










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.23 no.3 Ciudad de México Set./Dez. 2013
Artículos
Effects of temperature and salinity during the embryonic period of Chirostoma humboldtianum and Chirostoma riojai (Atherinopsidae) until hatching
Efectos de temperatura y salinidad durante el período embrionario de Chirostoma humboldtianum y Chirostoma riojai (Atherinopsidae) hasta la eclosión
María Cecilia Hernández-Rubio1 and Gerardo Figueroa-Lucero2
1Departamento de Zoología, Escuela Nacional de Ciencias Biológicas. Instituto Politécnico Nacional. Prolongación de Carpio y Plan de Ayala. Santo Tomás. México, D.F., 11340. México. E-mail: cecheru@yahoo.com.mx
2Departamento de Hidrobiología. DCBS. Universidad Autónoma Metropolitana-Iztapalapa. Av. San Rafael Atlixco 186, Colonia Vicentina. México, D. F., 09340. México.
Recibido: 10 de septiembre de 2012.
Aceptado: 11 de Julio de 2013.
ABSTRACT
Effects of incubation temperature (18, 20, 22 and 24 °C) and salinity (0, 4, 8, 12, 16, 20 and 24 g/L) on survival, development time and hatching size, during the embryonic period until hatching, of two freshwater endemic Mexican aterinopsids: Chirostoma humboldtianum (Valenciennes, 1835) and Chirostoma riojaiSolórzano and López (1965) were investigated. Embryos of both species did not survive at salinities up to 12 g/L. Hatching percentage was the highest at 22 °C and 0 g/L salinity, in both species (87%) and the lowest at 18° and 4-8 g/L in C. humboldtianum (39.95%) and at 24 °C and 12 g/L in C. riojai(33.23%). Hatching times were inversely proportional to incubation temperature increase; it was 15.5 ± 0.26 days in C. humboldtianum, and 12.4 ± 0.31 days in C. riojai, at 18 °C. At 24 °C, this period was 9.8 ± 0.11 and 7.8 ± 0.24 days, respectively. Hatching size was larger on high temperature and smaller on high salinity. The optimal temperature-salinity combination for hatching size was 24° C and 0 g/L in C. humboldtianum and 24 °C and 4 g/L in C. riojai. There was an interactive effect of temperature and salinity on percentage hatching and on hatching size. Although these species have low salinity tolerance, the use of salt has prophylactic uses for controlling pathogens sensitive to salt. C. humboldtianum and C. riojai are potential candidates for aquaculture for restocking purposes.
Key words: Atherinopsidae, embryo, silverside, survival, tolerance.
RESUMEN
Los efectos de la temperatura (18, 20, 22 y 24 °C) y la salinidad (0, 4, 8, 12, 16, 20 y 24 g/L), fueron evaluados sobre la sobrevivencia, tiempo de desarrollo y talla de eclosión, de embriones de dos aterinópsidos endémicos dulceacuícolas mexicanos: Chirostoma humboldtianum (Valenciennes, 1835) y Chirostoma riojai Solórzano y López, 1965. Los embriones de ambas especies no sobrevivieron en salinidades superiores a 12 g/L. El mayor porcentaje de eclosión se obtuvo a 22 °C y 0 g/L de salinidad, en ambas especies (87%) y el más bajo a 18 °C y 4-8 g/L en C. humboldtianum (39.95%) y 24 °C y 12 g/L en C. riojai(33.23%). Los tiempos de eclosión disminuyeron conforme el incremento de la temperatura, desde 15.5 ± 0.26 días para C. humboldtianum y 12.4 ± 0.31 días en C. riojai a 18 °C, hasta 9.8 ± 0.11 y 7.8 ± 0.24 días, respectivamente, a 24° C. La talla de eclosión aumento con el incremento de la temperatura y disminuyó con salindades altas. La combinación óptima de temperatura-salinidad para la talla de eclosión fue 24° C-0 g/L en C. humboldtinum y 24 °C-4 g/L en C. riojai. Se presentó un efecto interactivo entre la temperatura y la salinidad en el porcentaje y la talla de eclosión. Aunque estas especies son poco tolerantes a la salinidad, el uso de la sal tiene efectos profilácticos para controlar patógenos sensibles a este. C. humboldtianum y C. riojai, son candidatos potenciales en acuacultura para propósitos de repoblación.
