










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkHidrobiológica
Print version ISSN 0188-8897
Hidrobiológica vol.22 n.3 Ciudad de México Sep./Dec. 2012
Artículos
Variación mensual de los estadios gonádicos de Sagitta euneritica, (Chaetognatha) en el complejo lagunar, Bahía Magdalena, Baja California Sur, México
Monthly variation of gonadic stages of Sagitta euneritica (Chaetognatha) in The Bahía Magdalena lagoon complex, Baja California Sur, Mexico
Cota-Meza María Soledad,1 María Ana Fernández-Álamo,2 José Nicolás Álvarez-Cadena†3 y Eduardo González-Rodríguez4
1 Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas. Laboratorio de Plancton y Ecología Marina. A. P. 592. La Paz, Baja California Sur, 23000. México
2 Universidad Nacional Autónoma de México, Facultad de Ciencias, Laboratorio de Invertebrados, Ciudad Universitaria, México, D. F, 04510. México
3(†) 1948-2009 Unidad Académica Foránea Puerto Morelos, ICMyL, UNAM. A. P. 1152, Cancún, Quintana Roo, 77501. México
4 Centro de Investigación Científica y de Educación Superior de Ensenada, Unidad La Paz, Miraflores 334, Fraccionamiento Bella Vista, La Paz, Baja California Sur, 23050. México. E-mail: mcota@ipn.mx
Recibido: 26 de enero de 2012.
Aceptado: 03 de septiembre de 2012.
RESUMEN
Se determinaron la abundancia y proporción de los estadios gonádicos del quetognato Sagitta euneritica en el complejo lagunar Bahía Magdalena (CLBM), Baja California Sur, México. Las recolectas se realizaron durante los meses de marzo, junio, julio, agosto, septiembre y noviembre de 1982. S. euneritica fue la especie numéricamente dominante y con más amplia distribución, aportando el 81% de la abundancia total del grupo. S. euneritica se encontró principalmente en los estadios juveniles 0 (34%), I y II (con 21 y 40%), seguidos por los estadios III y IV con porcentajes menores, implicando con ello que los organismos maduros están probablemente en la zona costera inmediata. El análisis de la estructura por tallas mostró organismos pequeños desde junio hasta agosto, aumentando el número de éstos en septiembre y noviembre, asumiéndose que el CLBM actúa como una zona de crianza para esta especie. El análisis de varianza entre los porcentajes de los estadios por zonas y por mes, mostró que en marzo los organismos de la etapa I fueron significativamente diferentes (p = 0.05). Los individuos de S. euneritica se encontraron en un amplio intervalo de temperaturas, desde los 15.5 hasta los 29.5 °C. El número máximo de quetognatos en Bahía Magdalena se relacionó con las aguas de temperatura más fría y el máximo en Bahía Almejas con el aumento gradual de la temperatura en verano-otoño. La distribución de S. euneritica dentro del CLBM tendió a ser heterogénea, presentando además una estructura de tallas predominantemente alta de organismos inmaduros, probablemente como una estrategia de sobrevivencia de la especie.
Palabras clave: Estadios gonádicos, quetognatos, tallas, temperatura superficial del agua.
ABSTRACT
The abundance and the proportion of gonadal stages of the chaetognath Sagitta euneritica in the lagoon complex Bahia Magdalena (CLBM), Baja California Sur, Mexico, was determined. The sampling was made during March, June, July, August, September, and November 1982. Sagitta euneritica was the species numerically dominant and most widely distributed species, with 81% of the total abundance of the chaetognaths, and was found mainly in the juvenile stages 0 (34%), I and II (21% and 40%). The stages III and IV were found in low percentages, implying that mature organisms are likely found in the immediate coastal zone. The size-structure analysis showed small organisms from June to August, increasing in September and November; consequently that the CLBM can be the area acting as a breeding area for this species. The analysis of variance between the percentages of the stages by area and by month showed that in March the organisms of stage I were significantly different (p = 0.05). Individuals of S. euneritica were found in a wide temperature range from 15.5 to 29.5 °C. The maximum number of chaetognaths in the Magdalena Bay area was related to the cold water temperature and the maximum in the area of Almejas Bay was associated with a gradual increase of the water temperature in summer-autumn. The distribution of S. euneritica in the CLBM tends to be heterogeneous, also with a high size-structure of organisms predominantly immature, as a survival strategy of the species.
