinco años (2006-2010) en la región norte de la laguna de Términos Campeche, México, y además saber si las concentraciones de salinidad y los valores del potencial redox del agua intersticial de un bosque monoespecífico de Rhizophora mangle, variaron significativamente entre años y temporadas, paralelamente a los cambios de la precipitación. Así mismo, se describe la variación anual y estacional de la producción total de hojarasca, y de flores e hipocótilos, del bosque de R. mangle durante el mismo período de estudio, se analizaron todas las respuestas en función de las variables ambientales. El análisis de varianza de medidas repetidas efectuado, validó la variación de la precipitación (F=15.83, p<0.003; F=29.8, p<0.0001), salinidad, potencial redox del agua intersticial (F=1003.25, p<0.0001; F=213.32, p<0.0001) y de la producción de hojarasca e hipocótilos (F=39.35, p<0.001; F=8.74, p<0.025) del bosque de manglar entre años y temporadas del 2006 al 2010. Se encontró que la precipitación fue directamente proporcional a la producción de hipocótilos (r=0.43, p<0.005) e inversa con la salinidad del agua intersticial (r= - 0.36, p<0.02). La precipitación y la salinidad son factores determinantes de la producción de hojarasca e hipocótilos (R2= 0.85; p<0.05 y R2= 0.70; p<0.05). Los resultados de este estudio sugieren una habilidad resiliente de R. mangle, la cual le permite tolerar la variación anual y estacional de la precipitación.
Palabras claves: Hojarasca, índice de precipitación estandarizada, manglares, potencial redox, salinidad.
INTRODUCTION
Climate variation, on a seasonal and a yearly basis, affect the temporal and spatial patterns of precipitation and temperature at local and global scales in many coastal regions of the world (Cook et al., 1992). This variation is mainly caused by El Niño/The South Oscillation (ENSO), and by the Pacific Decade Oscillation (DPO), climatic phenomena that cause disruption of the normal ranges of intensity, frequency, and duration of storms and hurricanes, droughts, hails, and frosts. Overall consequences of these phenomena are effects on biodiversity, on the ecosystem services, and also, on the general wellness of the human society (Rodríguez-Herrera & Bozada-Robles, 2010). Mangrove communities are particularly vulnerable to changes in the precipitation regimes, because the hydroperiod, and the physicochemical conditions of the sediment and the ground water, are directly modified by the climatic variables (Drexler & Ewel, 2001; Flores-Verdugo et al., 2007). A minimum change in the dynamics of the precipitation could cause a response of the entire biota in terms of composition and species richness of mangroves, as well as in their productivity (Mitsh & Gosselink, 1993). According to Field (1995), changes in the precipitation may cause differences in the distribution and the surface of mangrove ecosystems, affecting particularly diverse regeneration processes of these ecosystems. Salinity gradients of the ground water, derived from changes of the precipitation (as well as from the inundation regime and the microtopography among other hydroperiod factors), may act as regulating determinants of physiologic processes in mangrove communities, such as tree photosynthesis and growth (Lichtenthaler, 1996).
Mexican mangroves also depend in a great extent on geomorphology and tectonics (Carranza-Edwards et al., 1975), and regional climatic variables (Flores-Verdugo efal., 1992). The Gulf of Mexico, for example, has a wide continental platformand a high precipitation regime. These conditions have favored the development of large areas of coastal wetlands (Toledo-Ocampo, 2005) along the Atlantic coast. Particularly, Tabasco and Campeche, Mexican states located at the South of the Gulf of Mexico, include the ecological coastal unit most important of Mesoamerica, having the largest cover of mangrove forests in Mexico (CONABIO, 2009). Particularly, Campeche has the 25% of the total area of mangroves in Mexico. It includes Laguna de Términos, the largest system of wetlands in Campeche (107, 262 hectare in 2010: Agraz-Hernández et al., 2012).
Here, we studied the patterns of annual and seasonal variation of the precipitation at the North of Laguna de Términos, Campeche, Mexico, during a five-yr period. We also describe the variation of the ground water chemistry (salinity and redox potential) of a monospecific mangrove forest of a fringe type (with a trend to be a riverine forest) of Rhizophora mangle Linneo during the same period, as well as the productivity, based on litterfall, and the reproductive phenology (flowers and hypocotile production) of the mangrove. We expect that the variability of the ground water chemistry, and the biological responses of this mangrove forest will reflect the variation of the precipitation regime during 2006-2010. We ask the following questions: (1) How does the precipitation vary, on a yearly and seasonal basis, during the period of study?, (2) Are there significant differences among years and seasons in the variation of the salinity and the redox potential of the ground water?, (3) What are the responses to this variation in terms of the productivity (total litterfall) and the reproductive phenology (flower and hypocotile production) of the studied mangrove?, and (4) Which environmental factors may be possible determinants of the variation of these responses?
MATERIAL AND METHODS
Study area. Laguna de Términos locates at the South of the Gulf of Mexico and at the southwestern part of the Yucatan Peninsula, between the meridians 91° 10 and 92° 52'W, and the parallels 18° 23' y 18° 52'N, in a transitional zone between calcareous terrain from the Yucatan Peninsula and the alluvial terrains of the Gulf of Mexico (Álvarez-Arellano & Gaitán-Morán, 1994). Isla del Carmen is a barrier island that has an area of 112.5 km2. It locates at the North of the Laguna de Términos (Thom, 1969). There are two permanent communication channels to the sea, the inlets- Puerto Real and Del Carmen, at the extreme points of the lagoon. The surrounding zone of the lagoon is part of the Area Natural Protegida Laguna de Términos (natural area under a conservation regime of federal adscription), which is one of the most productive coastal areas in Mexico. At the southern and western parts of Campeche and the borderline with Tabasco state, the climate is described as hot and humid with abundant precipitation in the summer, Am(f); the rest of the area has a hot and subhumid climate, Aw(w) (García, 1981). The minima and maxima evaporation rates are 1,055.9 and 2,274.5mm yr1, respectively, and the ranges of precipitation and temperature are 366.6-1915 mm yr-1and 20-49.5°C, respectively. Sediments of Isla del Carmen show a dominance of transitional sands (80.2-67.4%), with a content of 14.4 to 49.1% of calcium carbonate and a content of 16 to 67% of organic matter. The borders of the lagoon are covered by mangrove forests, especially dense at the southwest, eastern and south-southeastern parts of Isla del Carmen. Rhizophora mangle and Avicennia germinans (Linneo) Linneo are the dominant mangrove species (Coronado-Molina et al., 2012). The monospecific forest of R. mangle of this study is located at the midwest of the leeward face of Isla del Carmen, in Campeche, Mexico (Fig. 1).