Palabras clave: Atherinopsidae, embrión, sobrevivencia, tolerancia.
INTRODUCTION
Life history of fish has a series of crucial developmental events, which are affected by extrinsic and intrinsic factors. Among the extrinsic factors, temperature is the factor with the greatest effect on development, growth and survival. Temperature effects on embryonic period may have an impact later in life history of fish (Wootton, 1990; Kamler, 1992).
Higher temperature values that are within species tolerance interval often induce abnormalities, particularly in the embryonic and larval periods. These abnormalities may affect the efficiency with which both the embryo uses the yolk and how food is used in subsequent developmental periods (Pavlov & Moksness, 1997; Tucker, 1998; Sfakianakis et al., 2004). Fish development is also associated with changes in temperature. If the temperature tolerance range at which a species can live is wide, it becomes significantly shorter for each stage of development (Nikolsky, 1963). During the endogenous feeding period the only source of energy is the yolk sac; therefore both survival and yolk utilization efficiency define the optimum temperature range (Kamler et al., 1998).
Salinity is another extrinsic factor that also affects fish development. For those species that live in water with fluctuating salinity, or which move between bodies of water with different salinities, salinity acts as a masking factor, influencing osmotic and ionic regulation. It also acts as a directive factor in movement for those species that migrate. In euryhaline species, salinity preferences can change with the physiological and developmental stage fish (Holliday, 1969; Wootton, 1990).
Moreover, salinity reduces stress on fish during handling (Haswell &Thorpe, 1982), acts as a prophylactic agent (Altinok & Grizzle, 2001; Figueroa-Lucero et al., 2004; Hernández-Rubio et al., 2006) and increases survival and motility of the sperm, resulting in higher reproductive success (Elofsson et al., 2003).
In this study, it is recognized the genus Chirostoma in agreement to results of Bloom (Bloom et al., 2009, 2013). Genus Chirostoma Swainson includes endemic species, with a marine common ancestor, that inhabit lentic bodies of the Mesa Central in Mexico, although three taxa, C. jordaniWoolman 1894, C. humboldtianum (Valenciennes 1835) and C. mezquital Meek 1904, extend beyond this region (Barbour, 1973; Miller et al., 2005). C. humboldtianum is the species most similar to this ancestor and reaches a length of 200-250 mm; C. riojaiSolórzano & López 1965, is a species with primitive features and reaches a length of 74 mm (Barbour, 1973). These species, like others in this genus, are commercially fished and traditionally consumed in Mexico. They have aquaculture potential; however, the aquaculture success is poor because of the limited knowledge regarding factors, which affect their early development.
In recent years, populations of these species have been markedly affected by pollution and habitat alteration (Soto-Galera et al., 1998; Ford et al., 2000). In consequence, C. humboldtianum populations are endangered and C. riojai is in risk of extinction (Mexican Official Standard NOM-059-SEMARNAT-2010 ) and it must be considered Critically Endangered because is sensitive to environmental degradation and its range is now less than 10 km2 and previously contiguous populations are now severely fragmented (Soto-Galera et al., 2008). This is the first study to examine salinity-tolerance/effects in these species. The aim of this paper was to determine temperature and salinity effects and their combined effects on timing, survival and size at hatching of both species, in order to determine optimum levels of these factors during embryonic period for maximizing free embryos survival.
MATERIALS AND METHODS
Broodstocks of Chirostoma humboldtianum from Las Tazas dam and broodstocks of C. riojai from San Jacinto dam, both in Mexico State, Mexico, were caught to obtain eggs. Broods of each species were maintained in ponds of 5 m3, at 20 °C. C. humboldtianum adults were fed with juveniles of Heterandria sp Agassiz 1853 (Poecilidae) and C. riojai adults were fed with cladoceran Moina macrocopa (Straus, 1820).
Eggs were obtained by in vitro fertilization (Hernández-Rubio et al., 2006). Broods were anesthetized in benzocaine (8 mg L-1). Ova and semen were placed in water at 5 g L-1 of salinity and 20 °C and were gently mixed for a few minutes. The silverside eggs have filaments to attach to vegetation during natural spawning. These were cut once the perivitelline space had been formed, to avoid proliferation of fungi and bacteria.