Key words: Gonadics stages, chaetognaths, sizes, water surface temperature.
INTRODUCCIÓN
Los quetognatos se encuentran entre los organismos carnívoros más numerosos del zooplancton en las regiones oceánicas, neríticas y costeras, contribuyendo substancialmente a la biomasa de esta comunidad pelágica (Kotori, 1972). Ejercen una significativa presión de depredación en otras poblaciones del zooplancton, principalmente en los copépodos (Feigenbaum & Maris, 1984; Álvarez-Cadena, 1993). En algunas regiones los quetognatos desempeñan un papel importante en el flujo vertical del carbono debido al tamaño relativamente grande de susheces (Dilling & Alldredge, 1993; Thuesen & Childress, 1993). Cuando sus poblaciones alcanzan abundancias elevadas pueden modificar la composición de la estructura de las tallas desus presas (Pearre, 1980), actuando como reguladores de la comunidad zooplanctónica (Villate, 1991).
En el complejo lagunar de Bahía Magdalena (CLBM) se han registrado doce especies de quetognatos pertenecientes a los géneros Sagitta Slabber, 1778 y Pterosagitta Ritter-Zahony, 1911 (Cota-Meza et al., 1992; Cota-Meza & Fernández-Álamo, 1998 y Cota-Meza, 2011), siendo Sagitta euneritica Alvariño, 1961, la más abundante. Hasta hoy la distribución de esta especie se restringe al Pacífico oriental, desde Cabo Mendocino, California (E.U.A.) hasta el Golfo de Panamá (Alvariño, 1965a, Cambrón 1981). Alvariño (1963) reporta esta especie en aguas de California y en la Bahía Vizcaíno, en la península de Baja California. Alvariño (1992) analiza la distribución batimétrica, diurna y nocturna de diez y siete especies de quetognatos en las aguas de California y Baja California, mostrando que S. euneritica es la segunda especie en importancia después de Sagitta bieri Alvariño, 1961.
Por su carácter nerítico S. euneritica es habitante común de los sistemas lagunares a todo lo largo del Pacifico mexicano (Pantoja-Vega, 1973; Vázquez, 1973; Gómez-Aguirre & Rivero-Beltrán, 1988). Cota Meza et al. (1992) estudiaron la distribución y la abundancia de las poblaciones de quetognatos en el CLBM durante un ciclo estacional en 1988-1989, encontrando que sus abundancias se incrementaban de abril a octubre con sus máximos en el último mes. A pesar de la importancia de S. euneritica en las redes alimenticias y de su aporte a la biomasa del zooplancton, se conoce relativamente poco de ella con respecto a la estructura de sus tallas y al desarrollo gonádico, por lo que el objetivo de este estudio es determinar las variaciones de estos parámetros y relacionarlos con la temperatura superficial del mar durante 1982, contribuyendo así al conocimiento de la biología de esta especie en el área de estudio.
MATERIALES Y MÉTODOS
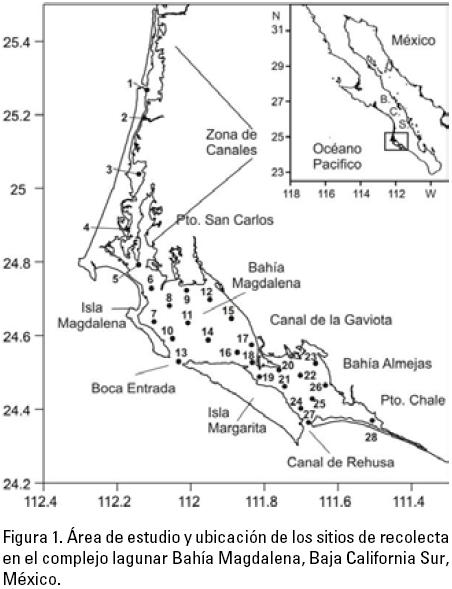
Área de estudio: El complejo lagunar de Bahía Magdalena (CLBM) carbono debido al tamaño relativamente grande de sus heces abarca un área total aproximada de 1390 km2, está localizado en (Dilling & Alldredge, 1993; Thuesen & Childress, 1993). Cuando sus el estado de BCS en la costa occidental de la península de Baja poblaciones alcanzan abundancias elevadas pueden modificar la California, entre los 24° 15’ y 25° 20’ N y 111° 30’, 112° 15’ W (Fig. 1). Este complejo se divide en tres zonas: 1) la parte noroccidental denominada Zona de Canales, de forma irregular, compuesta por esteros, lagunas y canales; con una profundidad promedio de 3.5 m y una área total de 137 km², 2) la región central que es propiamente la Bahía Magdalena, con una superficie de 883 km2, comunicada con el Océano Pacífico por una boca de 5.6 km de amplitud y que en su parte más profunda (boca) alcanza los 40 m; y 3) la zona suroccidental, denominada Bahía Almejas que tiene un área de 370 km2 y se abre al Océano Pacífico a través de dos bocas de poca profundidad y se comunica con Bahía Magdalena por medio de un canal de 2.5 km de ancho y una profundidad de 3 m (Álvarez-Borrego et al., 1975; Funes-Rodríguez et al., 2007). El sistema lagunar tiene una batimetría irregular, con zonas de canales y bancos superficiales, que favorecen una amplia variación en las condiciones físico-químicas, debido a un mayor calentamiento y evaporación en las zonas bajas y un mayor transporte de agua marina en las zonas más profundas y sin un considerable aporte continental (Sánchez-Montante et al., 2007).