Drought Analysis. We characterized the distribution of the rainfall for the period of study, on a yearly and seasonal basis, through the Standardized Precipitation Index (McKee et al., 1993). This parameter allows to evaluate the deficit or excess of precipitation of a site for a specific period of time (from 1 month to 4 years), having as a reference a long term data set of the region (Bordi & Sutera, 2007; Rossi et al., 2007). Most SPI values commonly fall in the range -2 to +2, which correspond to distinct drought categories. A SPI that is near to 0 means that the conditions are regular; if the SPI is below 0, it indicates that the accumulated precipitation for the study period is lower than the mean of the reference period; when the SPI is -0.8, then a moderate drought period initiates; values less than -2.0 indicate that there is an exceptional drought period (Nagarajan, 2009) (Table 1).
In order to calculate the SPI index different data sets were used for our period of study. The first data set corresponds to the annual precipitation of the nearest meteorological station to the study site (Isla Aguada, No. 4015) for the long term period 1959-2010.This data set was used to detect a pattern of drought periods from 1959 to 2010, and to obtain a mean annual precipitation value of reference. The second set of data corresponds to the same station for our period of study (2006-2010); it includes the annual and the seasonal precipitation of all years and seasons of that period. We used these data to calculate the mean annual precipitation for this period, which is compared to the mean annual precipitation for the long term reference period. Seasonal variation of precipitation was also evaluated among the years, during 2006-2010, in order to detect seasonal differences among years.
Seasonal periods, based on temperature and precipitation data. A cluster multivariate analysis was applied to a data set of monthly precipitation and temperature, obtained from the nearest weather station to the study site, Isla Aguada (No. 4015), for a five-yr (2006-2010).
In addition, the seasonal pattern of variation of the Isla Aguada was compared to data from 41 other meteorological stations distributed all over Campeche. All data were provided by the Servicio Nacional Meteorológico (National Service of Meteorology) and the Comisión Nacional del Agua (Federal Agency of Water Administration (CONAGUA, 2013).
Yearly and seasonal variation of the precipitation. We used a precipitation data set of Isla Aguada station (4015), for the period 20062010, adding the data from November and December of 2005, and from January of 2011, to complete the seasonal periods. We considered this data set for the repeated measures ANOVA (see below) to know if the precipitation significantly changes on a seasonal and yearly basis.
Ground water chemistry. From 2006 to 2010, with a monthly basis, we measured the salinity and the redox potential of the ground water of the studied mangrove. The redox potential measures the activity of the electrons of the chemicals found in the ground water, and it is specifically related with the oxygen dissolved in water (Hermosilla et al., 2005). Along a 30 m-transect from the coast line of the lagoon to the interior of the vegetation (up to the borderline of the mangrove), we installed three 0.1 m-diameter and 1.5 m length PVC tube collectors at a depth of 50 cm from the sediment surface. These tubes were drilled, with 1 cm diameter holes, at 30 cm depth (at this depth the largest amount of root biomass of trees is usually located). Water samples were collected after the tubes were drained and water was stabilized. Salinity was measured with a refractometer A & O, considering output ranges from 0 to 100 PSU (Practical Salinity Unit) (Agraz-Hernández et al., 2011). Redox potential was measured with a data logger set at multiparametric mode (HACH-HQ11d). The electrodes used at the field was HACH LANGE-MTC101. The HQd meter used a probe (ORP-Redox), which was gel unfilled. The solution ORP-Redox considered intervals of +1200 mV.
Physiognomic type of the forest. In January 2006, we estimated the structure of the mangrove forest through the basal area, density and tree height, using the point quarter method described by Cintrón & Schaeffer-Novelli (1983) for mangroves (trees >2.5 cm diameter). The general procedure consists of tracing a transversal transect, which is divided in 20 points (centered quarter points), from the coast line of the lagoon towards the forest. Each point is the center of Cartesian grid system; in each quadrant the distance to the nearest mangrove tree is measured, as well as the diameter and the height of the tree. From these data, the basal area of the trees and the density for each species is calculated. The modified procedure for mangroves consists of a differential measure of the tree diameter according to the species: this measurement is at a fixed tree height of 1.30 cm in the case of Avicennia germinans, Laguncularia racemosa (Linneo) Gaerth f. and Conocarpus erectus Linneo. If the species is Rhizophora mangle the tree diameter is measured 30 cm above the binding of the oldest aerial roots.
The physiognomic type of the forest was determined through the density, basal area and height of the forest following Flores-Verdugo et al. (1992).
Productivity based on litterfall production. We evaluated the productivity of the mangrove forest in terms of the litterfall production of Rhizophora mangle during a five-yr period (2006-2010), as the total dry weight of litterfall per square meter per year and per season. We randomly installed 14 net litter traps (area of 0.25 m2 each) with 0.1 mm of mesh size in a total area of 0.5 hectare. Traps were hold at their four corners below the tree canopy and above the highest tide level. The collected material was dried in a convection oven at 65°C for approximately three days until constant weight (Agraz-Hernández et al., 2011; Kamruzzaman et al., 2013). Once the material was dried, we separated and weighed the material in branches, leaves, flowers, hypocotiles and stipules. We grouped the data in monthly sets of seasonal and yearly groups to do the statistical analyses. The productivity of the mangrove was estimated as the total litterfall dry weight per square meter per day, and the reproductive phenology was evaluated by the flower and hypocotile production per square meter per day (Agraz-Hernández et al., 2011; Kamruzzaman et al., 2013).
Statistical data analyses. We applied a repeated measures analysis of variance to test the effect of years and seasons on the variation of the precipitation during 2006-2010, the ground water chemical parameters (salinity and redox potential), and the biological data (litterfall and hypocotile production). We did a multiple mean comparison, when needed, with the Fisher's protected LSD test. For this analysis we transformed data to the root-square as recommended by Guisande et al. (2005) to accomplish normality, and used a 0.05 significance level. We applied the Pearson correlation coefficient to correlate the precipitation with the physicochemical and productivity parameters. Finally, a multiple regression model was applied to evaluate the dependence of the productivity (litterfall production) and the hypocotile production on precipitation, salinity and the redox potential. We previously proved the homogeneity of variances with the test of sphericity of Mauchly, and there were non-significant differences between data groups (p>0.05, Table 2 and 3). All statistical analyses were done using STATISTICA (v. 6) and SPSS statistical programs, and procedures criteria followed Zar (2010).
RESULTS
Temporal distribution of the precipitation. We could differentiate three seasons for the study site (Fig. 3): a dry season from February to April, a rainy season from May to October, and a period of intermediate precipitation from November to January, called Nortes season (humidity conditions during this period of time are due to strong cold winds, coming from the Northeast). These results were confirmed with a larger data set from 41 weather stations distributed all over Campeche.