Experimental design and rearing conditions. Batches of ten eggs, with three replicates, were placed in 250 mL covered glass with filtered water, at each experimental temperature and salinity. The water of each treatment was replaced every third day, until hatching. Each treatment was one combination of the following salinities: 0, 4, 8, 12, 16, 20 and 24 g L-1 (Sigma, TM) and temperatures: 18, 20, 22 and 24 °C.
Under laboratory conditions, both species spawned between 17 and 21 °C; reproduction was not successful at 15 °C (Figueroa-Lucero et al., 2007); subsequently temperatures rise during C. humboldtianum and C. riojai early development. Therefore, in the present study, the experimental viable temperatures were 18, 20, 22 and 24 °C. With respect to salinity, we chose this range of salinities to test because the salinity tolerance of the eggs in these species is unknown.
Age of embryos was estimated from activation, i.e. the moment when water was added to the ova mixed with milt. Every day, mortality and developmental state of embryos were registered. Percentage hatching, incubation time and size at hatching were registered. Developmental stages were defined as described by Balon (1985). The term embryo is applied to fish developing inside the egg and the term free embryo or eleutheroembryo (ELH) to fish after hatching.
Values for % hatch (pij) were transformed to the arcsine √pij (angular transformation) before analysis. Homogeneity of variances was verified by Bartlett's test before each analysis (Sokal & Rohlf, 1969) and χ2 was no significant in all cases. Temperature and salinity effects on % hatch, incubation time, and on eleutheroembryos (ELH) notochordal length (NL) at hatching of each species, were evaluated through a two-way full factorial ANOVA, with α = 0.05. Duncan's post hoc multiple range test, for equal sample sizes, was used for comparing all pairs of treatments means following ANOVA analyses with α set at 0.05 (Montgomery, 1991). Regressions of % hatch over temperature were performed by least squares iteration and were fitted to polynomial curves, for each species and regressions of % hatch over salinity were fitted to linear functions, y = a + bx, (y = % hatch and x = salinity) (Sokal & Rohlf, 1969).
Developmental time to hatch over temperature was fitted to an exponential function y = a exb by least squares iteration, where y represents time in days to hatch, x represents constant temperature (°C), within the optimum range, as reported in Kamler, (1992). Regressions of notochordal length (NL) over temperature and salinity were performed by least squares methods, where y = NL (mm) and x = temperature (°C) or salinity (gL-1) (Sokal & Rohlf, 1969).
RESULTS
Chirostoma humboldtianum ovum has one to four long filaments (mode = 1) and C. riojai ovum has only one long filament. In both species, filaments are in vegetal pole and involve their surface. Eggs are telolecithal, spherical, and pelagic and their chorion is smooth. C. humboldtianum eggs are 1.36 ± 0.047 mm (n = 100) in diameter and slightly amber colored; C. riojai eggs are transparent and have a diameter 1.24 ± 0.05 mm (n = 100). Mean egg diameter of each species differed significantly (t, p < 0.05).
Percentage of hatching and incubation time. Eggs subjected up to 16 g L-1 of salinity, died 24 h post-fertilization (division phase) and those in 16 g/L-1 of salinity died before hatching, in all the temperatures, in both species. The experimental viable salinities were 0, 4, 8 and 12 gL-1.
Both temperature and salinity had a significant effect on percentage hatching (F3,32, p < 0.001, two-way ANOVA, in each species). Duncan tests demonstrated significant differences among treatments and showed an optimal type curve respect to temperature. At hatching, 87% of ELH survived at 22° C in C. humboldtianum and at 20° C in C. riojai. Percentage hatching was inversely proportional to salinity, survival was the highest at 0 gL-1 (87% in both species) and it was depressed at 4-8 gL-1 salinity in C. humboldtianum (40%) and at 12 gL-1 in C. riojai (33%).
The interaction between temperature and salinity was also significant (F9,32, p < 0.001, two-way ANOVA, in both species) but it explained only 6.3 and 15% of the variance on % hatching, respectively (Figs. 1-2).
Mean time to hatch was inversely proportional to temperature for each species (Fig. 3). This period ranged from 15.6 ± 0.26 (C. humboldtianum) and 12.4 ± 0.31 (C. riojai) days at 18 °C, to 9.8 ± 0.11 and 7.8 ± 0.24 days at 24 °C, respectively. In both species, temperature had the highest effect on mean time to hatch (F3, 32, p < 0.0001, two-way ANOVA). Salinity had a significant effect on time to hatch in C. riojai embryos but explained only 9.5% of the variance. The interaction between factors was significant in mean time to hatch of C. humboldtianum embryos but explained only 2.4% of the variance (F3, 32, p = 0.03).