Sánchez-Montante (2004), registran que Bahía Magdalena y Bahía Almejas presentan mareas tipo mixtas semidiurnas y la estructura termohalina en la mayor parte del sistema lagunar, corresponde a una zona bien mezclada en el plano vertical. Cervantes-Duarte et al. (2010) realizaron mediciones de las concentraciones de nitrógeno inorgánico disuelto (NID): nitrito (NO2), nitrato (NO3), amonio (NH4), fosfato (PO4), temperatura (T), salinidad (S) y concentración de clorofila a (Cla), en dos condiciones ambientales en Bahía Magdalena: fría (febrero, abril y junio) y cálida (agosto, octubre y diciembre). La relación de la temperatura y de la salinidad mostró que las aguas más frías y menos salinas generalmente se encuentran en profundidades de >10 m, asociadas a las aguas provenientes de la plataforma continental adyacente, mientras que las más cálidas y salinas, se localizan en la zona de canales donde prevalecen las condiciones antiestuarinas.
Gómez-Gutiérrez et al. (2007) demostraron con datos hidrográficos e hidroacústicos que el frente de marea fue más intenso en las mareas vivas, transportando el zooplancton desde la plataforma continental adyacente, hasta aproximadamente 6 km al interior de la región de Bahía Magdalena.
Trabajo de campo y laboratorio. Los arrastres de zooplancton se realizaron en el CLBM del 23 al 25 de marzo; del 7 al 10 de junio; del 20 al 23 de julio; del 9 al 11 de agosto; del 7 al 10 de septiembre y del 17 al 19 de noviembre de 1982.
En los primeros cuatro meses, se usó una red cónica con bridas al frente, de 0.30 m de diámetro de boca, un metro de largo, con luz de malla de 333 micras y provista de un flujómetro digital (General Oceanics). En septiembre y noviembre se utilizó una red cónica de 0.60 m de diámetro en la boca. De acuerdo con las bitácoras, en noviembre, el flujómetro presentó fallas, por lo que únicamente se presentan las abundancias de S. euneritica Alvariño, 1961 para marzo, junio, julio y agosto. Todos los muestreos fueron realizados durante el día, cerca de la superficie y siguiendo una trayectoria circular, durante 5 minutos a una velocidad aproximada de 4 km h-1. Los organismos recolectados fueron fijados con una solución de formaldehído hasta alcanzar una concentración del 4% en agua de mar, neutralizada con borato de sodio (Steedman, 1976). Asimismo se registró la temperatura superficial con un termómetro de inmersión. En el laboratorio se estimó la biomasa zooplanctónica por el método de volumen desplazado (Steedman, 1976). Dependiendo del volumen de biomasa, las muestras fueron fraccionadas con un separador Folsom para obtener alícuotas con un volumen aproximado de 10 ml. Las muestras con volúmenes inferiores a 15 ml se analizaron en su totalidad. La abundancia por muestra se normalizó a número de organismos por 100 m3 de agua filtrada según Fleminger (1964).