Drought analysis of the study site. The distribution of the precipitation was not homogeneous during the reference period from 1959 to 2010. During this time, the strongest drought occurred from 1960 to 1965 (SPI = -2.81), but other relatively severe droughts occurred from 1968 to 1972, from 2001 to 2007, and from 2009 to 2010. In contrast, the precipitation surpassed the reference average in 1975, 1980, 1992 and 1997 (SPI values of 2.05, 1.89, 2.13, and 1.72, respectively) (Fig. 2).
Compared to the period 1959-2010, there was a dryness trend during the period of study (2006-2010), as negative SPI values show, with the exception of the year 2008, which was a relatively wet year (SPI = 0.93) (Fig. 2). During this period, seasonal precipitation differed among years. In 2009, the SPI was negative in all seasons, particularly, in the rainy season (-0.83), which means that this season was relatively less humid than the previous long term period of reference. In contrast, SPI was positive for this season (1.27) in 2008, which resulted in a wetter year compared to 2006, 2007, 2009 and 2010 (Figs. 2, 3 and 4). During the Nortes season, unexpectedly, the SPI was negative in all years (from -1.0, in 2006, to -1.27, in 2008), which indicates a relatively dry Nortes season for the study period. With respect to the dry season, precipitation was less than the expected in 2006, whereas it increased in 2007 and 2008 (SPI= 1.67 and 1.21, respectively), indicating the presence of a relatively humid dry season during these two years. The SPI was negative in 2009 and 2010 (-0.39 and -0.77, respectively), indicating even more dryness than expected for this season for these two years.
Yearly and seasonal variation of the precipitation from 2006 to 2010. Precipitation significantly varied across years and seasons during the period of study (F=15.83, p<0.003; F=29.8, p<0.0001; Tables 2 and 3). The year 2008 showed a precipitation 1.5 higher than the years 2006, 2007, 2009 and 2010 (1,550 mm yr-1, compared to 965, 1,150, 1,000 and 1,107 mm yr-1, respectively). Considering only the rainy season, 2008 showed an extremely high precipitation (1,415 mm), this value surpassed the precipitation of the rest of the years during the period of study (952 mm), and the year 2009 showed the lowest precipitation during this season (770 mm). In contrast, 2008 showed the least amount of precipitation in the Nortes season (49 mm), whereas in 2009, the precipitation of the Nortes season was the highest (195 mm). The dry season for the period of study had a mean value of 64 mm, but it had a high degree of variation among the years (s= 63 mm) (Fig. 4; Table 4).

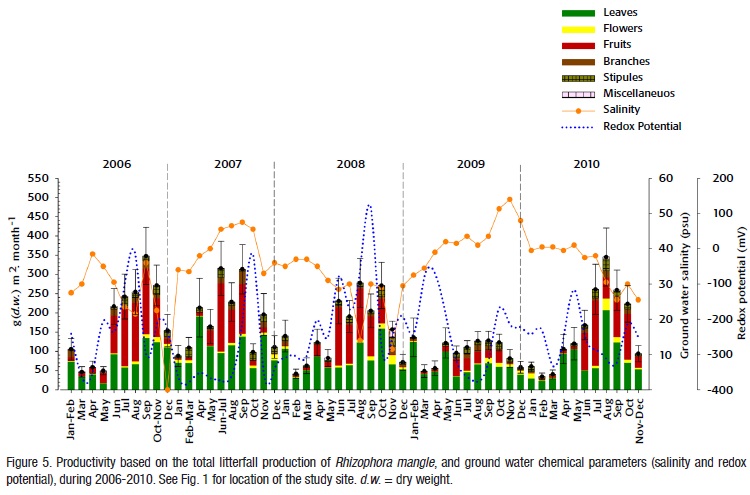
Ground water chemistry. Salinity and the redox potential of the ground water significantly varied among years (F=1003.25, p<0.0001; F=213.32, p<0.0001; Table 2) and among seasons (F=888.2, p<0.0001; F=164.65, p<0.0001; Table 3) during the period of study.
Reduced conditions and the highest salinity were recorded in 2007 and 2009 (40.0 ± 7.0 PSU, -300 ± 119 mV; and 42 ± 5.0 PSU, -279.0 ± 115 mV; respectively), for this reason, these two years were categorized as mesohaline-euhaline.
In 2007, there was a direct correlation between the precipitation and the redox potential of the interstitial water (r=0.914, p<0.05), and an inverse correlation between salinity and the redox potential (r=-0.692, p<0.05) in 2009. Salinity increased in both rainy and dry seasons in 2009 (41.4±7.8 and 42.8±3.9 PSU, respectively), and there were anoxic conditions all the year (from -254.9 to -371.0 mV for all the seasons). In this year, salinity was high during both the rainy season (43.1±1.6 PSU) and the Nortes season (41.8±8.6 PSU), and the redox potential was low in the rainy season (-326.5±72.2 mV), and the Nortes season (342.5±29.8 mV). Indeed, these two parameters were inversely correlated (r=-0.696, p<0.02) (Fig. 5). In contrast, salinity conditions in 2008 were different, showing oligohaline-mesohaline characteristics (28.0±8.0 PSU; -163±136 mV), because salinity decreased and the oxygenic conditions increased, compared to 2007. In 2008, there was a high correlation between the redox potential and the precipitation (r=0.692, p<0.009). Salinity of the ground water was low for both the rainy and the Nortes seasons (25.9±6.2 and 26.2±13.3 PSU, respectively). In 2006, salinity conditions were considered mesohaline (30±6.0 PSU, Fig. 5).In 2010, precipitation increased during the rainy and Nortes seasons (contrasting with 2009) favoring mesohaline and oxygenic-hypoxic conditions. During the dry season, conditions were mesohaline and hypoxic (40±1 PSU and -325±8 mV).
During the overall period, the water body was characterized as mesoha-line (35.1 + 8.4 PSU), oxygenic with a trend to be hypoxic (-247.2+117.4 mV) (Fig. 5). There was a direct correlation among the precipitation and the redox potential of the ground water (r=0.44, p<0.003), and an inverse correlation between salinity and the redox potential (r = - 0.5, p<0.001), for the whole period.
Physiognomic type of the forest. We characterized the mangrove forest of this study as a monospecific fringe type with a riverine trend (Flores Verdugo et al., 1992) composed of Rhizophora mangle, with a density of 1,400 trees ha-1, 24 m2 ha-1 of basal area, and 6.6 m of mean height.
Forest productivity based on total litterfall production. Total litterfall production of Rhizophora mangle was significantly different across years and seasons (F=39.35, p<0.001; F=58.83, p<0.0001; Tables 2 and 3). The interaction among the two factors was non-significant (F=1.2, p >0.29). Forest productivity varied from 1160.0 to 1838.3 g m-2 yr-1 during the period 2006-2010, showing a maximum peak in 2007 and a minimum value in 2009 (Fig. 5).