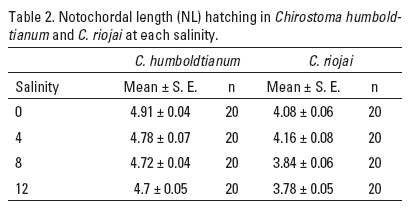
Size at hatching. In both species, higher incubation temperature caused a significantly greater length al hatch (F3, 64, p = 0.003, C. humboldtianum and F3, 64, p < 0.0001, C. riojai, two-way ANOVA) and NL embryos was increased inversely to salinity (F3, 64, p = 0.006, and F3, 64, p < 0.0001, respectively, two-way ANOVA). The interaction between both factors was significant only for C. humboldtianum ELH (F9,64, p = 0.004, two-way ANOVA) (Tables 1-2) (Figs. 4-5).

DISCUSSION
Temperature is the major extrinsic factor controlling the embryonic period of C. humboldtianum and C. riojai. The first lives in water temperature range between 12-25 °C. The second inhabit in water temperature between 8-24 °C (Figueroa-Lucero et al., 2003). In environmental natural conditions, both species spawn all year; the first has a maximum in January-April and June-August (Figueroa-Lucero et al, 2007). The second has a maximum from June to September (Figueroa-Lucero et al., 2003). When the field temperatures increase (march-august), temperatures near shore are higher; it is the place where embryos and larvae live and it is the time when these species are on their early ontogeny (embryonic, larval periods).
In laboratory conditions, both species spawned between 17 and 21 °C; reproduction was not successful at 15 °C. Increased temperature resulted in a significantly spawning rate (Figueroa-Lucero et al., 2007). In the present study, incubation temperature response was adjusted to an optimal type curve, at both the low and high ends (18 and 24 °C) of the temperature range, percentage hatching was lower in both species. These results coincide with the response to this factor for most fish species, at constant temperature. Thermal optima can vary depending on life history and optima for egg incubation may be different from others events during development (Kamler, 1992; Black, 1998).
Salinity effects in early development of fish have been studied in several marine and estuarine species (Holliday, 1969; Middaugh et al., 1986; Phonlor & Sampaio, 1992) but the use of brackish water in freshwater fish culture is not a common practice, despite of cichlids which tolerate a great salinity range (Watanabe et al, 1984), or the coho salmon that tolerate up to 19 g L-1 of salinity (Otto, 1971).
Salinity tolerance can vary in the course of ontogeny. In the osmoregulation, participant hormones and structures are incorporated progressively in the homeostasis of the organism, whose function depends of the environment where it lives and on its development type (Jobling, 1995; Howell et al, 1998).
Chirostoma eggs are pelagic, for what they are affected directly by the environmental factors. Sodium ions, in high concentrations, pass over the chorion and they affect osmotic balance, and in salinities up 12 g L-1, embryos did not survived. Holliday (1969) points there are high mortalities at specific stages of development as at gastrulation and at hatching as response to salinity. It seems to be an inhibitor of chorionasa production. Besides, there are effects in the incidence and concentration of particular ions, the availability of oxygen (the higher the salinity, the lower the oxygen content of the water) and the specific gravities of different salinities may exert an effect through the different buoyancies that the organisms will show. In Odontesthes bonariensis (Valenciennes 1835) eggs, only 14% of the embryos hatched on 5 gL-1 of salt (Nardez et al., 1997). In contrast, there are freshwater fish, which survival and hatching are better in brackish water than in freshwater. Abramis brama (Linnaeus 1758), Lucioperca lucioperca (Linnaeus 1758) and Alosa pontica (Eichwald 1838) hatched in salinities of 10-20 gL-1 of salinity and survival and hatching were better in 2.5 and 5 gL-1 than in freshwater (Holliday, 1969). Survival and hatching of Odontesthes humensis de Buen 1953 were better in 2-10 gL-1 of salinity than in freshwater (Sampaio & Phonlor, 1996). In this study, percentage hatching in C. humboldtianum was 80% at 22 °C and 4 gL-1 of salinity and C. riojai was 83% at 20 °C and 8 gL-1 of salinity.
