










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.23 no.2 Ciudad de México Mai./Ago. 2013
Listado actualizado de las medusas de la Laguna de Términos, Campeche, México
An updated list of the jellyfish from Laguna de Terminos, Campeche, Mexico
Ricardo Cortés-Lacomba, Carlos Álvarez-Silva y Francisco Gutiérrez-Mendieta
Departamento de Hidrobiología. Universidad Autónoma Metropolitana Unidad Iztapalapa. Av. San Rafael Atlixco No. 186 Col. Vicentina. Delegación Iztapalapa, México, D.F. 09340. México. e-mail: rikardo.hidro@gmail.com.
Recibido: 13 de septiembre de 2012.
Aceptado: 20 de enero de 2013.
RESUMEN
Se analizó la composición y abundancia de las medusas presentes en 42 muestras de zooplancton de la Laguna de Términos, Campeche que fueron recolectadas durante marzo y octubre de 2009 y noviembre de 2010. Se identificaron siete especies, seis de ellas son nuevos registros geográficos para este sistema costero y una (Phialopsis diegensis) es también un nuevo registro para el Atlántico mexicano. Considerando todo el periodo estudiado, la especie más abundante fue Helgicirrha schulzeicon el 52.24% del total de medusas examinadas, seguida por Bougainvillia muscus con el 18.04%. La presencia de P. diegensis y otras especies no registradas anteriormente en la zona, pueden ser resultado de la influencia oceánica en este sistema o bien son posibles especies invasoras derivadas del impacto de las actividades humanas y el tráfico marítimo, actividades que afectan principalmente a las comunidades del zooplancton en las lagunas costeras.
Palabras clave: Abundancia, composición, Laguna de Términos, medusas.
ABSTRACT
The composition and abundance of the jellyfish present in 42 zooplankton samples obtained from the Laguna de Terminos, Campeche, during March and October 2009 and November 2010, were analyzed. Seven species were identified, six of them are new geographic records for the zone, and one (Phialopsis diegensis) is also a new record for the Mexican Atlantic. Considering the entire study period, the most abundant species was Helgicirrha schulzei with 52.24% followed by Bougainvillia muscus with 18.04%. The presence of P. diegensis and other previously unrecorded species in the area could be the result of the oceanic influence in the system or are invasive non-indigenous species derived from the impact of human activities and ship traffic, both activities mainly affecting the zooplankton communities of coastal lagoons.
Key words: Abundance, composition, Jellyfish, Laguna de Terminos.
INTRODUCCIÓN
Las medusas son importantes depredadores planctónicos que se alimentan de copépodos, larvas de invertebrados y larvas de peces (Purcell, 1985; 1997). Debido a su naturaleza depredadora y competencia por el alimento, los cambios en su distribución y abundancia estacional pueden tener consecuencias económicas en las pesquerías locales, ya que pueden competir por alimento con especies comerciales y/o causar impactos negativos en las actividades de crianza de peces. La presencia de metabolitos tóxicos en las medusas las ha hecho importantes en la investigación farmacológica (Barnes & Horridge, 1965). Igualmente, algunas especies de Rhizostomeae, como Stomolophus meleagris (Agassiz, 1862), son importadas en grandes cantidades en Asia donde, junto con otras especies, se utilizan como alimento (Omori & Nakano, 2001). La creciente frecuencia de las agregaciones masivas de medusas en diversas regiones del planeta (Purcell, 2005) es otro aspecto que aumenta la relevancia de los estudios enfocados en este grupo del zooplancton gelatinoso.
