










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.23 no.1 Ciudad de México ene./abr. 2013
Artículos
Variación de la captura del recurso camarón, en relación con la disposición geográfica de los sedimentos en la zona 90 de pesca, Golfo de Tehuantepec, Oaxaca, México
Variation of shrimp total catch related to the geographic distribution of sediments at fishing zone 90, Gulf of Tehuantepec, Oaxaca, Mexico
Pedro Cervantes-Hernández y Aideé Egremy-Valdez
Universidad del Mar, campus Puerto Ángel Instituto de Recursos. Carretera a Zipolite, Ciudad Universitaria s/n, Distrito de San Pedro Pochutla A. P. 47. Puerto Ángel, Oaxaca, 70902. México e-mail: pch@angel.umar.mx
Recibido: 05 de junio de 2012.
Aceptado: 15 de marzo de 2013.
RESUMEN
La zona 90 de pesca se localiza en el Golfo de Tehuantepec, y está dividida en cinco sub-zonas de la S-91 a S-95. La "Costa Oaxaqueña" se ubica entre la ciudad de Salina Cruz y la Boca de Tonalá (en el sistema lagunar Mar Muerto), donde se localizan las sub-zonas S-91 y S-92. La "Costa Chiapaneca" se ubica entre la Boca de Tonalá y Puerto Chiapas, abarcando las sub-zonas S-93, S-94 y S-95. A partir de la S-91 y hacia la S-95, la captura total del recurso camarón mostró un incremento de 160 a 1,030 Kg, coincidiendo con un gradiente descendente de gravas y un gradiente ascendente de lodos (g). Las arenas estuvieron presentes en todos las sub-zonas, pero éstas no fueron dominantes a nivel costero, excepto hacia el frente oceánico. Con base en técnicas de sistemas de información geográfica, en la zona 90 de pesca se observaron y delimitaron dos sub-sistemas productivos de camarón: i) la "Costa Oaxaqueña", en donde se registró un bajo nivel de captura de camarón, con gravas dominantes en la S-91 y una mezcla de gravas y arenas en la S-92; ii) la "Costa Chiapaneca", en donde se observó un alto nivel de captura de camarón, con lodos dominantes en la S-93, S-94 y S-95. En estas últimas, la captura del recurso camarón evidenció una distribución ecológica agregada (binomial negativa). La correlación entre la captura total del recurso camarón y los lodos resultó significativa (R = 0.867, p < 0.05), por otra parte con respecto a las gravas y arenas ésta no fue significativa.
Palabras claves: Camarón, captura, Chiapas, Golfo de Tehuantepec, sedimentos.
ABSTRACT
The fishing zone 90 is located in the Gulf of Tehuantepec and this is composed of five sub-sectors from S-91 to S-95. The "Oaxaqueña Coast" is located between Salina Cruz City and The Boca de Tonalá (in the Mar Muerto lagoon sytem), where the S-91 and S-92 sub-sectors are located. The " Chiapaneca Coast" is located between the Boca de Tonalá and Puerto Chiapas, where the S-93, S-94 and S-95 sub-sectors are located. From S-91 towards S-95 sub-sectors the total catch of shrimp resource increased from 160 to 1,030 Kg, coincided with a descendent gradient of gravel and an ascendant gradient of mud (g). Sand was present in all sub-sectors, but was not dominant at the coast level except towards the oceanic front. Based on geographic information system techniques, at the fishing zone 90 two shrimp harvest sub-systems were observed and delimited. The first of those is the "Oaxaqueña Coast" where was recorded a lower shrimp catch level with gravel dominant in the S-91 sub-sector and a gravel-sand mix in the S-92 sub-sector. The second the " Chiapaneca Coast" where was recorded a higher shrimp catch level with mud dominant in the S-93, S-94 and S-95 sub-sectors. In this last place, shrimp resource showed an ecological distribution aggregate (Negative Binomial). Estimated correlation between total catch of shrimp resource and mud was significant (R = 0.867, p < 0.05) but it was not significant with gravel and sand.
Key words: Catch, Chiapas, Gulf of Tehuantepec, sediments, shrimp.
INTRODUCCIÓN
El Golfo de Tehuantepec (GT) en la costa sur del Pacífico Tropical mexicano, es una región de gran complejidad ecológica (Gallegos-García & Barberán-Falcón, 1998), e intensa producción pesquera (Ramos-Santiago et al., 2006; Cervantes-Hernández et al., 2010). En el GT se localiza la zona 90 de pesca (subdividida en cinco sub-zonas, de la S-91 a la S-95), delimitada entre Punta Chipehua, próxima a la ciudad de Salina Cruz, Oaxaca, México (16º01'31.39''N y 95º22'24.56'' O) y Puerto Chiapas, Chiapas, México (14º40'55.81'' N y 92º23'44.13''O) (Cervantes-Hernández et al., 2012 b) (Fig. 1). En la zona 90 de pesca, se registra la mayor actividad de pesca marina y ribereña (en los sistemas lagunares), orientada principalmente a la explotación del recurso camarón (Cervantes-Hernández et al., 2012b). Las especies mayormente explotadas, son: "el café" Farfantepenaeus californiensis (Holmes, 1900) y "el blanco" Litopenaeus vannamei (Boone, 1931) (Cervantes-Hernández, 2008).
En 1958 se documentó que los pescadores de camarón del norte de California, E.U.A., desarrollaron un conocimiento empírico, que les permitía relacionar el volumen de captura obtenido, de acuerdo a los arrastres realizados sobre de diferentes tipos de sustrato (Williams, 1958). En el caso de F. californiensis y L. vannamei, trabajos recientes sugieren que los adultos reproductores marinos (Cervantes-Hernández, 2008) y la fracción juvenil recién reclutada provenientes de los sistemas lagunares del GT (Cervantes-Hernández et al., 2012b), evidencian variaciones de abundancia a largo de la zona 90 de pesca, por lo que se advierte preliminarmente, que ésta se encuentra condicionada por el tipo de fondo marino. Gracia (1992), señaló que la disposición geográfica y el tipo de sustrato, son factores relevantes, que deben ser considerados para explicar la distribución ecológica de los camarones de la familia Penaeidae.