In the other hand, salinity effect on certain freshwater bacteria and fungi may mean that a slightly enhanced salinity is favorable to developing egg (Holliday, 1969; Sampaio & Phonlor, 1996; Fashina-Bombata & Busari, 2003).
Salinity and temperature have interactive effects on development and should be considered together (Howell et al., 1998). These factors acting together produced the lowest percentage hatching at 24 °C and 12 g/L, and the highest at 22 °C and freshwater. This response has been shown in other species. Parophrys vetulus Girard 1854 had the most rapid rate of development to hatching at 25 gL-1 of salinity and 6-12 °C (Holliday, 1969). Odontesthes bonariensis had a percentage hatching of 46% at 18 °C and 5 gL-1 of salinity, but results were better (88%) with a reduction of dissolved oxygen (1 mg O2.L-1) (Nardez et al., 1997).
Likewise, incubation time was inversely proportional to temperature, in both species. The same pattern has been observed in natural conditions (Ware, 1975). The relationship between both temperature and development time was given by an exponential function, as it has been shown in other species, over the restricted range of favorable temperatures. Development time is more prolonged at low temperatures and decreases with increasing temperature. Temperature is the major controlling factor for developmental rate (Kamler, 1992, 1998). Thus, individuals of same species, under different temperatures, reach a development stage at different times (Margalef, 1980).
It has been observed that larger free embryos (ELH) hatch from larger eggs and smaller eggs give origin to smaller free embryos (Begon et al., 1985; Wootton, 1990; Pepin, 1991; Rideout et al., 2005). Within the atherinopsids, C. humboldtianum is a large species that produces larger eggs (1.36 mm) than those of C. riojai (a small species) and their free embryos maintain this same relationship. However, in other species of the family, this condition is not fulfilled. C. estor estorJordan 1879 is a large species, but its eggs are smaller (1.02-1.16 mm) than those of C. riojai(1.24 mm) and its embryos are larger (4.75 mm) (Estrada, 1991) than those of C. riojai(4 mm). In a close related species to Chirostoma, such as Menidia menidia (Linnaeus 1766) and M. beryllina (Cope, 1867), which have sizes similar to those of C. riojai, the eggs are smaller (1 and 0.8 mm, respectively), but their free embryos are of similar size to those of C. riojai(4 mm) (Wang, 1974).
It has been determined that temperature has a direct effect on yolk utilization efficiency, and that this is higher to low temperatures within the tolerance range of the species and consequently produce larger size at hatch (Ojanguren & Braña, 2003). However, variable results have been obtained in several species (Kamler, 1992). In C. humboldtianum and C. riojai at hatch, NL was reduced with decreased temperature. Similar results have been obtained in other fish species (Kamler, 1992; Pepin et al., 1997). It has been demonstrated that divergence in NL at hatch is due to incubation temperature effect (Kamler, 1992; Fuiman et al., 1998; Pepin et al., 1997; Martell et al., 2005). Kamler (1992) explains that a higher development Q10 than metabolic Q10, will cause the amount of cumulative energy, expended in metabolic processes, decreases with increasing temperature and the remaining energy will be invested in growth of tissue. On the other hand, in C. humboldtianum and C. riojai, both survival and NL at hatch differing in the degree of temperature dependence; the temperature optimum for survival was lower than the temperature at which the largest ELH were obtained.
The differential influence of salinity was manifest in decreased NL at hatch, in higher salinities, in each species. This may be attributed to the non-development of homeostatic mechanism (for salinity adjustment) at this age (Holliday, 1969). A size difference in ELH at hatch, related to salinity has been reported in marine species. Clupea arengus Linnaeus 1758 and Pleuronectes platessa Linnaeus 1758 hatching in salinities of 5-25 gL-1 were up to 23% longer than those hatching in salinities of 35-55 gL-1 (Holliday, 1969). Newly hatched striped bass were longer at 5 gL-1 than at 1 or 10 gL-1 salinity (Peterson et al., 1996). A higher salinity produces shorter individuals. This response is not an osmotic effect only. There are several examples of eurihaline animals, which are longer in lower salinities, owing their higher water content (Margalef 1980; Fridman et al., 2012), however, there are a limited information on salinity tolerance on freshwater fish, specially on the genus Chirostoma.
There was an interactive effect of temperature and salinity upon body size at hatch in C. humboldtianum ELH, only. However, in both species, free embryo NL at hatching, was longer in higher temperatures and lower salinities.