La Laguna de Términos, Campeche es una de las lagunas costeras más extensas y estudiadas de México. El conocimiento de la composición de su zooplancton se inició con recolecciones efectuadas en la década de los 1960's (Gómez-Aguirre, 1986). Las medusas de esas muestras fueron analizadas por Canudas-González (1979), quien reportó la presencia de tres especies de la clase Hydrozoa: Eirene pyramidalis (Agassiz, 1862), Octophialucium aphrodite (Bigelow, 1919) y Liriope tetraphylla (Chamisso & Eysenhardt, 1821), y una de la clase Scyphozoa: Aurelia aurita (Linnaeus, 1758). Después de este primer estudio de plancton, el sistema costero ha sido muestreado con diversos objetivos como el análisis de las postlarvas de peneidos, zooplancton, macroplanton y medusas (Suárez-Caabro & Gómez-Aguirre, 1965; Signoret, 1974; Gómez-Aguirre, 1974; Flores Coto & Álvarez Cadena, 1980; Gómez-Aguirre, 1986). Se destaca en estos trabajos el hallazgo de adultos de Chiropsalmus quadrumanus (Müller, 1859), representante del orden Cubomedusae en la Laguna de Términos (Gómez-Aguirre, 1986), ya que este grupo no había sido registrado en las lagunas costeras de México. En general, no se tiene un conocimiento actualizado sobre la composición y abundancia de las medusas en la laguna, ni se han registrado cambios en su composición faunística. La presente investigación tuvo como objetivo principal elaborar un inventario actualizado de la composición y abundancia de las especies de medusas presentes en tres temporadas de muestreo en la Laguna de Términos, Campeche, a partir de muestreos recientes, y hacer algunas consideraciones sobre la biología y distribución de estos taxa.
MATERIALES Y MÉTODOS
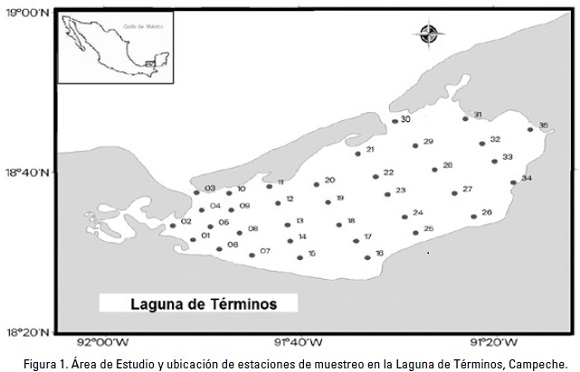
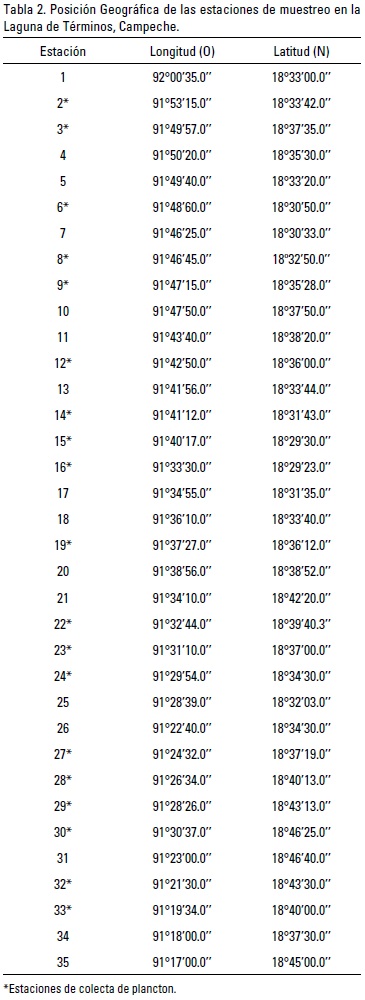
El material analizado se obtuvo durante el desarrollo del proyecto JEST (Joint Environmental Study of Terminos Lagoon) efectuado entre el IRD (L'Institut de Recherche pour le Développement) y la Universidad Autónoma Metropolitana Unidad Iztapalapa. Los especímenes analizados se obtuvieron a partir del análisis de muestras de zooplancton recolectadas con base en una red de 35 estaciones establecidas en la laguna (Fig. 1); aunque no en todas ellas se encontraron medusas (Tabla 2. *Estaciones de colecta de medusas). En el área de estudio (18°52' y 19°00' N y 91°15' y 92°00' W), los muestreos se efectuaron en marzo y octubre de 2009, así como en noviembre de 2010 (Tabla 2). Se utilizó una red estándar con 30 cm de diámetro de boca, largo de 90 cm y luz de malla de 250 um. La red fue arrastrada durante 5 minutos a una velocidad entre 1 y 1.5 nudos, y aproximadamente a 50 cm de la superficie. Las muestras se fijaron con formol al 4%, neutralizado con borato de sodio, y posteriormente fueron selladas y trasladadas al Departamento de Hidrobiología de la Universidad Autónoma Metropolitana Unidad Iztapalapa para su análisis. Adicionalmente, se midió in situ la temperatura y la salinidad del agua mediante una sonda CTD Seabird SB19.