La distribución ecológica para diferentes especies de camarón, tales como Farfantepenaeus aztecus (Ives, 1891), Litopenaeus setiferus (Linneaus, 1767) y Farfantepenaeus duorarum (Burkenroad, 1939) y su relación con la disposición geográfica para diferentes tipos de sustrato, inició con los trabajos pioneros de autores como: Burkenroad (1939), Williams (1958) y Grady (1971) en la plataforma marina al norte de California, E.U.A. y el Golfo de México. Este tipo de trabajo, no se ha hecho para el Pacífico Tropical mexicano, sin embargo, con respecto a la profundidad, se han descrito patrones temporales de distribución, para diferentes especies de camarón en las costas de Sinaloa y Nayarit (Ruiz-Luna et al., 2010) y en el Golfo de California (Aragón-Noriega et al., 1999).
Respecto a la disposición geográfica de los sedimentos en el GT, se han realizado trabajos con muestras recolectadas durante mediados de los años ochenta e inicios del dos mil (Morales de la Garza & Carranza-Edwars, 1995; Tapia-García et al., 2007; Machain-Castillo et al., 2008).
A pesar del gran avance en la investigación pesquera para el recurso camarón del GT (Cervantes-Hernández et al., 2006, 2008a, 2008b, 2010, 2012a, 2012b; Ramos-Cruz et al., 2006; Cervantes-Hernández, 2008), aún no se han detectado y delimitado geográficamente, las áreas de mayor abundancia del recurso camarón en la zona 90 de pesca. Esto último, constituye el objetivo principal del presente trabajo, en donde para verificar y explicar los cambios de abundancia del recurso camarón a largo de la zona 90 de pesca, se utilizó la disposición geográfica para diferentes tipos de sedimentos. Lo anterior, es de vital importancia en el establecimiento de zonas estratégicas de conservación, que integradas a un plan de manejo, permitirían recomendar, la reducción del esfuerzo pesquero, mediante un señalamiento precautorio geográficamente dirigido.
MATERIAL Y MÉTODOS
En 31 estaciones georreferidas a lo largo de la zona 90 de pesca (y cuatro fuera de ésta), entre los 10 y 100 m (Fig. 1), el Buque Oceanográfico (B/O) UMAR, propiedad de la Universidad del Mar campus Puerto Ángel, Oaxaca, México, realizó arrastres científicos, durante la temporada de veda agosto 2009, y de carácter comercial, en la temporada de pesca octubre 2010. Las maniobras fueron perpendiculares a la línea de costa con una hora de duración, utilizando redes de monofilamento con una luz de malla de 57.15 mm. Los registros de la captura total (Kg) del recurso camarón de tipo científica y comercial, fueron entregadas al laboratorio de SIG de la UMAR, para su análisis. Un resumen de la captura total del recurso camarón se presenta en la Tabla 1.
Por otra parte, el B/O Altair B1-03, propiedad de la Secretaría de Marina (SEMAR) de la Armada de México, realizó el crucero oceanográfico CHIS-COYVER-0109 en 49 estaciones georreferidas a lo largo del GT, entre los 13 y 201 m de profundidad del 17 de abril al 23 de mayo 2009. De éste crucero, únicamente se seleccionaron 34 estaciones georreferidas, las que estaban ubicadas dentro de la zona 90 de pesca, y que para facilitar su representación en este trabajo fueron reenumeradas (Fig. 1). En cada estación de recolecta, sólo una muestra de sedimentos de aproximadamente 2.5 kg fue recolectada con una draga pre-fabricada del tipo POMAR, con una boca de 30 × 20 cm.
La Estación de Investigación Oceanográfica de Salina Cruz, Oaxaca, México, supervisó el envío de todas las muestras de sedimentos al laboratorio de sedimentología del Centro Regional de Investigación Pesquera de Salina Cruz (CRIP-SC), quien fue el encargado de su procesamiento, el cual consistió en la clasificación del tipo de sustrato, según su diámetro en: gravas (entre 2 y 60 mm), arenas (entre 0.06 y 2 mm) y lodos (entre 0.002 y 0.06 mm) en g. Para lo anterior, el CRIP-SC utilizó tamices de malla cuadrada de la serie ASTM (del No 8 al No 230), siguiendo el procedimiento de clasificación granulométrica propuesta por Folk (1969). El CRIP-SC generó una base de datos con la información antes señalada, la cual fue puesta a disposición del laboratorio de SIG de la UMAR, para su análisis. Un resumen de la información para los diferentes tipos de sustrato se presenta en la Tabla 2.
Correlación múltiple. Para cada sub-zona de pesca (Tabla 1, Tabla 2), se estimó el valor promedio de la captura total del recurso camarón, gravas, arenas, lodos, gravas más arenas y arenas más gravas, así como la composición porcentual de éstas últimas. Con los valores promedio, y mediante un análisis de correlación múltiple, se analizó la variación de la captura total del recurso camarón, con relación a los diferentes tipos de sustrato. Esto último, se realizó con el programa computacional Statistica® versión 7.
Distribución ecológica. Los registros de la captura total del recurso camarón, se reorganizaron en una nueva matriz (matriz columna de orden 35), denominada Ni en la cual cada elemento (Yi), representa la suma de la captura total del recurso camarón de las dos temporadas (yi1 + yi2), para las 35 estaciones de recolecta (Tabla 1).