Los estadios de desarrollo gonádico de S. euneritica Alvarño, 1961, se determinaron a partir de 40 individuos tomados al azar de cada muestra. Los estadios I, II, III, y IV se definieron siguiendo los criterios propuestos por Alvariño (1963). En el estadio I los testículos y las vesículas seminales no aparecen todavía. Los ovarios se extienden hasta los dos tercios posteriores de la extensión de las aletas sobre el tronco. En el estadio II los testículos son visibles. Las vesículas seminales inician su desarrollo. Los ovarios llegan hasta la altura del borde anterior de las aletas posteriores. En el estadio III los testículos llenan por completo la aleta caudal. Las vesículas seminales están ya desarrolladas y los ovarios alcanzan el borde posterior de las aletas posteriores. En el estadio IV la cola está parcialmente vacía de esperma, las vesículas seminales están plenas y los ovarios llegan hasta la mitad o más allá de este nivel a la altura de las aletas anteriores. Se adiciona el estadio 0 equivalente a lo propuesto por Reeve (1970), en donde los testículos, las vesículas seminales y los ovarios están ausentes. La longitud del cuerpo se midió desde la punta de la cabeza hasta el final de la cola, excluyendo la aleta caudal (Zo, 1973).
Análisis de datos: La distribución de la frecuencia relativa en cada estadio de desarrollo gonádico, se presenta en función de la talla, agrupando a los organismos en intervalos de 1.0 mm. Para analizar las tallas de los quetognatos en relación a las zonas de muestreo, se agruparon las estaciones de la manera siguiente: para la zona de canales se agruparon las estaciones 1-4, para la zona de Magdalena las localidades 5-18 y para la zona de Almejas las estaciones 19-28, de acuerdo a la fisiografía del área. Se estimó el tamaño mínimo de muestra a medir para obtener estadísticos con a = 0.05. Asimismo, se realizó un análisis de variancia (p = 0.05), para saber si existe una distribución diferencial por estadio gonádico en términos de porcentajes en las diferentes zonas del CLBM, utilizando el programa Statistica (6.0). Por último se obtuvieron gráficas de la frecuencia de los ejemplares por estadio gonádico en intervalos de 2 °C, para cada zona del CLBM.
RESULTADOS
En todo el sistema lagunar la temperatura superficial mostró un amplio intervalo de variación mensual, con los valores más bajos en las bocas, particularmente en marzo (15.5 a 21.6 °C); los valores más altos ocurrieron en las zonas de canales y Bahía Almejas durante septiembre (23.7 a 29.5 °C). Los valores promedios de temperatura por mes aumentaron de marzo a septiembre y disminuyeron en noviembre. Los promedios de salinidad se obtuvieron en solo dos meses, observándose un ligero aumento de julio a noviembre (Tabla 1). En agosto y noviembre se observaron los valores mínimos de biomasa (ml/m3) y los más altos se observaron en julio y agosto. El mayor promedio de biomasa ocurrió en julio (56.6 ml/m3).
Sagitta euneritica (Alvariño, 1961) fue la especie más abundante y frecuente de la comunidad de quetognatos del complejo lagunar. La frecuencia relativa de S. euneritica fue de 81.2%, y Sagitta enflata (Grassi, 1881) obtuvo el 6.1%. Las especies de quetognatos que no alcanzaron el 5% de abundancia relativa fueron: S. bedoti (Beraneck, 1895), S. bieri (Alvariño, 1961), S. minima (Grassi, 1881), S. neglecta (Aida, 1897), S. pacifica (Tokioka, 1940), S. peruviana (Sund, 1961), S. pseudoserratodentata (Tokioka, 1939) y S. regularis (Aida, 1897). Las especies S. bedfordi (Doncaster, 1903), S. bipunctata (Quoy y Gaimard, 1827) y Pterosagitta draco (Krohn, 1853) mostraron porcentajes < 0.1%. La población de S. euneritica presentó amplias variaciones estacionales, alcanzando su máxima abundancia durante el verano con valores cercanos a los 60,000 organismos por 100 m3 en las bahías Magdalena y Almejas, disminuyendo hacia el otoño hasta 8000 organismos por 100 m3 (Fig. 2).