En el laboratorio se procedió a la separación e identificación de las medusas utilizando microscopios ópticos y de disección. La densidad se estandarizó a org/1000 m3. Se recurrió a literatura especializada para la identificación: Ramírez y Zamponi (1981), Bouillon y Boero (2000), Kramp (1959, 1961), Mayer (1910), Schuchert (2007) y Segura-Puertas et al. (2003). Se utilizo para la toma de fotografías una cámara SONY Digital Mavica complementándose con un adaptador universal para microscopios ópticos y de disección. Las medusas fueron depositadas en la colección de zooplancton del Departamento de Hidrobiología de la UAM-Iztapalapa (UAMMLT).
RESULTADOS
Parámetros Ambientales: Durante marzo la temperatura osciló entre los 25 y los 28.3 °C, el promedio fue de 26.3 °C; en octubre fluctuó entre 26.9 y 29.5 °C, el promedio fue de 28.2 °C y en noviembre los valores se encontraron entre 25.9 y 27.4 °C, con un promedio de 26.7 °C. Durante marzo la salinidad osciló entre 8.01 y 38.5 ups, el promedio para este mes fue de 31.58 ups; en octubre fluctuó entre 0.59 y 37.71 ups con un promedio de 32.3 ups, y en noviembre se encontraron valores entre 0.59 y 33.9 ups, con un promedio de 16.6 ups.
Sistemática: Se determinaron siete especies que fueron clasificadas siguiendo el arreglo taxonómico de Bouillon y Boero (2000) y Segura-Puertas etal. (2003) (Tabla 1).
A continuación se presenta algunos de los datos sobre el sitio de colecta y su distribución.
Subclase Anthomedusae Haeckel, 1879
Familia Bougainvillidae Lütken, 1850
Bougainvillia muscus (Allman, 1863) (Figs. 2a, 3a).
Caracteres diagnósticos: Es el primer registro geográfico de esta especie en la Laguna de Términos. Su característica distintiva es la presencia de tentáculos orales divididos 1-2 (raramente 3 veces; Fig. 3a).
Estaciones de colecta: (mes, estación y número de ejemplares): marzo 2(1), 3(15), 6(2), 8(4), 9(11), 19(4), 22(27), 24(9), 27(10), 28(31), 32(11), 33(4).
Parámetros ambientales: 25.14-35.63 ups y 25-27 °C.
Abundancia: 58-925 org • 1000 m-3 (promedio: 100.28 org • 1000 m-3).
Distribución en México: Laguna Madre, Tamaulipas (Mendoza-Becerril, 2006) y Laguna de Términos, Campeche, México (presente estudio).
Distribución Mundial: Océano Atlántico del Norte incluyendo Mar Mediterráneo (Van der Land et al., 2001), Atlántico Sur (Bouillon, 1999), Océano Ártico e Indo-Pacífico (López et al., 2007), Oeste de África (Segura-Puertas et al., 2009).
Thamnostoma tetrellum (Haeckel, 1879) (Figs. 2b, 3b).
Caracteres diagnósticos: Esta especie se caracteriza por la presencia de un manubrio cúbico y tres tentáculos orales insertados al final de la boca (Fig. 3b).
Estaciones de colecta: marzo 9(1), 19(2), 22(15), 27(5), 28(20), 32(8).
Parámetros ambientales: 25.14-35.63 ups y 25.7-27 °C.
Abundancia: 58-596 org • 1000 m-3 (promedio: 43.38 org • 1000 m-3).
Distribución en México: Mar Caribe (Segura-Puertas, 1992), Laguna de Términos, Campeche (presente estudio). Es el primer registro geográfico de esta especie en la Laguna de Términos.
Distribución Mundial: Caribe y Sur del Océano Atlántico (Segura-Puertas, 1991); Brasil (Segura-Puertas et al., 2009).
Subclase Leptomedusae Haeckel, 1879
Familia Campanulariidae Johnston, 1836
Clytia globosa (Mayer, 1900) (Figs. 2c, 3c, d).
Caracteres diagnósticos: Taxonómicamente, C. globosa se caracteriza por la presencia de gónadas ovales y vesículas marginales cerradas (Figs. 3 c-d).
Estaciones de colecta: marzo 9(2), 28(3), 32(2); noviembre 8(61).
Parámetros ambientales: 23.13-35.63 ups y 25.7-27 °C.