De acuerdo con Ludwig y Reynolds (1988), Yi se utilizó para estimar la varianza (S2) y la media (Y) de Ni. Y fue integrada a la función de densidad probabilística "Poisson" (Krebs, 1999), para verificar estadísticamente las siguientes hipótesis:
Ho: S2 = Y, El recurso camarón evidenció en la zona 90 de pesca una distribución ecológica aleatoria. Ha: S2 > Y, el recurso camarón evidenció en la zona 90 de pesca una distribución ecológica agregada.
En el caso de que la hipótesis Ha fuese aceptada, la validez de ésta se corroboraría estadísticamente según el procedimiento de Ludwig y Reynolds (1988), integrado a S2 y Y en la función de densidad probabilística "Binomial Negativa".
Para el caso específico de la función de densidad probabilística "Poisson", la confrontación estadística entre Yi y Yi, se realizó utilizando la distribución ji-cuadrada (Xi 2) con n-2 grados de libertad. Y ^i = s ⋅ P(Yi), donde s es la suma total de Yi y P(Yi) es la estimación de la probabilidad de "Poisson" para Yi.
Para el caso específico de la función de densidad probabilística "Binomial Negativa", la confrontación estadística entre Yi y Y i, se realizó utilizando la distribución Xi 2 con 3 grados de libertad. Y ^i = s ⋅ P(Yi), donde s es la suma de Yi y P(Yi) es la estimación de la probabilidad de "Binomial Negativa" para Yi.
El análisis de la distribución ecológica, se realizó con los programas computacionales Microsoft Office Excel® versión 2007 y Statistica® versión 7.
Los registros de la captura total del recurso camarón en yi1 y yi2 (Tabla 1), se transformaron para las 35 estaciones de recolecta, a valores probabilísticos de captura o P(c), dentro un rango estandarizado entre cero y uno. Para realizar la trasformación, se usó la función de densidad probabilística ligada a la hipótesis aceptada. La transformación se realizó con el fin de reducir el efecto de la variación en el registro de la captura total del recurso camarón (Tabla 1), y facilitar con ello, su interpolación geográfica, utilizando a la P(c).
Mediante la creación de un raster de valor-punto (raster temático A), a cada una de las 35 estaciones georreferidas se le asignó el correspondiente valor de P(c), distinguiendo por capas temáticas, a yi1 y yi2. Los diferentes tipos de sustrato inscritos en la Tabla 2, fueron asignadas a las 34 estaciones georreferidas, mediante la creación de un raster de valor-punto (raster temático B), distinguiendo por capas temáticas a las gravas, arenas y lodos. Todo lo anterior, se realizó con el programa computacional Integrated Land and Water Information System® versión 3.3. (ILWIS por sus siglas en ingles).
Análisis Geo-Estadísticos. Interpolación. Para complementar el análisis de correlación por sub-zona de pesca, los espacios vacíos entre estaciones georreferidas, en los raster de punto A y B, fueron interpolados utilizando la técnica de "Promedio Móvil". De acuerdo con Uriel (1995), el promedio móvil aplicado a una capa temática (entrada), genera para ésta misma, un raster interpolado de medias ponderadas (salida). En ILWIS, la media ponderada fue estimada con:
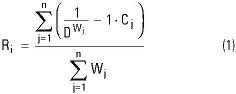
Donde:
R es el raster de salida i de medias ponderadas para las capas temáticas.
C es el n registro temático en las estaciones j.
W es el peso j asignado al n registro temático en las estaciones j, que condiciona la salida Ri.
D es la distancia Euclidiana entre n registros temáticos en las estaciónes j y el valor de salida vecino Ri.
En ILWIS, Wj fue estimada con la función de "distancia inversa", definida como 1/DWj (ecuación 1). Durante el proceso de interpolación, las salidas Ri se estructuraron utilizando a los pixeles que resultaron con un peso alto y significativo (Wj > 1); los pixeles con un peso bajo y no significativo (Wj < 1), se eliminaron y en su lugar, mediante la aplicación de una máscara cartográfica de color negro, se insertó un valor de cero en los pixeles que la constituyeron. Para este caso en particular, Wj se interpretó de la siguiente manera: Si Wj > 1 ocurre en Ri, entonces un determinado tipo de sustrato está presente y es predominante. Si Wj < 1 ocurre en Ri, entonces un determinado tipo de sustrato está presente, pero no es predominante.
Las salidas interpoladas para los diferentes tipos de sustrato, fueron utilizadas para representar cartográficamente, su disposición geográfica en la zona 90 de pesca. Las salidas interpoladas yi1 y yi2, se utilizaron para regionalizar a P(c). No se consideró la regionalización para ninguno de los tipos de sustrato, ya que éstos no presentaron variación temporal.
Regionalización. Con las salidas interpoladas de P(c), se desarrolló en ILWIS, un modelo de "regionalización" basado en la técnica de análisis de componentes principales (ACP) según Hair et al. (1999). Para este caso en particular, el ACP generó dos imágenes regionalizadas de P(c) o componentes principales, pero solo uno de éstas fue seleccionada para representar la regionalización final de P(c). De acuerdo con el criterio de la raíz latente (Hair et al., 1999), la componente principal seleccionada, fue aquella cuyo eigen-valor resultó mayor o igual a uno. Con la regionalización, fueron identificadas y delimitadas geográficamente, las aéreas con mayor y menor persistencia de P(c), prevalecientes en las temporadas agosto 2009 y octubre 2010 (yi1 y yi2, respectivamente). Esto último, se realizó con el programa computacional Statistica® versión 7.