During the endogenous feeding period the only source of energy is yolk, therefore both survival and yolk utilization efficiency define the optimum temperature range (Kamler, 1998). In summary, 20-24 °C and a salinity of 0-8 gL-1, proved to be viable for the embryonic period of C. humboldtianum and C. riojai until hatching, based in on the combination of survival and yolk utilization efficiency.
The present study identified several attributes of C. humboldtianum and C. riojai embryonic period: high survival potential, shorter incubation time and longer size at hatch, at 20-24 °C in fresh and brackish water (0-8 gL-1). Their low tolerance to salinity, it could be useful for controlling pathogens sensitive to salt. These attributes make these species candidates for larval rearing in controlled conditions for restocking purposes.
ACKNOWLEDGMENTS
The authors acknowledge the financial support received from Secretaría de Investigación y Posgrado-IPN (SIP-20111206), CONACyT (130220) and PROMEP (PROMEP-UAM-J-CA-130), Mexico.
REFERENCES
Altinok, I. & J. M. Grizzle. 2001. Effects of brackish water on growth, feed conversion and energy absorption efficiency by juvenile euryhaline and freshwater stenohaline fishes. Journal of Fish Biology 59: 1142-1152. [ Links ]
Balon, E. K. 1985. Early life histories of fishes. New developmental, ecological and evolutionary perspectives. Dr. W Junk Publishers, The Netherlands. 280 p. [ Links ]
Barbour, C. D. 1973. The systematics and evolution of the genus Chirostoma Swainson (Pisces: Atherinidae). Tulane Studies in Zoology and Botany 18 (3): 97-141. [ Links ]
Begon, M., J. L. Harper & C. R. Townsend. 1985. Ecology, Individuals, Populations and Communities. Oxford: Blackwell Publishing. 876 p. [ Links ]
Black, k. D. 1998. The environmental interactions associated with fish culture. In: Black, K. D. & A. D. Pickering (Eds.). Biology of farmed fish Sheffield Academic Press, London, pp. 284-326. [ Links ]
Bloom, D. D., K. R. Piller, J. Lyons, Ν. Mercado-Silva & M. Medina-Nava. 2009. Systematics and biogeography of the silverside Tribe Menidiini (Teleostomi: Atherinopsidae) based on the mitochondrial ND2 gene. Copeia 2: 408-417. [ Links ]
Bloom, D. D., J. T. Eeir, K. R. Piller & Ν. R. Lovejoy. 2013. Do freshwater fishes diversify faster than marine fishes? A test using state-dependent diversification analyses and molecular phylogenetics of new world silversides (Atherinopsidae). Evolution 12074: 1-18. [ Links ]
Elofsson, Η., Β. G. McAllister, D. E. Kime, I. Mayer & B. Borg. 2003. Long lasting stickleback sperm; is ovarian fluid a key to success in fresh water? Journal of Fish Biology 63: 240-253. [ Links ]
Estrada-Rodríguez, M. C. 1991. Verificación a nivel experimental de la existencia de híbridos entre las especies Chirostoma estor estor y Chirostoma grandocule (Pisces: Atherinidae) del Lago de Pátzcuaro, Michoacán, México. Tesis Licenciatura. Universidad Michoacana de San Nicolás de Hidalgo. México. 113 p. [ Links ]
Fashina-Bombata, Η. A. & Α. Ν. Busari. 2003. Influence of salinity on the developmental stages of African catfish Heterobranchus longifilis (Valenciennes, 1840). Aquaculture 224 (1-4): 213-222. [ Links ]
Figueroa-Lucero, g., J. Paulo-Maya & M. C. Hernández-Rubio. 2003. Retrospectiva y avances en el conocimiento de la biología y ecología de los charales y peces blancos del género Chirostoma (Atheriniformes: Atherinopsidae) ENCB-IPN. In: Rojas-Carrillo, P. M. & D. Fuentes-Castellanos (Eds.). Historia y avances del cultivo de pescado blanco. SAGARPA-INP, México, pp. 29-48. [ Links ]