Abundancia: 2.71-89 org • 1000 m-3 (promedio: 80.23 org • 1000 m-3).
Distribución en México: Laguna Madre de Tamaulipas (Mendoza-Becerril et al., 2009) y Laguna de Términos, Campeche (presente estudio); es un nuevo registro geográfico para esta laguna.
Distribución Mundial: Común en zonas tropicales, Florida, la India, Océano Atlántico Sur (Segura-Puertas et al., 2009); considerada cosmopolita (Mendoza-Becerril et al., 2009).
Familia Eirenidae Haeckel, 1879
Eutima mira (McCrady, 1857) (Figs. 2d, 3e).
Caracteres diagnósticos: E. mira se puede distinguir de sus congéneres conocidos por la presencia de cirros laterales y bulbos en el margen umbrelar (Fig. 3e).
Estaciones de colecta: marzo 19(4), 22(3), 27(3), 28(1); octubre 22(1), 23(7), 24(1), 30(1).
Parámetros ambientales: 25.14-36.95 ups y 25.0-28.25 °C.
Abundancia: 29-534 org • 1000 m-3 (promedio: 23.68 org • 1000 m-3).
Distribución en México: Bahía de La Ascensión, Quintana Roo (Suárez-Morales et al., 1997), sur del Golfo de México en la plataforma Continental de Yucatán (Segura-Puertas, 1992) y Laguna de Términos, Campeche (presente estudio). Este es el primer registro de la especie en la Laguna de Términos.
Distribución Mundial: Aguas europeas del Atlántico Oriental (Van der Land et al., 2001); Atlántico Occidental, desde Nueva Inglaterra a Brasil (Segura-Puertas et al., 2009).
Helgicirrha schulzei Hartlaub, 1909 (Figs. 2e, 3f).
Caracteres diagnósticos: Su característica distintiva es la presencia de cirros laterales y bulbos rudimentarios en el margen umbrelar (Fig. 3f).
Estaciones de colecta: marzo 12(12), 19(5), 22(6), 24(7), 28(12); octubre 22(1), 23(81), 24(50), 27(5), 32(11).
Parámetros ambientales: 25.14-36.95 ups y 25.7-28.25 °C.
Abundancia: entre 67-6180 org • 1000 m-3 (promedio: 290.44 org • 1000 m-3).
Distribución en México: Mar Caribe (Segura-Puertas et al., 2003), Bahía de la Ascensión, Quintana Roo (Suárez-Morales et al., 1997; Canché-Canché & Castellanos-Osorio, 2005) y Laguna de Términos, Campeche (presente estudio). Es un nuevo registro geográfico para esta laguna.
Distribución Mundial: Tropical, subtropical. Sur del Océano Atlántico (Ramírez & Zamponi, 1981) aguas europeas del Atlántico Nororiental (Van der Land et al., 2001).
Phialopsis diegensis (Torrey, 1909) (Fig. 2f).
Caracteres diagnósticos: Phialopsis diegensis se caracteriza por la presencia de cirros marginales dispersos, bulbos rudimentarios y vesículas marginales.
Estaciones de colecta: marzo 9(1) ,19(4), 24(7), 28(2).
Parámetros ambientales: 25.14-35.63 ups y 25.7-27 °C.
Abundancia: 59.6-304 org • 1000 m-3 (promedio: 16.75 org • 1000 m-3).
Distribución en México: Laguna de Términos, Campeche, México (presente estudio). Es un nuevo registro para la Laguna de Términos.
Distribución Mundial: Tropical, Atlántico Norte, Atlántico Sudoccidental, Costa este de África, en el Pacífico que se registra desde Sur oeste de las Islas Galápagos hasta San Diego en California (Kramp, 1959; Ramírez & Zamponi, 1981).
Subclase Trachymedusae Haeckel, 1866 (1879)
Familia Geryoniidae Eschscholtz, 1829
Liriope tetraphylla (Chamisso & Eysenhardt, 1821) (Fig. 2g).
Caracteres diagnósticos: Esta especie se caracteriza por una boca con 4 labios pequeños. Las gónadas se localizan sobre los 4 conductos radiales y observan una gran variedad de formas y tamaños, desde oval, circular y en forma de corazón. Los estatocistos son 8 y se localizan cerca de la base de los tentáculos.
Estaciones de colecta: noviembre 19 (1).
Parámetros ambientales: 23.13-30.22 ups y 25.8-26.8 °C.