Cartografía. Las salidas interpoladas para los diferentes tipos de sustrato y la regionalización de P(c), fueron desplegadas cartográficamente sobre de una imagen digitalizada del GT, acotada entre los 14º 36'-16° 33' N, 92º17'-95º11' O (Fig. 1). En ILWIS, ésta última fue creada con proyección UTM (zonas 15 y 16), elipsoide tipo Clarke 1866, datum tipo Norte Ámerica 1927 (NAD 27) y a una resolución de pixel de 113.446 m. La barimetría generada para la Figura 1, se obtuvo con el modulo "world_bathy_5min", incluida en programa computacional Widows Image Manager® versión 2012 (WIM por sus siglas en ingles).
RESULTADOS
La captura total del recurso camarón mostró un incremento gradual desde la sub-zona de pesca S-91 (160 Kg) hasta la S-95 (1030 Kg). Con este mismo rumbo, el incremento de la captura total del recurso camarón, coincidió con el descenso de las gravas (de 80.7 a 17.21%) (Fig. 2) y el incremento de los lodos (de 4.02 a 48.24%) (Fig. 2). Las arenas fueron observadas casi invariantes en todas las sub-zonas, sin embargo, en la S-94 y S-95, la composición porcentual de los lodos fue mayor a las arenas. En la S-91, la composición porcentual de las gravas fue mayor a las arenas (Fig. 2).
La captura total del recurso camarón evidenció una "correlación inversa" con las gravas y las gravas más arenas. Por otra parte, ésta mostró una "correlación directa" con los lodos, las arenas y las arenas más lodos. Únicamente la correlación obtenida con los lodos resultó significativa, mientras que, la menor de éstas y no significancia, fue generada con las arenas (Tabla 3).
Con base en los resultados inscritos en la Tabla 3 y Figura 2, se evidenció que la captura total del recurso camarón fue significativamente mayor y explicada en un 75.1%, por los lodos. Con un 55.3%, el descenso de ésta fue explicada por las gravas y en un 59.5%, por las gravas más arenas. Aunque la captura total del recurso camarón resultó alta y explicada en un 60.8% por las arenas más lodos, ésta según la Figura 2, sólo ocurrió cuando la composición porcentual de los lodos fue dominante. Debido a la presencia casi invariante de las arenas en la zona 90 de pesca, la correlación entre ésta y la captura total del recurso camarón, no pudo determinarse adecuadamente (7.3%).
Distribución ecológica. La hipótesis Ha fue aceptada; esto es, el recurso camarón evidenció en la zona 90 de pesca una distribución ecológica agregada (s 2 = 37.39, Y = 7.99, s 2 > Y ). La confrontación entre Yi y Y i, resultó significativa para la función de densidad probabilística "Binomial Negativa" (Xi 2 = 4366, X20.05 = 46.19 y p < 0.05) y no así para la función de densidad probabilística "Poisson" (Xi 2 = 1.6E+291, X 20.05 = 47.40 y p < 0.05). La P(c), fue estimada con la función de densidad probabilística "Binomial Negativa".
Análisis Geo-Estadísticos. Interpolación. Las gravas fueron predominantes a lo largo y ancho de la sub-zona de pesca S-91 y parte de la S-92; mientras que, en la S-93, éstas evidenciaron una menor predominancia. Las gravas fueron escasas en la S-94 y S-95 (Fig. 2, Fig. 3).
Las arenas estuvieron presentes en toda la zona 90 de pesca, excepto en la sub-zona de pesca S-91. Con una mayor predominancia, éstas fueron observadas hacia el frente oceánico, penetrando a nivel costero en la S-92 y S-94 (Fig. 4). En la S-92, las arenas se mezclan con las gravas, pero la composición porcentual dominante fue de las gravas (Fig. 2, Fig. 3, Fig. 4). En la S-93, S-94 y S-95, las arenas se mezclaron con los lodos, pero al avanzar hacia la S-95, la composición porcentual predominante fue de lodos (Fig. 2, Fig. 4, Fig. 5).
Debido a que las arenas permanecieron frecuentemente mezcladas con las gravas y los lodos (Fig. 4, Fig. 5), la correlación entre ésta y la captura total del recurso camarón resultó sub-estimada (Tabla 3), ya que en éstas mezclas, la menor composición porcentual fue de arenas (Figura 3, Figura 5); razón por la cual, en el análisis de correlación múltiple, prevaleció el efecto de la composición porcentual dominante (Tabla 3).
Los lodos estuvieron presentes en las sub-zonas de pesca S-93 y S-94, pero éstos fueron predominantes a lo largo y ancho en la S-95. Los lodos fueron escasos en la S-91 y S-92 (Fig. 5).
Regionalización. Dos imágenes regionalizadas de P(c) fueron generadas con el ACP, Cp1 con un eigen- valor de 1.4 y Cp2 con un eigenvalor de 0.5. La varianza explicada para Cp1 y Cp2 resultó de 67.80 y 32.20%, respectivamente. En la Figura 6 se muestra la regionalización final de P(c) o Cp1.
En las sub-zonas de pesca S-93 y S-94 y a lo largo y ancho de la S-95, la P(c) fue mayor. El valor intermedio de P(c), fue observado hacia el frente oceánico de la S-93 y S-94, así como en la mayor parte de la S-92. El menor valor de P(c), fue detectado en la S-91 (Fig. 6).
Con la regionalización de P(c), fueron identificadas y delimitadas en la zona 90 de pesca, las sub-zonas de pesca de mayor abundancia del recurso camarón, en términos de P(c) (Fig. 6). Con base en los resultados obtenidos, en la zona 90 de pesca fueron observados y delimitados dos sub-sistemas productivos de camarón. El primero de éstos fue denominado "Costa Oaxaqueña", delimitado entre Salina Cruz y la Boca de Tonalá (en el sistema lagunar Mar Muerto) (Fig. 1), en donde la P(c) (Fig. 6) y la captura total del recurso camarón fueron predominantemente bajos (Fig. 2). El segundo fue denominado "Costa Chiapaneca", delimitado entre la Boca de Tonalá y Puerto Chiapas (Fig. 1), en donde la P(c) (Fig. 6) y la captura total del recurso camarón fueron predominantemente altos (Fig. 2).