The litterfall distribution within a year showed a similar pattern across the years, showing the highest peaks in the rainy season (706.6 to 1,260.0 g m-2) and the least values in the dry season (180.5 to 362.2 g m-2), but 2009 differed from the rest of the years, because it showed lower values of productivity in all months and lower differences among months (Annual litterfall: 1,159.0 g m-2 yr-1; rainy season: 706.6g m-2; Nortes season: 212.0g m-2; dry season: 362.8 g m-2) (Fig. 5); this different pattern of seasonal variation probably contributed with the significance of the season (Table 3).
During the period of study, the leaf fraction of the litterfall was the largest among all fractions (53%, 953.0 g m-2 yr-1). Similarly than the total litterfall, leaf litterfall varied significantly across years and seasons (F=75.17, p<0.0001; F=108.52, p<0.0001; Tables 2 and 3, Fig. 5). The biggest peaks of leaf production occurred in 2007 and 2010 (1, 147.4 and 974.0 g m-2 yr-1, respectively). The leaf production was always high in the rainy season and low in the dry season (Figs. 4-5). The largest values of leaf production occurred in the rainy season in 2007 (521.6 g m-2, 46.5%), 2008 (467.1 g m-2, 60.1%) and 2010 (639.3 g m-2, 57%).
Reproductive phenology and hypocotile production. We could detect differences in flower production among years (F=12.16, p<0.013; Table 2); the largest production of flowers was in 2008 (200.6 g m-2yr-1) and, in 2010 (466.5 g m-2 yr-1), and the lowest production occurred in 2006 (141.9 g m-2 yr-1) (Fig. 5). Differences in flower production among seasons were significant (F=67.23, p<0.001; Table 3). Although flower production occurred in all the months from 2006 to 2010, the largest peak of production was in the rainy season and the beginning of the Nortes season (November and December) in those years (90.1 and 301.2 g m-2 in the rainy season of 2009 and 2010, respectively; 79.2 and 101.5 g m-2 in the Nortes season of 2007 and 2008, respectively). The lowest flower production occurred in the dry season of 2009 (8.9 g m-2), which was 62.2% less than the flower production in 2008 (Fig. 5).
There were significant differences in the hypocotile production across years and seasons (F=8.74, p<0.025; F=22.87, p<0.001; Tables 2 and 3). Flowers were produced in all months of the year, however, they were more abundant in the rainy season in all years. The highest production of hypocotiles occurred in 2006 (78.1 g m-2 yr-1) and 2008 (64.2 g m-2 yr-1), and the lowest production was in 2009 (19.1 g m-2 yr-1) (Fig. 5). During the Nortes season, this production drastically decreased from 2007 to 2008, being almost absent in 2009. In the dry season of 2009, contrary to the expected, hypocotiles were produced; actually, we obtained a relative high correlation between the hypocotile production and the seasonal precipitation of the dry period for this year (r=0.60, p<0.05). The precipitation, the salinity and the redox potential values were significantly correlated with the production of hypocotiles (r=0.37, p<0.017; r=-0.36, p<0.02, and r=0.43, p<0.005; respectively). Rhizophora mangle showed the largest production of hypocotiles during the rainy season and the beginning of Nortes in 2006 and 2008 (63.01and 56.11 g m-2), contributing with the 68.6% of the total litterfall. In fact, we found a high direct correlation between the potential redox values and the hypocotile production (r=0.64, p<0.025) and an inverse correlation with salinity (r= -0.6, p<0.036). The lowest production of hypocotiles was recorded in the rainy season of 2009 (17.6 g m-2), decreasing in 68% with respect to the previous year (2008). The hypo-cotile production showed a high direct correlation with the precipitation (r=0.60, p< 0.0061), and also with the redox potential values (r = 0.53, p<0.05). Hypocotile production drastically reduced in the rainy season of 2007 (46.7 g m-2). Its production was the 16% of the total reported in 2008, and the peaks of the production were in July and September (Fig. 5); the lowest production of hypocotiles occurred in the rainy season in 2009 (0.4 g m-2), even though precipitation during the Nortes season of this year surpassed the precipitation during the same season in 2008.
Prediction models for the litterfall and the hypocotile production. The multiple regression model obtained for the litterfall production is described as follows (R2=0.85; p<0.05):
y (litterfall production) = 7.5982+ 0.00944121x - 0.1241183z
where x (annual precipitation in mm), and z (salinity of the ground water in PSU) are significant factors that determine the variation of the response variable y (litterfall production in g m-2 day-1).
The multiple regression model obtained for the hypocotile production is described as follows (R2=0.70; p<0.05):
y (hypocotile production)= 4.63994 + 0.00617553x - 0.101467z
where x (annual precipitation in mm) and z (salinity of the groundwater in PSU) are significant factors of the variation of the response variable y (hypocotyle production in g m-2 day-1).
DISCUSSION
Precipitation regime during the period of study (2006-2010). During 2006-2010, except for 2008, the study site was relatively drier compared to the long term reference period 1959-2010. According to Mendez & Magaña (2010) Mexico is extremely vulnerable to droughts because of its geographic location. Since water availability in the North of Mexico is scarce, the impacts of droughts in this region are considered more severe than in the South, in consequence, they have been more deeply analyzed from the socioeconomic perspective (Magaña & Conde, 2003). However, dry and wet episodes are also part of the natural climatic variability in both central and southern México.
Considering the seasonal regime of the precipitation, the rainy seasons of the years 2006 and 2007 were less abundant compared to the reference long term period (1959-2010), and precipitation notably increased by 2008, whereas it decreased again in 2009, and stabilized in 2010 (Fig. 4). The El Niño-South Oscillation (ENSO) phenomenon may play a determinant role in causing variation among seasons between two-seven yr-periods (Mendez & Magaña, 2010). Also, Drexler & Ewel (2001) have demonstrated that short-term climatic disturbances such as ENSO, can influence important coastal process. Over repeated drought cycles, such disturbances have the potential to affect the structure and function of mangrove forests and upstream ecosystems. According to the National Oceanic and Atmospheric Administration (NOAA, 2013) the year 2008 showed the cold phase of ENSO, i.e. El Niño, and La Niña, the warm phase of this phenomenon, occurred in 2009.
The year 2009 was particularly dry in all seasons (Fig. 5; Table 4); Vega & Miranda (2009) refer to 2009 as the year with the most severe droughts in Mexico for the last 50 years. Actually, these droughts had negative effects on agriculture, damaging, among others, the corn crops in a 55%, and the bean crops in a 22%.
The rainy season of the year 2008 had a rainfall 46% higher than the mean precipitation for this season during the period 2006-2010. The year 2009 showed an opposite pattern, during this year, seasonal mean precipitation corresponding to the Nortes, the dry and the rainy seasons were below the mean precipitation for the entire period 2006-2010 (25%, 17% and 21%, respectively). Zhang et al. (2013) indicate there is growing evidence that precipitation regimes have become more extreme at global, regional and local scales. Such altered intra-annual precipitation patterns, and warmer temperatures, may affect the productivity and the water balance of the ecosystem, which results in changes of the structure and function of terrestrial ecosystems.