Abundancia: 44 org • 1000 m-3 (promedio: 1.25 org • 1000 m-3).
Distribución en México: Banco de Campeche, Laguna de Términos, Campeche y Pacifico Mexicano (Segura-Puertas et al., 2010).
Distribución Mundial: Pacífico Tropical, Atlántico Norte y Sur (Kramp, 1959), tropical.
DISCUSIÓN
Segura-Puertas et al. (2003) mencionan que existen seis especies del género Bougainvillia reportadas para el Golfo de México, aunque en un trabajo posterior Segura-Puertas et al. (2009) reconocen sólo cinco. Mendoza-Becerril (2006) reporta a Bougainvillia muscus como una especie de ambientes euhalinos en la porción norte del Golfo de México, particularmente en la Laguna Madre, en Tamaulipas. Loman-Ramos et al. (2007), señalaron la presencia de esta especie en la plataforma continental de los estados de Veracruz y Tabasco, donde su abundancia mostró valores altos y se le consideró como una especie circumtropical. Tales autores la consideran, como una especie nerítica y de aguas estuarinas.
Estos datos concuerdan con los encontrados en la presente investigación, ya que se encontró una elevada frecuencia de esta especie en las estaciones de muestreo (18.03% de abundancia relativa).
Thamnostoma tetrellum es la única especie del género reportada para el Golfo de México y el Mar Caribe, se considera como una especie propia de aguas neríticas (Segura-Puertas, 1991).
Clytia globosa se considera una especie hiperhalina, que ocurre en ambientes neríticos y estuarinos, con notable tolerancia a cambios en la salinidad (Mendoza-Becerril et al., 2009; Segura-Puertas et al., 2009). 6 especies del género Clytia son reconocidas en el Golfo de México (Segura-Puertas et al., 2009).
Eutima mira es una especie que se distribuye en ambientes costeros con salinidad variable; en México se ha registrado en la Bahía Ascensión, Quintana Roo Mar Caribe (Suárez-Morales et al., 1997) y en el sur del Golfo de México en la plataforma Continental de Yucatán (Segura-Puertas, 1992). Este también es el primer registro de la especie en la Laguna de Términos. Aunque existen más de 20 especies de este género, sólo tres de ellas han sido observadas en aguas del Golfo de México (Segura-Puertas et al., 2009).
Helgicirrha schulzei tiene una amplia distribución y es la especie más abundante en la Bahía de La Ascensión, Quintana Roo, donde representa entre el 35 y el 90% de la abundancia de las medusas en ese sistema costero (Suárez-Morales et al., 1997; Canché-Canché & Castellanos-Osorio, 2005).
Phialopsis diegensis es también un nuevo registro para la Laguna de Términos. La especie fue descrita originalmente a partir de material recolectado en San Diego, California (Torrey, 1909), posteriormente se citó para el Pacífico Tropical Oriental (Vanhbffen, 1912) y para el suroeste de las Islas Galápagos (Bigelow, 1919), como P. diegensis. Su distribución en el Pacífico Oriental se extiende hasta la región meridional, frente a las costas chilenas (Fagetti, 1973), se trata de una especie con una amplia distribución nerítica. Como es sabido, existen especies del zooplancton que habitan la región del Pacífico Tropical Oriental que también se encuentran en las aguas tropicales del Atlántico, posiblemente debido a la conexión directa de ambos océanos a través del actual canal o del Istmo de Panamá, en un período geológico anterior (Kramp, 1959, 1968).
Liriope tetraphylla ya había sido reportada anteriormente para esta laguna; Canudas-González (1979) encontró un ejemplar en enero de 1985, al igual que sucedió durante esta investigación, durante la que también se encontró un solo espécimen en noviembre de 2010. Ramírez y Zamponi (1981) la consideran cosmopolita y la han asociado con aguas oceánicas templadas y tropicales. Loman-Ramos et al. (2007) reportan a esta especie como la más abundante durante 1999 en el Banco de Campeche. Su presencia en la laguna de Términos se podría atribuir a su transportación pasivamente por corrientes marinas hacia el interior de la laguna por la boca de Paso Real al NE, mediante el flujo que pasa por Punta Matamoros y se dirige hacia la Boca del Carmen en la parte NW, donde Gómez-Aguirre (1986) registró zonas de salinidad mayor a 27.5 ups.