DISCUSIÓN
Para la zona 90 de pesca, los cambios en la abundancia y la distribución ecológica del recurso camarón, en relación con la disposición geográfica para diferentes tipos de sustrato, no había sido documentada. En este trabajo, se aborda por primera vez esta temática a nivel de recurso, ya que los registros de captura total obtenidos por el B/O UMAR, no fueron registrados con diferenciación entre especies comerciales; por esta razón, el tipo de distribución ecológica que el recurso camarón evidenció en el área de estudio, fue inferida, al igual que en otros trabajos enfocados para el recurso camarón del GT, considerando a las especies F. californiensis y L. vannamei como una sola población o un único recurso (Cervantes-Hernández et al., 2006; Cervantes-Hernández et al., 2012a). A pesar de que para el GT, existen registros de captura total con diferenciación de especies (Ramos-Cruz et al., 2006; Cervantes-Hernández, 2008; Cervantes-Hernández et al., 2008a, 2008b, 2010), éstos no pudieron ser utilizados, ya que no estuvieron georreferenciados.
Se ha demostrado que los organismos pertenecientes a los géneros Farfantepenaeus y Litopenaeus (Kensley, 1997), presentan hábitos conductuales preferenciales (Gunter, 1950; Fuss & Ogren, 1966; Fotheringham & Weissberg, 1979; Regnault, 1979; Dall & Smith, 1986; Rosas et al., 1995, 1999) para habitar sobre (Boddeke, 1983) y dentro (Williams, 1958) de sustratos de tipo limo-lodo-arcilloso y/o lodoso.
Para la plataforma marina al norte de California, E.U.A., autores como: Burkenroad (1939), Williams (1958) y Grady (1971), presentaron resultados consistentes, señalando que, F. aztecus y L. setiferus fueron predominantemente abundantes sobre de un sustrato limo-lodo-arcilloso, mientras que F. duorarum, exhibió la mayor abundancia sobre de un sustrato rocoso-calcáreo-arenoso. En el Golfo de México, Springer y Bullis (1954) reportaron esta misma tendencia para F. aztecus y L. setiferus, ambos fueron predominantemente abundantes sobre de un sustrato lodoso en la plataforma marina, que constituye la región Tamaulipas, Veracruz y Tabasco, México. Por otra parte en el Banco de Campeche, México, los autores reportaron una mayor abundancia de F. duorarum sobre de un sustrato calcáreo arenoso.
En sistemas lagunares, los hábitos conductuales de las distintas especies de camarón con relación al sustrato son persistentes. Zimmerman y Minello (1984), reportaron que en los estuarios de la isla de Galveston, Texas, E.U.A., los juveniles de F. aztecus fueron predominantemente abundantes sobre de un sustrato lodo-arcilloso, desprovisto de cubierta vegetal, mientras que, los juveniles de L. setiferus fueron predominantemente abundantes en el mismo tipo de sustrato, pero con cobertura vegetal. En la Laguna de Términos del Banco de Campeche, Gracia y Soto (1990), reportaron una alta abundancia de juveniles de L. setiferus sobre de un tipo de sustrato lodo-limo-arcilloso, rico en materia orgánica.
Lo anteriormente descrito, constituye una evidencia potencial, para sustentar que en la zona 90 de pesca, F. californiensis y L. vannamei, presentan los mismos hábitos conductuales preferenciales, para habitar sobre y dentro de sustratos de tipo limo-lodo-arcilloso y/o lodoso. De manera que, los resultados obtenidos, son pertinentes para explicar a nivel de recurso, la variación de la captura total y la P(c), en relación con la disposición geográfica de las gravas, arenas y lodos. No se obtuvo evidencia significativa, para sustentar que la captura total del recurso camarón (Fig. 2) y la P(c) (Fig. 6), fuesen predominantemente abundantes, en presencia de gravas, gravas más arenas y arenas (Fig. 2, Tabla 3).
En las sub-zonas de pesca con mayor predominancia de lodos (Fig. 5), el recurso camarón evidenció ampliamente la distribución ecológica agregada (Fig. 6), generando así un gradiente, en donde la captura total del recurso camarón (Fig. 2, Tabla 3) y la P(c) (Fig. 6), ascendieron gradualmente desde la sub-zona S-91 hasta la S-95. El gradiente ascendente resultó consistente con Ramos-Cruz (2009), quien durante la temporada de veda, entre junio y agosto 1999, reportó incrementos en la abundancia y la longitud total del recurso camarón, conforme los arrastres de veda avanzaron desde la S-91 hacia la S-95. Para este caso particular, la longitud total se incrementó de 12.9 (en la S-91) a 16.9 cm (en la S-95).
Con base en la disposición de las gravas, arenas y lodos, en la zona 90 de pesca, fueron observados y delimitados dos sub-sistemas productivos de camarón, la "Costa Oaxaqueña" con un bajo nivel de captura total y P(c), y la "Costa Chiapaneca", con un alto nivel de captura total y P(c).
La disposición geográfica de los sedimentos en el GT, ha sido reportada por autores como: Morales de la Garza y Carranza-Edwars (1995), Tapia-García et al. (2007) y Machain-Castillo et al. (2008). Para un propósito comparativo, los resultados obtenidos por estos autores, fueron acotados a la zona 90 de pesca (Fig. 1) y dentro de ésta, la disposición geográfica de los lodos, resultó consistente, concluyendo que, los lodos a partir de la sub-zona de pesca S-93, tienden a formar parches paralelos a la línea de costa, cuya extensión se incrementa hacia el sureste de la "Costa Chiapaneca", con rumbo al Río Suchiate, el de mayor influencia de descarga fluvial, según Morales de la Garza (1990).