Figueroa-Lucero, g., O. Meza-González, M. C. Hernández-Rubio, I. D. L. Α. Barriga-Sosa, Α. Rodríguez-Canto & J. L. Arredondo-Figueroa. 2004. Growth, survival and mandible development in the larvae of the shortfin silverside Chirostoma humboldtianum (Valenciennes) (Atheriniformes:Atherinopsidae) under laboratory conditions. Aquaculture 242: 689-696. [ Links ]
Figueroa-Lucero, g., M. C. Hernández-Rubio & G. Ontiveros López. 2007. Reproducción en charales y peces blancos. In: Olivera, L. J. I., J. M. T. Jaramillo, H. M. Molina & A. N. P. Téllez (Eds.). Reproducción y manejo de fauna silvestre 3. UAM-Universidad Veracruzana, México, pp. 323-336. [ Links ]
Ford, T. E., R. Ika, J. Shine, L. Lind Davalos & O. Lind. 2000. Trace metal concentrations in Chirostoma sp. from Lake Chapala, Mexico: elevated concentrations of mercury and public health implications. Journal of Environmental Science and Health, Part A 33 (3): 313-325. [ Links ]
Fridman, S., J. Bron & K. Rana. 2012. Influence of salinity on embryogenesis, survival, growth and oxygen consumption in embryos and yolk-sac larvae of the Nile tilapia. Aquaculture 334-337: 182-190. [ Links ]
Fuiman, L. Α., K. R. Poling & D. M. Higgs. 1998. Quantifying developmental progress for comparative studies of larval fishes. Copeia 1998: 602-611. [ Links ]
Haswell, M. S. & G. J. Thorpe. 1982. Millimolar quantities of sodium salts used as prophylaxis during fish hauling. Progressive fish-culture 44 (4): 179-183. [ Links ]
Hernández-Rubio, M. C., G. Figueroa-Lucero, I. D. L. Α. Barriga-Sosa, J. L. Arredondo-Figueroa & T. Castro-Barrera. 2006. Early development of the shortfin silverside Chirostoma humboldtianum (Valenciennes, 1835) (Atheriniformes:Atherinopsidae). Aquaculture 261: 1440-1446. [ Links ]
Holliday, F. G. T. 1969. The effects of salinity on the eggs and larvae of teleosts. In: Hoar, W. S. & D. J. Randall (Eds.). Fish physiology, Vol. I. Academic Press, New York & London, pp. 293-309. [ Links ]
Howell, Β. R., O. J. Day, T. Ellis & S. M. Baynes. 1998. Early life stages of farmed fish. In: Black, K. D. & A. D. Pickering (Eds.) Biology of farmed fish. Sheffield Academic Press, London, pp. 27-66 [ Links ]
Jobling, M. 1995. Environmental biology of fishes. Chapman & Hall. London. 455 p. [ Links ]
Kamler, E. 1992. Early life history of fish: an energetics approach. Chapman & Hall. London. 267 p. [ Links ]
Kamler, E., Η. Keckeis & E. Bauer-Nemeschkal. 1998. Temperature-induced changes of survival, development and yolk partitioning in Chondrostoma nasus. Journal of Fish Biology 53: 658-682. [ Links ]
Margalef, R. 1980. Ecología. Barcelona: Omega. 236 p. [ Links ]
Martell, D. J., J. D. Kieffer & E. Α. Trippel. 2005. Effects of temperature during early life history on embryonic and larval development and growth in haddock. Journal of Fish Biology 66: 1558-1575. [ Links ]
Middaugh, D. P., M. J. Hemmer & Y. Lamadrid-Rose. 1986. Laboratory spawning cues in Menidia beryllina and M. peninsulae (Pisces: Atherinidae) with notes on survival and growth of larvae at different salinities. Environmental Biology of Fishes 15 (2): 107-117. [ Links ]
Miller, R. R., W. L. Minckley & S. M. Norris. 2005. Freshwater fishes of México. The University of Chicago Press. Chicago, 490 p. [ Links ]
Montgomery, D. C. 1991. Design and analysis of experiments. John Wiley & Sons. New York. 589 p. [ Links ]
Nardez, S. R., G. Phonlor & L. Α. Sampaio. 1997. Shyncronization of the hatching time of the silverside Odontesthes bonariensis (Teleostei: Atherinopsidae) through temperature, salinity, and oxygen stimuli. Arquivos de Biología e Tecnología 40 (3): 688-692. [ Links ]
Nikolsky, G. v. 1963. The ecology of fishes. London & NY: Academic Press, 352 p. [ Links ]
N0M-059-SEMARNAT-2010. 2010. Diario Oficial de la Federación. SEGOB. México.