Abundancia y riqueza de especies: La abundancia promedio de las medusas encontradas (Tabla 3) osciló entre 32.79 (noviembre) y 115.12 (octubre) org/1000 m3. En marzo B. muscus fue la especie más abundante con 300.86 org/1000 m3, seguida por T. tetrellum con 130.14 org/1000 m3; ambas especies representaron el 68.2% del total de medusas. Durante este mes se presentó la mayor diversidad de especies. En octubre solamente se presentaron dos especies pero la abundancia total de medusas fue mayor que en los otros meses, E. mira y H. schulzei tuvieron una abundancia promedio de 23.68 y 290.44 org/1000 m3, respectivamente. En noviembre también se presentaron solo dos especies y la abundancia promedio fue la más baja de todo el periodo estudiado, C. globosa y L. tetraphylla tuvieron una abundancia promedio de 1.25 y 80.23 org/1000 m3, respectivamente. De las especies encontradas con anterioridad en la Laguna de Términos por Canudas-González (1979), solo L. tetraphylla fue encontrada en las muestras estudiadas en la presente investigación. Las 6 especies restantes encontradas representan nuevos registros geográficos para la Laguna de Términos e incluso Phialopsis diegensis constituye un nuevo registro para el Atlántico Mexicano.
Ruiz et al. (1997) consideran que las especies de medusas no nativas son cada vez más visibles en los hábitats marinos y estuarínos en todo el mundo; la mayoría de estas invasiones se debe a las actividades humanas y al tráfico marítimo global. Aunque la importancia relativa de los mecanismos de dispersión de las diferentes especies varía espacial y temporalmente, el movimiento mundial del agua de lastre de los buques parece ser el vector más grande para la transferencia de especies invasoras. Es muy común que las medusas invadan estuarios en todo el mundo. Genzano et al. (2006) relacionan la presencia de la hidromedusa Blackfordia virginica (Mayer, 1910), como una especie con características invasoras. Su distribución natural está asociada a aguas de Europa en el Mar Negro y el Mar Caspio y desde hace menos de 40 años se estableció en Brasil, donde fue registrada particularmente en Pernambuco, estuario Itamaracá y cuencas cercanas. También se ha registrado en el Golfo de México y en las costas del Pacífico (Mills & Sommer, 1995; Álvarez-Silva, 2003). Así, los estuarios han sido sitios comunes de invasiones pelágicos, acumulando desde decenas hasta cientos de especies no nativas por estuario que incluyen a la mayoría de los grupos taxonómicos y tróficos importantes en el zooplancton. Los resultados de la presente investigación parecen confirmar que esos procesos han afectado y modificado la composición de la medusofauna de sistemas costeros del Golfo de México, incluyendo la Laguna de Términos.
AGRADECIMIENTOS
Se agradecen los comentarios y la información proporcionada por el Dr. Peter Schuchert del Museo de Historia Natural de Ginebra, Suiza, que ayudaron a la determinación específica de las medusas. Al Dr. Eduardo Suárez Morales del Departamento de Ecología y Sistemática Acuática de El Colegio de la Frontera Sur (ECOSUR-CHETUMAL), por sus comentarios al manuscrito y al Dr. Gabriel Genzano del Departamento de Ciencias Marinas, Facultad de Ciencias Exactas y Naturales, Universidad Nacional de Mar del Plata, Argentina por sus observaciones y revisión final del manuscrito. El proyecto JEST estuvo financiado por l'Institut de Recherche pour le Développement.