En comparación con la "Costa Oaxaqueña", la "Costa Chiapaneca" registra una mayor actividad de descarga fluvial (Machain-Castillo et al., 2008), con una presencia de lodos dominante y persistente, constituidos principalmente de cuarzo, mica, polen y altos contenidos de materia orgánica. La descomposición continua de ésta última, ocasiona que la presencia de foraminíferos sea muy baja, particularmente entre la S-93 y S-95. Lo anterior está relacionado con las conclusiones de Tapia-García et al. (2007), quienes señalaron que durante la estación de lluvias en la "Costa Chiapaneca", predomina la arena lodosa con bajas cantidades de grava, así como valores bajos de salinidad y pH. Estas características hidrológicas son particularmente importantes en el caso de los camarones peneidos, ya que los incrementos en la descarga fluvial, favorecen positivamente la abundancia y la talla (Ruello, 1973); así como también, la magnitud del reclutamiento marino (Penn & Caputi, 1986), ya que el aporte continuo de sedimentos, incrementa la turbidez, que a su vez, disminuye la tasa de mortalidad por depredación (Cervantes-Hernández, 1999).
Para la zona 90 de pesca, se evidenció que las arenas son el tipo de sustrato dominante, con una disposición geográfica paralela a la línea de costa, penetrando a nivel costero, como se indicó en la Figura 4. Estos resultados, fueron consistentes con Morales de la Garza y Carranza-Edwars (1995) y Tapia-García et al. (2007). Por otra parte y particularmente para la sub-zona de pesca S-91, la disposición geográfica de las arenas, aunque presentes, resultaron con un Wj < 1 (Fig. 4), lo que demuestra, estadísticamente, que éstas no fueron predominantes. Veinte años antes a la realización del presente trabajo, los autores antes citados, reportaron la presencia de gravas y arenas frente al sistema lagunar Huave (en la S-91), sin embargo, no se evalúo la composición porcentual dominante. Para este caso en particular, el uso de la composición porcentual y el peso Wj, condicionaron las diferencias entre la comparación de los resultados bibliográ- ficos.
La dinámica oceanográfica del GT está bien estudiada (Blackburn, 1962; Monreal-Gómez & Salas de León, 1998; Farber-Lorda et al., 2004), pero la influencia de los sistemas lagunares sobre la disposición geográfica de los diferentes tipos de sustrato en la zona 90 de pesca, no ha sido estudiada a fondo. Al respecto, Cervantes-Hernández et al. (2012a) documentaron los episodios de cierre y abertura de la "Boca de San Francisco" en el sistema lagunar Huave (Fig. 1). Luego entonces, es importante destacar, que durante la realización de los cruceros oceanográficos MIMAR III, IV, V y FIQUIMBI-I (Garza & Carranza-Edwars, 1995; Tapia-García et al., 2007), la boca-barra permaneció abierta, posteriormente, se mantuvo cerrada entre mayo 2006 y agosto 2010, reabriéndose, nuevamente desde septiembre 2010 a la fecha. Resaltando entonces, que durante la realización del crucero oceanográfico CHIS-COYVER-0109, la "Boca de San Francisco" permaneció cerrada.
Anteriormente, se destacó al Río Suchiate como un elemento de gran influencia sobre la disposición geográfica de los lodos en la "Costa Chiapaneca" (Morales de la Garza, 1990). Sin embargo y a pesar de su gran dimensión (100,000 ha) (Espinoza et al., 2011), el sistema lagunar Huave no ha sido identificado como un elemento de influencia potencial, sobre la disposición geográfica de los sedimentos en la "Costa Oaxaqueña". Resalta el hecho de que durante la abertura de la "Boca de San Francisco", se reportó la presencia de gravas y arenas en la sub-zona de pesca S-91, mientras que, durante el periodo en el que se mantuvo cerrada, se evidenció la predominancia de gravas.
Es importante considerar, que la predominancia de las gravas en la sub-zona de pesca S-91, responde en parte, a las causas explicadas por Cervantes-Hernández et al. (2012a), quienes adicionalmente, reportaron una reducción del 75% en la entrada hídrica, que históricamente desembocó al oeste del Mar Tileme, en el sistema lagunar Huave; y que actualmente, ha sido desviada para alimentar al Distrito de Riego 19, para beneficiar a los municipios de San Blas Atempa, San Pedro Huilotepec, Santa María Xadani y Juchitán de Zaragoza, Oaxaca. México. Con una menor descarga fluvial proveniente del sistema lagunar Huave, la presencia de las arenas en la sub-zona de pesca S-91 resultó menor. Lo que demuestra, tal y como lo señalaron Morales de Garza y Carranza-Edwars (1995); que las condiciones hidrológicas locales a lo largo de la línea de costa en el GT, pueden influenciar la distribución geográfica de los sedimentos.
AGRADECIMIENTOS
Este trabajo fue desarrollado en la Universidad del Mar, Puerto Ángel, Oaxaca, con el apoyo económico del convenio 2IR1104. Agradecimientos para la Estación de Investigación Oceanográfica (SEMAR) y al Centro Regional de Investigación Pesquera con sede en Salina Cruz, Oaxaca. Así también a Ramón Tapia Martínez, José Luis Ramos Espinoza, Elmer Ventura Palomeque, Martín Zúñiga Ramírez, Saúl Serrano Guzmán, Isabel Gallardo Berumen, María Auxilio Esperanza Álvarez y a los revisores anónimos por las sugerencias y comentarios que mejoraron el manuscrito.