Ground water chemistry variation during the period of study (2006-2010). There was a significant yearly and seasonal variation of the chemical parameters of the ground water during the period of study, accordingly to the precipitation regime for the same period (Tables 2 and 3). The ground water globally showed a mesohaline and oxygenic (with a trend to be hypoxic) environment, but, particularly the years 2007 and 2009 showed the high estrange of variation, salinity increased (in a magnitude of 10 PSU from 2006 to 2007) and the redox potential decreased, making the chemical environment more saline and hypoxic (extreme anoxic conditions of the ground water was recorded in the dry and rainy seasons of 2007). As a result of a higher precipitation in 2008, however, conditions of the ground water environment turned to be oxygenic, favoring the availability of dissolved oxygen. Correlations between the precipitation and the environment variables of the ground water confirm this trend. Whenever the precipitation was higher, it was associated with an increase in the redox potential of the ground water (r=0.914; p<0.05); a lower salinity in the ground water was associated with an increase in its redox potential (r= -0.692; p<0.009). According with these results, Saravanakumar et al. (2008) found a high inverse correlation of the oxygen concentration of the ground water on salinity and temperature in mangroves of the Indian coast. The Intergovernmental Panel on Climate Change (Houghton et al., 2001) has reported that a reduction in the precipitation causes a decrease in the contribution of fresh water, which makes the soil more saline. Supporting this idea, Gilman et al. (2008) refer that an increase in salinity conditions can cause a severe reduction in the mangroves surface because of the conversion of upper tidal zones to hypersaline flats.
When salinity conditions of the ground water become extremely high, there are detrimental effects on mangrove individual trees, such as the inhibition of photosynthetic enzymes, and consequently a reduction in the CO2 assimilation rate. Also, the transpiration and filtration processes may be affected as a result of an excess of salts inside the mangrove plants. Rhizophora mangle is especially vulnerable when salinity of the ground water surpasses the 30 PSU because this species does not have any desalination glands, nor it accumulates salts in the vacuoles (Saenger, 2002), for this species, the elimination of salts demands a high energy investment. With the exception of the year 2008, salinity conditions of the ground water was lightly higher 30 PSU, which means the R. mangle was able to tolerate these conditions, but probably with higher costs in terms of growth and reproductive output.
Dominance of Rhizophora mangle in Laguna de Términos. The study site receives fresh water rich in nutrients and a constant contribution and transport of sediment, coming from the southeastern rivers Mamantel and Balchancah (at 34 and 31km from Isla Aguada, the nearest weather station to the study site, respectively); this geographical situation, together with the runoffs in the rainy season, and the contribution of tides, have created ideal conditions for the establishment of a monospecific mangrove forest of R. mangle. This species usually tolerates moderate to high saline conditions in the ground water. Day et al. (1987) have reported monospecific mangrove forests of R. mangle dominating the whole area surrounding Laguna de Términos. The mesohaline and oxygenic conditions of the ground water have created an optimal environment for the development and growth of R. mangle. Parallel, Agraz-Hernández et al. (2011) reported a good development of mangrove forests, dominated by R. mangle, along the border of the lagoon system La Mancha, in Veracruz, with a relatively low salinity in the ground water (2.5 to 23 PSU).
Compared to other mangrove species, R. mangle is very sensitive to changes in the hydroperiod, nutrient availability, dissolved oxygen in the ground water, and salinity. Psuty et al. (2009) mention for example that Avicennia germinans only develops in areas with scarce overflows and nutrient availability, whereas Laguncularia racemosa, is frequently found in overflowed terrains with high nutrients. In this sense, R. mangle may be considered as an indicator of the inter-annual and seasonal variation of the precipitation, because this species can tolerate a wider range of environment conditions of the ground water. At the present, R. mangle covers a large area in Laguna de Términos, however, Mitsch & Gosselink (1993) predict that R. mangle could be replaced by L. racemosa and A. germinans if hypoxic conditions of the ground water and a high salinity become frequent conditions, and longer residences of surface waters occur.
The variability in the precipitation regime may also be affecting survival of mangrove seedlings in Laguna de Términos (Lugo, 1986; Krauss et al., 2008). These authors agree with the idea of possible future changes in species composition and zonation of mangrove forests in the region.
Productivity of the monospecific mangrove of Rhizophora mangle. Litterfall production of the studied mangrove forest, during the period 2006-2010, ranged from 1,160.0 to 1,838.3 g m-2 yr-1, showing a maximum peak in 2007 and the lowest production in 2009 (Fig 4). Day et al. (1987), who studied a mixed mangrove forest of R. mangle, L. racemosa and A. germinans in a nearby region of Laguna de Términos, obtained a litterfall production of 1,252.0 g m-2 yr-1, value that falls in the range of the results of this study. Laguna de Términos has regular contributions of nutrients, from abundant fresh water from the local rivers, condition that must play an important role in determining the high productivity values obtained for the monospecific mangrove forest we studied. Nevertheless the productivity of this forest was relatively high, it significantly varied across years and seasons (Table 2, 3). Precipitation and salinity resulted significant factors driving the litterfall production of the studied mangrove forest of R. mangle. Precipitation contributed with the 57% of the variation of the litterfall production (Figs. 3-4; Table 4). According to Xiao & Moody (2004) tree water balance is a primary determinant of the spatial heterogeneity in terms of litterfall production.
In our study, the biggest peak of litterfall occurred in 2007, probably because the dry season extended to May in this year. Supporting this idea, Snedaker (1995), refers that, occasionally, a high defoliation of this species may occur when the dry season is extreme and long. As a consequence of a relatively drier environment, in 2007, the ground water environment turned to be hyperhaline and hypoxic (Fig. 4). Lichtenthaler (1996) and Saenger (2002) have reported that R. mangle is able to survive under extreme salinity conditions and hypoxia, because this species administrates well its energy budget to maintain vital physiological processes at a basal level, when the conditions are adverse.
Litterfall production was similar in 2006 and 2008, and it rapidly recovered from 2009 to 2010. These results demonstrate a resilient response of R. mangle to the variability in the precipitation regime. Nevertheless the extreme conditions that occurred in 2007, the litterfall production recovered in the subsequent year, 2008, since atmospheric conditions and the ground water environment became more favorable (Figs. 4-5).
There were differences in the litterfall production among seasons during the period 2006-2010, however, the production peak was always during the rainy season and the lowest production occurred was in the dry season (Fig. 5). Some authors have found similar results in other mangrove forests, detecting the highest peaks of production for the genera Avicennia, Bruguiera, Sonneratia and Rhizophora, in the rainy season (Wium-Andersen & Christensen, 1978; Duke, 1988). Some important seasonal differences occurred among years, in 2009, for example, litterfall production was relatively low (there was a litterfall loss of a 37%), reflecting the reduction of the precipitation during this year and the increase in the salinity of the ground water increments because the dry season was longer (Fig. 5; Table 4).