Ojanguren, Α. F. & F. Braña. 2003. Thermal dependence of embryonic growth and development in Brown trout. Journal of Fish Biology 62: 580-590. [ Links ]
Otto, R. G. 1971. Effects of salinity on the survival and growth of pre-smolt coho salmon (Oncorhynchus kisutch). Journal of Fisheries Research Board of Canada 28: 343-349. [ Links ]
Pavlov, D. Α. & E. Moksness. 1997. Development of the axial skeleton in wolfish Anarhichas lupus (Pisces, Anarhichadidae), at different temperatures. Environmental Biology of Fishes 49: 401-416. [ Links ]
Pepin, P. 1991. The effect of temperature and size on development and mortality rates of the pelagic early life history stages of marine fish. Canadian Journal of Fisheries and Aquatic Sciences 48: 503-518. [ Links ]
Pepin, P., D. C. Orr & J. T. Anderson. 1997. Time to hatch and larval size in relation to temperature and egg size in Atlantic cod (Gadus morhua). Canadian Journal of Fisheries and Aquatic Sciences 54 (Suppl. 1): 2-10. [ Links ]
Peterson, R. Η., D. J. Martin-Robichaud & Α. Berge. 1996. Influence of temperature and salinity on length and yolk utilization of striped sea bass larvae. Aquaculture International4: 89-103. [ Links ]
Phonlor, G. & L. Α. Sampaio. 1992. Effect of salinity on growth and survival of Odontesthes argentinensis larvae. Arquivos de Biología e Tecnología 35 (1): 153-155. [ Links ]
Rideout, R. M., E. Α. Trippel & M. K. Litvak. 2005. Effects of egg size, food supply and spawning time on early life history success of haddock Melanogrammus aeglefinus. Marine Ecology Progress Series 285: 169-180. [ Links ]
Sampaio, L. Α. & G. Phonlor. 1996. Efectos de la salinidad en huevos y larvas vitelinas de Odontesthes humensis (Teleostei: Atherinidae). In: Silva, A. & G. Merino (Eds.). Acuicultura en Latinoamerica. IX Congreso Latinoamericano. 2° Simposio Avances y perspectivas de la acuacultura en Chile. Univ. Católica del Norte. Asociación Latinoamericana de Acuicultura. Coquimbo, Chile. pp. 346-349. [ Links ]
Sfakianakis, D. G., G. Koumoundouros, P. Divanach & M. Kentouri. 2004. Os-teological development of the vertebral column and of the fins in Pagellus enythrinus (L. 1758). Temperature effect on the developmental plasticity and morpho-anatomical abnormalities. Aquaculture 232: 407-424. [ Links ]
Sokal, R. R. & F. J. Rohlf. 1969. Biometry: The principles and practice of statistics in biological research. W. H. Freeman & Co. San Francisco, CA. 778 p. [ Links ]
Soto-Galera, E., E. Díaz-Pardo, E. López-López & J. Lyons. 1998. Fish as indicator of environmental quality in the Río Lerma Basin, México. Journal Aquatic Ecosystem Health Manage 1: 267-276. [ Links ]
Soto-Galera, E., J. Paulo-Maya & J. Lyons. 2008. Threatened fishes of the world: Chirostoma riojai (Solorzano and López, 1965) (Atherinopsidae). Environmental Biology of Fishes 83: 343-344. [ Links ]
Tucker, J. W., Jr. 1998. Marine fish culture. Kluwer Academic Publishers. Boston. 750 pp. [ Links ]
Wang, J. C. S. 1974. Atherinidae-silversides. In: Lippson, A. J. & L. Moran (Eds.). Manual for identification of early development stages of fishes of the Potamac River Estuary. Environmental Technology Center. Baltimore, Maryland, pp. 143-151. [ Links ]
Ware, D. M. 1975. Relation between egg size growth, and natural mortality of larval fish. Journal Fisheries Resource 32: 2503-2512. [ Links ]
Watanabe W. O., C. M. Kuo & M. C. Huang. 1984. Experimental rearing of Nile tilapia fry (Oreochromus niloticus) for freshwater culture. ICLARM Technical Report 14. 28 p. [ Links ]
Wootton R. J. 1990. Ecology of teleost fishes. Chapman & Hall, London. 404 p. [ Links ]