REFERENCIAS
Álvarez-Silva, C., S. Gómez-Aguirre & M. G. Miranda-Arce. 2003. Variaciones morfológicas en Blackfordia virginica (Hydroidomedusae: Blackfordiidae) en lagunas costeras de Chiapas, México. Revista de Biología Tropical 51: 409-412. [ Links ]
Barnes, W. P. & G. A. Horridge. 1965. A neuropharmacological active substance from jellyfish ganglia. The Journal of Experimental Biology 42: 257-267. [ Links ]
Bigelow, H. B. 1919. Hydromedusae, Siphonophores and ctenophores of the "Albatross" Philippine Expedition. Contribution to the Biology of the Philippine Archipelago and adjacent regions 1: 279-362. [ Links ]
Bouillon, J. 1999. Hydromedusae. In Boltovskoy, D. (Ed.). South Atlantic zooplankton, Vol. 1. Backhuys Publishers, Leiden, the Netherlands, pp. 385-465. [ Links ]
Bouillon, J. & F. Boero. 2000. Phylogeny and Classification of Hydroidomedusae. The Hydrozoa: a new classification in the light of old knowledge. Thalassia Salentina 24: 1-46. [ Links ]
Canudas-González, A. 1979. Contribución al conocimiento de las medusas (Coelenterata) de la Laguna de Términos, Campeche, México. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 6 (1): 183-188. [ Links ]
Canché-Canché, V. E. & I. Castellanos-Osorio. 2005. Medusas (Cnidaria) de la Bahía de la Ascensión, Quintana Roo, México (1997). Hidrobiológica 15 (1): 65-72. [ Links ]
Fagetti, E. 1973. Medusas de aguas Chilenas. Revista de Biología Marina Valaparaíso 15 (1): 31-75. [ Links ]
Flores-Coto, C. & J. Álvarez-Cadena. 1980. Estudios preliminares sobre abundancia y distribución del ictioplancton en la Laguna de Términos, Campeche. Anales Centro Ciencias del Mar y Limnología. Universidad Nacional Autónoma de México 2: 67-78. [ Links ]
Genzano, G., H. Mianzan, E. M. Acha & E. Gaitán. 2006. First record of the invasive medusa Blackfordia virginica (Hydrozoa: Leptomedusae) in the Río de la Plata estuary, Argentina-Uruguay. Revista Chilena de Historia Natural 79: 257-261. [ Links ]
Gómez-Aguirre, S. 1974. Reconocimientos estacionales de Hidrología y Plancton en la Laguna de Términos, Campeche México (1964-1965). Anales Centro Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 1: 61-82. [ Links ]
Gómez-Aguirre, S. 1986. Ocurrencia de Chiropsalmus quadrumanus (Scyphozoa: Cubomedusae) en la laguna de Términos. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 57, Serie Zoología 2: 227-234. [ Links ]
Kramp, P. L. 1959. The Hydromedusae of the Atlantic Ocean and adjacent Waters. Dana Reports 46: 1-283. [ Links ]
Kramp, P. L. 1961. Synopsis of the Medusae of the world. Journal of the Marine Biological Association ofthe United Kingdom 40: 81-198. [ Links ]
Kramp, P. L. 1968. The Hydromedusae of the Pacific and Indian Oceans. Sections II and III. Dana Reports72: 1-200. [ Links ]
Loman-Ramos, L., U. Ordóñez-López & L. Segura-Puertas. 2007. Variación espacial de la comunidad de medusas (Cnidaria) del sur del Golfo de México, durante el otoño de 1999. Hidrobiológica 17 (1): 203-212. [ Links ]
López, R., J. Medellín, D. Castañeda & 0. Martínez. 2007. Programa: dinámica ecológica del zooplancton en el Océano Pacífico colombiano. Cruceros 2007-2010 de la serie ERFEN (Estudio Regional del Fenómeno El Niño). Informe de resultados crucero ERFEN, Universidad Militar Nueva Granada, Bogotá, pp. 43-61. [ Links ]
Mayer, A. G. 1910. Medusae ofthe World volume I. The Hydromedusae. Carnegie Institution of Washington, pp. 155-272. [ Links ]
Mills, C. E. & F. Sommer. 1995. Invertebrate introductions in marine habitats: Two species of hydromedusae (Cnidaria) native to the Black Sea, Maeotias inexspectata and Blackfordia virginica, invade San Francisco Bay. Marine Biology 122: 279-288. [ Links ]
Mendoza-Becerril, M. A. 2006. Diversidad de Medusas (Phylum: Cnidaria) en la Laguna Madre, Tamaulipas. México. Tesis Profesional. Escuela Nacional de Ciencias Biológicas. Instituto Politécnico Nacional, 60 p. [ Links ]