In 2010, in contrast, the rainy season prolonged, returning the litterfall production to the productivity values registered in 2008. During this season, there was a higher contribution of fresh water and nutrients, a lower salinity, and oxygenic conditions in the ground water (Fig. 5). Similarly, Chen et al. (2010), studying a mangrove forest of Sonneratia caseolaris (Linneo) Engl. in China, found a direct correlation between the litterfall production and the precipitation, during a ten-yr period of study.
Reproductive phenology of the monospecific mangrove forest of Rhizophora mangle. Flower production of R. mangle significantly varied across years and seasons during the period 2006-2010 (Tables 2-3). The most abundant production of flowers was in the rainy and the Nortes seasons. Similar results were obtained by Aké-Castillo et al. (2006) in the lagoon system Laguna de Sontecomapan, in Veracruz, Mexico, and by Barreiro-Güemes (1999), in the estuarine system Pom-Atasta, in Laguna de Términos. In 2006 and 2007, the peak of flower production was in the rainy season, whereas in 2008, it occurred in the Nortes season. During these periods a low concentration of salts dominated in the ground water. Xiao & Moody (2004) indicates that the seasonality of precipitation is an important determinant of the soil-water budgets, which have a direct influence in the productivity of different vegetation types.
We recorded the biggest peaks of hypocotile production in the rainy season. In this season, salinity is low and the nutrients are more available as well as the dissolved oxygen in the ground water (Fig. 5). Agraz-Hernández et al. (2011) indicates that R. mangle maintains the flowering and propagule production all the year, however, the highest litterfall and hypocotile production always occur in the rainy season because the oxygen and nutrients availability in the ground water is higher in this season, whereas salinity decreases (also, Navarrete & Oliva-Rivera, 2002; Barreiro-Güemes, 1999; and Aké-Castillo et al., 2006). Some authors have reported that R. mangle commonly produces flowers and propagules all the year (Aké-Castillo et al., 2006; Agraz-Hernández et al., 2011). In this study this behavior was present only in 2006 and 2008, when the precipitation was more abundant. In 2007 and 2009, the precipitation decreased, creating hypersalinity and hypoxic conditions, therefore, the production of hypocotiles was the lowest (the correlation between the redox potential and the production of hypocotiles in 2009 was inverse (Figs. 4-5). Similarly, Chen et al. (2010) obtained a direct correlation between the precipitation and the production of propagules across ten annual cycles in forests of Sonneratia caseolaris.
The hypocotile production was significantly dependent on the precipitation and salinity. Agraz-Hernández et al. (2011) have indicated that although R. mangle reproduce all the year, its most heavy hypocotile production occurs during the rainy season. A number of authors coincide with these results, who point out that this species permanently reproduce during the year, but presents a marked seasonality in the rainy season where salinity is relatively low and nutrient availability is relatively high (López-Portillo & Ezcurra, 1985; Barreiro-Güemes, 1999; Aké-Castillo et al., 2006).
Possible scenarios of change. The patterns of yearly and seasonal variation of the productivity and reproductive phenology observed in the monospecific mangrove forest of R. mangle during the period 20062010, are reflecting the variability of the precipitation regime, which modifies the patterns of variation of the chemical conditions of the ground water. According to the results of our study, we establish three possible scenarios of change for the studied mangrove: 1) when an increase in the precipitation occurs (such as in 2008 and 2010), it may favor the entrance and drainage of fresh water to the forest, increasing the nutrients, the oxic conditions, reducing salinity and making the conditions of the ground water to be acidic; all these conditions may cause a net increase of the litterfall production; 2) If the precipitation decreases (as it occurred in 2009), salt concentration increases, and reducing conditions and nutrient contribution are lower, which may drive to a reduction of the litterfall production; and 3) when dry conditions extends and salinity increases (up to 40 PSU or more) (as it occurred in 2007 and 2009), this may affect the nutrient contribution and the oxygen concentration in the ground water, environment conditions that may drive to a massive defoliation event and a reduction or absence of propagules production.
In agreement with these ideas, Chen et al. (2010), describing the pattern of variation of litterfall production in a forest of a basin type with a dominance of S. caseolaris, during 10 annual cycles (1996-2005), attribute the observed abnormalities to the precipitation regime. Gleason et al. (2003) relates the hydroperiod dynamics with the oxidation status of the ground water, and report an important increase in phosphates and nitrates availability, conditions that promote the production of new photosynthetic tissue, which probably reflects a high leaf litterfall production. Field (1995) and Ellison (2000) indicate that changes in the precipitation regime can originate a climate variation that will affect the growth, and survival of propagules in the mangrove forests. The IPCC (2001) refers that predictive models for mangrove forests of Central America and Australia, indicate that the precipitation may decrease in certain areas as a result of changes in the precipitation regime, which is probably a consequence of the global climate change.
Conclusions and perspectives. During 2006-2010 there was a general trend of dryness, although 2008 was a particularly wet year. By 2010, the mangrove forest could recover from an extreme dry year, 2009, increasing its values of litterfall and hypocotile production. Precipitation significantly varied across years and seasons, as well as salinity and the redox potential of the ground water, and also the productivity and the reproductive phenology of the studied mangrove. Precipitation and salinity were determinants of the total litterfall and hypocotile production of the forest. According to our results, R. mangle is a highly resilient species as it could cope with a high variability in the precipitation and ground environment regimes. More precise studies about the effect of long term variations of climatic variables on both environment ground water and biological responses of this and other mangrove species are needed in order to better understand the actual responses of mangrove forests to the climate variability.
ACKNOWLEDGMENTS
We specially thank the careful revisions made by the anonym reviewers of previous versions of this work that considerably improved it. Diverse Mexican federal agencies and institutions contributed with financial support to this investigation: The Federal Agency of Electricity Administration of the Yucatan Peninsula (Comisión Federal de Electricidad; Residencia General de Construcción y Supervisión de LT y SE Peninsula), the Biodiversity Federal Agency (CONABIO; Comisión Nacional para el Conocimiento de la Biodiversidad en México), Water Federal Agency (CONAGUA; Comisión Nacional del Agua) and the National Council of Science and Technology (CONACYT; Consejo Nacional de Ciencia y Tecnología); project FOMIX-Campeche 126430.