REFERENCIAS
Aragón-Noriega, E. A., C. Cervantes-Valle, A. R. García-Juarez & L. E. Calderón-Aguilera. 1999. Distribución y abundancia de la población desovante de camarones del norte del Golfo de California durante el verano de 1996. Ciencia y Mar 3 (9): 37-48. [ Links ]
Blackburn, M. 1962. An oceanographic study of the Gulf of Tehuantepec. Fish and Wildlife Service. Technical Report, U.S. 404: 1-28. [ Links ]
Boddeke, R. 1983. Survival strategies of penaeid shrimps and their significance for shrimp culture. First International Conference on Warmwater Aquaculture-Crustacean. 9-11 February, Laine, Hawaii, pp. 514-523. [ Links ]
Burkenroad, M. D. 1939. Further observations on Penneidae of the Northern Gulf of Mexico. Bulletin of the Bingham Oceanographic Collection 6 (6): 1-62. [ Links ]
Cervantes-Hernández, P. 1999. Relaciones stock-reclutamiento del camarón rosado Farfantepenaeus duorarum (Burkenroad, 1939) en el Banco de Campeche. Tesis de Maestría en Ciencias (Biología Marina), Instituto de Ciencias del Mar y Limnología, UNAM, México, 37 p. [ Links ]
Cervantes-Hernández, P., S. Ramos-Cruz & A. Gracia. 2006. Evaluación del estado de la pesquería de camarón en el Golfo de Tehuantepec. Hidrobiológica 16 (3): 233-239. [ Links ]
Cervantes-Hernández, P. 2008. Method to obtain indices of abundance in the population of brown shrimp from the Gulf of Tehuantepec, Oaxaca, Mexico. Revista de Biología Marina y Oceanografía 43 (1): 111-119. [ Links ]
Cervantes-Hernández P., M. I. Gallardo-Berumen, S. Ramos-Cruz, M. A. Gómez-Ponce & A. Gracia. 2008a. Análisis de las temporadas de veda en la explotación marina de camarones del Golfo de Tehuantepec, México. Revista de Biología Marina y Oceanografía 43 (2): 285-294. [ Links ]
Cervantes-Hernández, P., B. Sánchez-Meraz, A. Frías-Velasco, J. S. Serrano-Guzmán, S. Ramos-Cruz & A. Gracia. 2008b. Variación interanual de la abundancia de Farfantepenaeus californiensis (Holmes 1900) en el Golfo de Tehuantepec. Hidrobiológica 18 (3): 215-226. [ Links ]
Cervantes-Hernández, P., A. Flores-Gómez, S. J. Serrano-Guzmán, S. Ramos-Cruz & M. I. Gallardo-Berumen. 2010. Historical exploitation and evaluation of brown shrimp fishery Farfantepeneaeus californiensis (Decapoda, Dendrobranchiata) in the Gulf of Tehuantepec, Oaxaca, Mexico. Pan-American Journal of Aquatic Sciences 5 (4): 486-494. [ Links ]
Cervantes-Hernández, P., M. I. Gallardo-Berumen & S. J. Serrano-Guzmán. 2012a. Explotación y distribución de la captura artesanal de camarón en el sistema lagunar Huave, Oaxaca, México. Revista de Ciencias Marinas y Costeras (4): 67-81. [ Links ]
Cervantes-Hernández, P., M. A. Gómez-Ponce & P. Torres-Hernández. 2012b. Marine and lagoon recruitment of Litopenaeus vannamei (Boone, 1931) (Decapoda: Penaeidae) in the "Cabeza de Toro - La Joya buenavista" lagoon system, Chiapas, Mexico. CICIMAR Oceánides 27 (2):51-58. [ Links ]
Dall, W. & D. Smith. 1986. Oxygen consumption and ammonia-N excretion in fed and starved tiger prawns Penaeus esculentus Haswell. Aquaculture 55: 23-33. [ Links ]
Espinoza-Tenorio, A., L. C. Bravo-Peña, J. S. Serrano-Guzmán, J. A. Ronsón-Paulín, M. A. Ahumada, P. Cervantes-Hernández, E. Robles-Zavala, M. P. Fuentes & M. I. Gallardo-Berumen. 2011. La diversidad étnica como factor de planeación pesquera artesanal: Chontales, Huaves y Zapotecas del Istmo de Tehuantepec, Oaxaca, México. In: Alcala, G. & A. Camargo (Eds.). Pescadores en América Latina y el Caribe: espacio, población, producción y política. Vol. 1. Facultad de Ciencias Unidad multidiciplinaría de Docencia e Investigación-SISAL, UNAM. pp. 167-216. [ Links ]
Farber-Lorda, J., M. F. Lavin & M. A. Guerrero-Ruiz. 2004. Effects of wind forcing on the trophic conditions, zooplankton biomass and krill biochemical composition in the Gulf of Tehuantepec. Deep Sea Research Part II 51: 601-614. [ Links ]
Folk, R. L. 1969. Petrología de rocas sedimentarias. Instituto de Geología, UNAM. 405 p. [ Links ]
Fotheringham, N. & G. Weissberg. 1979. Some causes, consequences and potential environmental impacts of oxygen depletion in the northern Gulf of Mexico. Proceedings 11th Annul Offshore Technical Conference 4 (3611): 2205-2208. [ Links ]
Fuss, M. & L. Ogren. 1966. Factors affecting activity and burrowing habits of the pink shrimp, Penaeus duorarum Burkenroad. Biological Bulletin 130 (2): 170-191. [ Links ]
Gallegos-García, A. & J. Barberán-Falcon. 1998. Surgencia eólica. In: Tapia-García, M. (Eds.). El Golfo de Tehuantepec: el ecosistema y sus recursos. México, pp. 27-34. [ Links ]
Gracia, A. & L. Soto. 1990. Population study of the penaeid shrimp of Términos lagoon, Campeche, México. Anales del Instituto de Ciencias del Mar y Limnología 17 (2): 241-255. [ Links ]