Mendoza-Becerril, M. A., A. Ocaña-Luna, M. Sánchez-Ramírez & L. Segura-Puertas. 2009. Primer registro de Phialella quadratay ampliación del límite de distribución de ocho especies de hidromedusas (Hydrozoa) en el Océano Atlántico Occidental. Hidrobiológica 19 (3): 257-267. [ Links ]
Omori, M. & E. Nakano. 2001. Jellyfish fisheries in Southeast Asia. Hydrobiologia 451: 19-26. [ Links ]
Purcell, J. E. 1985. Predation on fish eggs and larvae by pelagic cnida-rians and ctenophores. Bulletin of Marine Science 37: 739-755. [ Links ]
Purcell, J. E. 1997. Pelagic cnidarians and ctenophores as predators: selective predation, feeding rates and effects on prey populations. Annales de l'Institut Oceanographique 73: 125-137. [ Links ]
Purcell, J. E. 2005. Climate effects on formation of jellyfish and ctenophore blooms: a review. Journal of the Marine Biological Association of U.K. 85: 461-476. [ Links ]
Ramírez, F. C. & M. 0. Zamponi. 1981. Hydromedusae. In: Boltovskoy, D. (Ed.). Atlas del Zooplancton del Atlántico Sudoccidental y métodos de trabajo con el zooplancton marino. Publicación Especial del Instituto Nacional de Investigación y Desarrollo Pesquero, Mar de Plata, Argentina, pp. 443-469. [ Links ]
Ruiz, G. M., J. T. Carlton, E. D. Grosholz & A. H. Hines. 1997. Global Invasions of Marine and Estuarine Habitats by Non-Idigenous Species: Mechanisms, Extent, and Consequences. American Zoologist 37: 621-632. [ Links ]
Schuchert, P. 2007. The European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Filifera Part 2. Revue Suisse de Zoologie 114 (2): 195-396. [ Links ]
Segura-Puertas, L. 1991. New records of two species of Hydromedusae (Cnidaria) from the Mexican Caribbean. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 18 (1): 132-135. [ Links ]
Segura-Puertas, L. 1992. Medusae (Cnidaria) from the Yucatan Shelf and Mexican Caribbean. Bulletin of Marine Science 51: 353-359. [ Links ]
Segura-Puertas, L., E. Suárez-Morales & L. Celis. 2003. A checklist of the Medusae (Hydrozoa, Scyphozoa and Cubozoa) of Mexico. Zootaxa 194: 1-15. [ Links ]
Segura-Puertas, L., L. Celis & L. Chiaverano. 2009. Medusozoans (Cnidaria: Cubozoa, Scyphozoa and Hydrozoa) of The Gulf of Mexico. In: Felder, D. L. & D. R. Camp (Eds.). Gulf of Mexico, Origin, Waters, and Biota, Vol. 1, Biodiversity, Chapter 18. Texas A&M University Press, U.S.A, pp. 269-386. [ Links ]
Segura-Puertas, L., C. Franco-Gordo, E. Suárez-Morales, R. Gasca & E. Godínez-Domínguez. 2010. Composición y distribución de las medusas (Cnidaria: Medusozoa) en la plataforma continental central del Pacífico mexicano. Revista Mexicana de Biodiversidad81 (1): 103-112. [ Links ]
Signoret, M. 1974. Abundancia, tamaño y distribución de camarones (Crustacea Penaideae) de la Laguna de Términos, Campeche, México, y su relación con algunos factores hidrológicos. Anales del Instituto de Ciencias del Mar y Limnología, Universidad Nacional Autónoma de México 45. Serie Zoología 1: 119-139. [ Links ]
Suárez-Caabro, J. A. & S. Gómez-Aguirre. 1965. Observaciones sobre el plancton de la Laguna de Términos, Campeche, México. Bulletin of Marine Science 15 (4): 1072-1120. [ Links ]
Suárez-Morales, E., M. Zamponi & R. Gasca. 1997. Hydromedusae (Cnidaria: Hydrozoa) of Bahía de la Ascensión, Caribbean coast of México: a seasonal survey. Proceedings of the 6th International Conference on Coelenterate Biology. National Natuurhistorisch Museum. Leiden. pp. 465-472. [ Links ]
Torrey, H. B. 1909. The Leptomedusae of the San Diego region. University of California Publications in Zoology 6: 11-31. [ Links ]
Van der Land, J., W. Vervoort, S. D. Cairns & P. Schuchert. 2001. Hydrozoa. In: Costello, M. J., C. Emblow & R. J. White (Ed.). European register of marine species: a check-list of the marine species in Europe and a bibliography of guides to their identification. Collection Patrimoines Naturelles. Muséum National d'Histoire Naturelle. Paris, 50: 112-120. [ Links ]
Vanhoffen, E. 1912. Die Craspedoten Medusen des "Vettor Pisani". Zoologica Stuttgart 67: 1-34. [ Links ]