REFERENCE
Agraz-Hernández, C. M., C. García-Zaragoza, S. Iriarte-Vivar, F. J. Flores-Verdugo & P. Moreno-Casasola. 2011. Forest structure, productivity and species phenology of mangroves in the La Mancha lagoon in the Atlantic coast of Mexico. Wetlands Ecology Management 19: 273-293. [ Links ]
Agraz-Hernández, C. M., J. Osti-Sáenz, C. A. Chan Keb, G. Expósito-Díaz, E. Márquez-García, J. Acosta, S. Castillo-Domínguez, D. Gómez-Ramírez, J. E. Reyes-Castellanos, K. P. Conde-Medina, Y. M. Cach-Ruíz & J.G. Martínez-Kumul. 2012. Los ecosistemas de manglar en el estado de Campeche: Diagnóstico de la conservación e identificación de áreas potenciales para la restauración. Informe Final. Fondos Mixtos del Gobierno del estado de Campeche y Ciencias y Tecnología (CONA-CYT-FOMIX). Universidad Autónoma de Campeche. Clave 1264430. 179 p. [ Links ]
Aké-Castillo, J. A., G. Vázquez & J. López-Portillo. 2006. Litterfall and decomposition of Rhizophora mangle L. in a coastal lagoon in the southern Gulf of Mexico. Hidrobiology 559: 101-111. [ Links ]
Álvarez-Arellano, A. D. & K. Gaitán-Morán. 1994. Lagunas costeras y el litoral mexicano: Geología. In: de la Lanza-Espino, G. & C. Cáceres Martínez (Eds.). Lagunas costeras y el litoral mexicano. Universidad Autónoma de California, pp. 13-74. [ Links ]
Barreiro-Güemes, M. T. 1999. Aporte de hojarasca y renovación foliar del manglar en un sistema estuarino del sureste de México. Revista de Biología Tropical 47: 729-737. [ Links ]
Bordi, I. & A. Sutera 2007. A drought monitoring and forecasting at large scale. In: Rossi, G., T. Vega & B. Bonaccorso (Eds.). Methods and Tools for Drought Analysis and Management. Springer, pp. 3-27. [ Links ]
Carranza-Edwards, A. M., M. Gutiérrez-Estrada & R. Rodríguez-Torres. 1975. Clasificación morfotectónica de las costas de México. Anales del Centro Ciencias del Mar y Limnología. Universidad Autónoma de México 2: 81-88. [ Links ]
Chen, L., Q. Zan, M. Li, J. Shen & W. Liao W. 2010. Litter dynamics and forest structure of the introduced Sonneratia caseolaris mangrove forest in Shenzhen, China. Estuarine, Coastal and Shelf Science 85: 241-246. [ Links ]
Cintrón, G. & Y. Schaeffer-Novelli.1983. Introducción a la ecología del manglar. Oficina de Ciencia y Tecnología de la UNESCO para América Latina y el Caribe ROSTLAC. Montevideo Uruguay. 109 p. [ Links ]
CONABIO (Comisión Nacional para el Conocimiento y uso de la Biodiversidad). 2009. Manglares de México: Extensión y distribución. 2 ed. CONABIO. México. 99 p. [ Links ]
CONAGUA. (Comisión Nacional del Agua) 2013. Base de datos CLICOM, Información de variables climatológicas. Disponible en linea: http://smn.cna.gob.mx/index.php?option=com_content&view=article&id=4&Itemid=5 (Consultado el 30 de enero de 2013). [ Links ]
Cook, E. R., D. W. Stahle, & M. K. Cleaveland. 1992. Dendroclimatic evidence from eastern North America. In: Bradley, R. S. & P. D. Jones (Eds.). Climate Since A.D. 1500. London & New York, pp. 331-348. [ Links ]
Coronado-Molina C., H. Alvarez-Guillen, J. W. Day Jr., E. Reyes, B. C. Perez, F. Vera-Herrera & R. Twilley. 2012. Litterfall dynamics in carbonate and deltaic mangrove ecosystems in the Gulf of Mexico. Wetlands Ecology Management 20: 123-136. [ Links ]
Day Jr., W. H. Conner, F. Ley-Lou, R. H. Day & A. Machado-Navarro.1987. The productivity and composition of mangrove forests, Laguna de Términos, Mexico. Aquatic Botany 27 (3): 267-284. [ Links ]
Drexler, J. W. & K. C. Ewel. 2001. Effect of the 1997-1998 ENSO-Relate drought on hydrology and salinity in a Micronesian wetland complex. Estuaries 24 (3): 347-356. [ Links ]
Duke, N. C. 1988. Phenologies and litter fall of two mangrove trees, Sonneratia alba Sm. and S. caseolaris (L.) Engl. and their putative hybrid, S. Gulngai N.C. Duke. Australian Journal of Botany 36: 473482. [ Links ]
Ellison, J. C. 2000. How South Pacific mangroves may respond to predicted climate change and sea level rise. Chapter 15. In: Gillespie, A. & W. Burns (Eds.). Climate change in the South Pacific: Impacts and Responses in Australia, New Zealand and Small Islands States. Kluwer Academic Publishers, Dordrecht, pp. 289-301. [ Links ]
Field, C. D. 1995. Impacts of expected climate change on mangroves. Hydrobiología 295 (1-3): 75-81. [ Links ]
Flores-Verdugo, F. J., F. González-Farías, D. S. Zamorano & P. Ramírez-García. 1992. Mangrove ecosystems of the Pacific coast of Mexico: Distribution, structure, litterfall and detritus dynamics. In: Seeliger, U. (Ed.). Coastal Plants Communities of Latin America. Academic Press, pp. 269-288. [ Links ]
Flores-Verdugo, F. J., P. Moreno-Casasola, C. M. Agraz-Hernández, H. López-Rosas, D. Benítez-Pardo, & A. C. Travieso-Bello. 2007. La Topografía y el Hidroperiodo: dos factores que condicionan la restauración de los humedales costeros. Boletín de la Sociedad Botánica de México 80: 33-47. [ Links ]
García, E. 1981. Modificaciones al sistema de clasificación climática de Kóppen (para adaptarlo a las condiciones de la República Mexicana). Instituto de Geografía, UNAM. 246 p. [ Links ]
Gilman, E. L., J. Ellison, N. C. Duke & C. Field. 2008. Threats to mangroves from climate change and adaptations: A review Aquatic botany. Aquatic Botany 80 (2) 237-250. [ Links ]
Gleason, S. M., K. C. Ewel & N. Hue. 2003. Soil redox conditions and plant soil relationships in a Micronesia mangrove forest. Coastal and Shelf Science 56: 1065-1074. [ Links ]
Guisande, C. A., I. M. Barreiro, Riveiro & A. R. Vergara. 2005. Tratamiento de datos. Universidad de Vigo, Vigo. 277 p. [ Links ]