Gracia, A. 1992. Explotación y manejo del recurso camarón. Ciencia y desarrollo 18 (106): 82-95. [ Links ]
Grady, J. R. 1971. The distribution of sediment properties and shrimp catch on two shrimping grounds on the continental shelf of Mexico. Proceedings of Gulf and Caribbean Fisheries Institute 23: 139-148. [ Links ]
Gunter, G. 1950. Seasonal population changes and distribution as related to salinity of certain invertebrates of Texas Coast, including commercial shrimp. Publications of the Institute of Marine Science 1: 7-51. [ Links ]
Hair, F., J. Anderson, L. Tatham & C. Black. 1999. Multivariate data analysis. Prentice Hall, New Jersey. 542 p. [ Links ]
Krebs, C. J. 1999. Ecología. Estudio de la distribución y la abundancia. Harla, México. 753 p. [ Links ]
Ludwig, J. A. & J. F. Reynolds. 1988. Statistical ecology. John Wiley & Sons, U.S.A. 337 p. [ Links ]
Machain-Castillo, M. L., M. A. Monreal-Gómez, E. Arellano-Torres, M. Merino-Ibarra & G. González-Chávez. 2008. Recent planktonic foraminiferal distribution patterns and their relation to hydrographic condition of the Gulf of Tehuantepec, Mexican Pacífic. Marine Micropaleontology 66: 103-119. [ Links ]
Monreal-Gómez, M. A. & D. A. Salas de León. 1998. Dinámica y estructura termohalina. In: Tapia-García, M. (Eds.). El Golfo de Tehuantepec: el ecosistema y sus recursos. México, pp. 13-26. [ Links ]
Morales de la Garza, E. 1990. Estudio de sedimentos fosfatados en el Golfo de Tehuantepec, México. Tesis de Maestría en Ciencias (Oceanografía Geológica), UNAM, México, 113 p. [ Links ]
Morales de la Garza, E. A. & A. Carranza-Edwars. 1995. Sedimentos fosfatados en el Golfo de Tehuantepec. Hidrobiológica 5 (1-2): 26-35. [ Links ]
Penn, J. W. & N. Captuti. 1986. Spawning stock-recruitment relationships and environmental influences on the tiger prawn Penaeus esculentus fishery in Exmouth Gulf, Western Australia. Australian Journal of Marine and Freshwater Research 37: 491-505. [ Links ]
Ramos-Cruz, S. 2009. Tallas, sexo y desarrollo gonádico de Litopenaeus vannamei (Crustacea: Decapoda: Penaeidae), durante la veda de 1999 en el Golfo de Tehuantepec, México. Ciencia Pesquera 17 (2): 29-39. [ Links ]
Ramos-Cruz S., B. Sánchez-Meraz, F. Carrasco-Ayuso & P. Cervantes-Hernández. 2006. Estimación de la tasa de mortalidad natural de Farfante penaeus californiensis (Holmes 1900) y Litopenaeus vannamei (Boone 1931) en la zona costera del Golfo de Tehuantepec, México. Revista de Biología Marina y Oceanografía 41 (2): 221-229. [ Links ]
Ramos-Santiago, E., J. M. Ramírez-Gutiérrez, R. Mendoza-Rodríguez & M. Tapia-García. 2006. Reproducción, distribución y abundancia del pez Pseudupeneus Grandisquamis (Perciformes: Mullidae), en el Golfo de Tehuantepec, México. Revista de Biología Tropical 54 (4):1103-1112. [ Links ]
Regnault, M. 1979. Ammonia excretion of sand shrimp Crangon crangon (L) during the moult cycle. Journal of Comparative Physiology 133: 199-204. [ Links ]
Rosas, C., A. Sánchez & E. Díaz. 1995. Oxygen consumption and ammonia excretion of Penaeus setiferus, P. schmitti, P. duorarum and P.notialis postlarvae fed purified test and diets: effect of protein level on substrate metabolism. Aquatic Life Research 8: 161-169. [ Links ]
Rosas, C., E. Martínez, G. Gaxiola, R. Brito, A. Sánchez & L. Soto. 1999. The effect of dissolved oxygen and salinity on oxygen consumption, ammonia excretion and osmotic pressure of Penaeus setiferus (Linnaeus) juveniles. Journal of Experimental Marine Biology and Ecology 234: 41-57. [ Links ]
Ruello, N. 1973. The influence of rainfall on the distributions and abundance of the school prawn Metapenaeus macleayi in the Hunter river region (Australia). Marine Biology 23: 221-228. [ Links ]
Ruiz-Luna, A., Meraz-Sánchez, R & J. Madrid-Vera. 2010. Abundance distribution patterns of commercial shrimp off northwestern Mexico modeled with geographic information systems. Ciencias Marinas 36 (2): 107-120. [ Links ]
Springer, S. & H. Bullis. 1954. Exploratory shrimp fishing in the Gulf of Mexico. Summary Report for 1952-1954. Commercial Fisheries Review 16 (10): 1-16. [ Links ]
Tapia-García, M., M. C. García-Abad, A. Carranza-Edwards & F. Vázquez-Gutierrez. 2007. Environmental characterization of the continental shelf of the Gulf of Tehuantepec, Mexico. Geofísica Internacional 46 (4): 249-260. [ Links ]
Uriel, E. 1995. Análisis de datos, series temporales y análisis multivariante. Universidad de Valencia, Madrid. 433 p. [ Links ]
Williams, A. 1958. Substrates as factor in shrimp distribution. Limnology and Oceanography 3 (3): 283-290. [ Links ]
Zimmerman, R. & J. Minello. 1984. Densities of Penaeus aztecus, Penaeus setiferus, and other natant macrofauna in Texas SALT marsh. Estuaries 7 (4a): 421-433. [ Links ]