Hermosilla, Z., C. Jorge, I. Romero, E. Martí & M. A. D. Cabañero. 2005. Diferencias espaciales y estacionales en el contenido de nutrientes, demanda de oxígeno y potencial redox en sedimentos bajo una instalación de producción acuícola en jaulas. Boletín Instituto Español de Oceanográfia 21: 29-35. [ Links ]
Houghton, J. T., Y. Ding, D. J. Griggs, M. Noguer, P. J. van der Linden, X. Dai, K. Maskell & C.A. Johnson (Eds.) 2001. Climate change 2001: The scientific basis. Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, New York. 83 p. [ Links ]
IPCC (Grupo Intergubernamental de expertos sobre el cambio climatico ,(2001). https://www.ipcc.ch/pdf/climate-changes-2001/impact-adaptation-vulnerability/impact-spm-ts-sp.pdf (Consultado el 25 de enero de 2013). [ Links ]
Kamruzzaman, M., Sharman S., Kamara M. & A. Hagihara. 2013. Vegetative and reproductive phenology of the mangrove Bruguiera gymnorrhiza (L.) Lam. On Okinawa Island, Japan. Trees. 27: 619-628. [ Links ]
Krauss, K. W., C. E. Lovelock, K. L. McKee, L. López-Hoffman, S. M. L. Ewe & W. P. Sousa. 2008. Environmental drivers in mangrove establishment and early development: a review. Aquatic Botany 89: 105-127. [ Links ]
Lichtenthaler, H. K. 1996. Vegetation stress: an introduction to the stress concept in plants. Journal of Plant Physiology 148: 4-14. [ Links ]
López-Portillo, J. & E. Ezcurra E.1985.Litterfall of Avicennia germinans L. in a One-Year cycle in a mudflat at the Laguna de Mecoacan, Tabasco. Mexico. Biotropica 17 (3): 186-190. [ Links ]
Lugo, A. E. 1986. Mangrove understory: an expensive luxury?'Journal of Tropical Ecology 2 (3): 287-288. [ Links ]
Magaña, V. & C. Conde. 2003. Climate variability and climate change and their impacts on the freshwater resources in the border region: A case study for Sonora, Mexico. In: Diaz, H. F. & B. S. Morehouse (Eds.). Climate and Water-Transboundary Challenges in the Americas. Kluwer Academic Publishers, pp. 373-393. [ Links ]
McKee, T. B., Doesken, N. J, Kleist, J., 1993. The relationship of drought frequency and duration to time scales. In: American Meteorological Society (Ed.). Manuscripts of the 8th Conference on Applied Climatology. AMS, AAG, WMO, pp. 179-184. [ Links ]
Méndez, M. & V. Magaña. 2010. Regional aspects of prolonged meteorological droughts over Mexico and Central America. Journal of Climate 23: 1175-1188. [ Links ]
Mitsch, W. J. & J. G. C. Gosselink. 1993. Mangrove wetlands. In: Mitsch, W. J. & J. G. Gosselink (Eds.). Wetlands (2nd edn.). Van Nostrand Reinhold, New York, pp. 293-328. [ Links ]
Nagarajan, R. 2009. Drought Assessement. Springer. India. 424 p. [ Links ]
Navarrete, A. J. & J. J. Oliva-Rivera. 2002. Litter Production of Rhizophora mangle at Bacalar Chico, Southern Quintana Roo, México. Colegio de la Frontera Sur-Unidad Chetumal, Quintana Roo, México. Universidad y Ciencia 36 (18): 79-86. [ Links ]
NOAA (National Oceanic and Atmospheric Administration). 2013. Disponible en línea en: http://www.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ensoyears.shtml. (Consultado el 26 enero 2013) [ Links ]
Psuty, N. P., L. M. Martínez, J. López-Portillo, T. M. Silveira, J. G. García-Franco & N. A. Rodríguez. 2009. Interaction of a long-shore sediment transport and habitat conditions at Laguna La Mancha, Veracruz, Mexico. Journal of Coastal Conservation 13: 77-87. [ Links ]
Rodríguez-Herrera, H. & L. Bozada-Robles. 2010. Vulnerabilidad social al cambio climático en las costas del Golfo de México: un estudio exploratorio. In: Botello, A.V., S. Villanueva Fragoso, J. Gutiérrez & J. L. Rojas-Galaviz (Eds.). Vulnerabilidad de las zonas costeras mexicanas ante el cambio climático (segunda edición). Universidad Autónoma Metropolitana Iztapalapa, UNAM-ICMYL, Universidad Autónoma de Campeche, pp. 583-624. [ Links ]
Rossi, G., T. Vega & B. Bonaccorso. 2007. Methods and Tools for Drought Analysis and Management. Springer, Dordrecht, The Netherlands. 418 p. [ Links ]
Saenger, P. 2002. Mangrove ecology, silviculture and conservation. Kluwer Snedaker, S. C. 1995. Mangroves and climate change in the Florida and Caribbean region: scenarios and hypotheses. Hydrobiología 295:43-49. [ Links ]
Thom, B. G. 1969. Problems of the development of the Isla del Carmen, Campeche, Mexico. Geomorphologie, Neial Tolge 13 (4): 406-413. [ Links ]
Toledo-Ocampo, A. 2005. Marco Conceptual: Caracterización ambiental del Golfo de México. In: Botello, A. V., J. Rendon-von Osten, G. Gold-Bouchot& C. Agraz-Hernández (Eds.). Golfo de México: Contaminación e Impacto Ambiental: Diagnostico y Tendencias. 2da. Edición. Univ. Autón. De Campeche, Univ. Nal. Autón. De México, Instituto Nacional de Ecología, pp. 25-52. [ Links ]
Vega, M. C. & S. Miranda. 2009. Efectos de la sequía en la producción sembrada a nivel nacional en 2009. Revista Económica Actual (4): 29-31. [ Links ]
Wium-Andersen, S., & B. Christensen. 1978. Seasonal growth of mangrove trees in South Thailand II. Phenology of Bruguiera cylindrical, Ceriop stagal, Lumnitzera littorea, and Avicennia marina. Aquatic Botany 5: 383-390. [ Links ]
Xiao, J. & A. Moody. 2004. Photosynthetic activity of US biomes: responses to the spatial variability and seasonality of precipitation and temperature. Global Change Biology 10:437-451. [ Links ]
Zar, J. H. 2010.Biostatistical Analysis. 5th edition. Pearson Prentice Hall, Upper Saddle River , NJ , USA . 944 p. [ Links ]
Zhang, U., S. M. Moran, M. A. Nearing, G. E. Ponce-Campos, A. R. Huete, A. R. Buda, D. D. Bosch, S. A. Gunter, S. G. Kitchen, W. H. McNab, J. A. Morgan, M. P., M. P. McClaran, D. S. Montoya, D. P. C. Peters & P. J. Starks. 2013. Extreme precipitation patterns and reductions of terrestrial ecosystem production across biomes. Journal of Geophysical Reserch 118 (1):148-157. [ Links ]


{kind=link}