En cuanto a la distribución de S. euneritica por tallas, en las tres zonas que comprende el CLBM, fue posible observar la especie que estuvo presente en todos los meses analizados, con organismos de 1 mm de longitud de marzo hasta agosto y de 2.0 a 3.0 mm en septiembre y noviembre. La distribución de los organismos por estadios gonádicos por zona evidenció lo siguiente: en la zona de Canales, exceptuando junio, se observaron organismos de los cuatro primeros estadios y ausencia de ejemplares en estadio IV (Fig. 3). En la zona de bahía Almejas se presentaron organismos de los estadios gonádicos 0, I, y II, mientras que en la zona de Bahía Magdalena se observaron organismos en estadios 0 y II a partir de julio y hasta noviembre. En junio y agosto se registró un incremento substancial del estadio 0 en bahías Almejas y Magdalena, respectivamente (Tabla 2). Los estadios I y II se capturaron durante todo el periodo de estudio pero el estadio I, fue menos abundante en marzo en relación al conjunto de los meses analizados. Los organismos del estadio III fueron escasos aunque con excepción de septiembre, se capturaron en todos los meses de muestreo. Los organismos en estadio IV fueron escasos (menos de 15 organismos) en la zona de Bahía Magdalena durante marzo y noviembre con una talla promedio de 9.3 y 10.5 mm respectivamente (Tabla 2).
El análisis de variancia no mostró diferencias significativas (p = 0.05) entre la proporción de los estadios gonádicos de S. euneritica en las diferentes zonas del CLBM salvo para el estadio I (menos numeroso).
La distribución en tres dimensiones entre el número de quetognatos (Y), la temperatura °C (X), y el mes (Z), para las tres áreas del CLBM, se presenta en la figura 4. En general se observó que todos los estadios se separaron en dos grupos, el primero de cada grupo de izquierda a derecha (excepto el estadio l), se presentó en marzo y junio, cuando ocurrieron temperaturas bajas (Tabla 1), y el segundo grupo de cada gráfica se relacionó con los meses de julio, agosto, septiembre y noviembre cuando las temperaturas eran altas en el sistema lagunar. El estadio (E-1), aunque en menor número, estuvo representado en las tres áreas.
Se hace énfasis en la distribución del estadio gonádico (0) ya que fue de los más abundantes y posiblemente el más cercano a la eclosión. La distribución de los ejemplares del estadio gonádico 0 presenta tres máximos de abundancia: el primero en bahía Almejas con un intervalo de temperatura de 19.5 a 21.5 °C, el segundo y tercer máximos en Bahía Magdalena en los intervalos de 21.5 a 23. 5 °C y 23.5 a 24. 5°C, respectivamente (Fig. 4a).
El estadio gonádico I presentó dos máximos, el primero en Bahía Magdalena asociado a agua con temperatura de 19.5 a 21.5 °C. El segundo en la zona de canales en el intervalo 21.5 a 23.5 °C, (Fig. 4b). Los organismos en estadio gonádico II fueron tan abundantes como los recién eclosionados y se localizaron en aguas con temperatura de 23.5 a 25.5 °C en la zona de Bahía Almejas (Fig. 4c). Los organismos en estadio gonádico III presentaron tres valores máximos, dos de ellos en Bahía Magdalena en los intervalos 17.5 a 19.5 y 23.5 a 25.5 °C, y el otro máximo en Bahía Almejas, en el intervalo 23.5 a 25.5 °C, coincidiendo con los máximos de los organismos en estadio gonádico IV en Bahía Magdalena (Figs. 4d, e). La distribución del número de quetognatos de S. euneritica medidos en relación a la temperatura superficial para todas las épocas del año, mostró dos valores máximos, el primero en julio, asociado a temperatura de 20 °C, y el segundo en noviembre relacionado a la isoterma de los 24 °C, principalmente en Bahía Almejas (Fig. 5).
DISCUSIÓN
La distribución geográfica documentada para Sagitta euneritica Alvariño, 1961, comprende California (Alvariño, 1965b), Bahía Magdalena (Cota-Meza, 2011), el Golfo de Tehuantepec (Cambrón, 1981) y las aguas tropicales del Golfo de Panamá (Alvariño, 1965a; 1972). La variabilidad estacional de S. euneritica en el CLBM es similar a la registrada por Alvariño (1992) en las aguas de California y Bahía Vizcaíno. En el CLBM se observa un fuerte incremento de la especie a partir de la primavera hasta alcanzar su máximo poblacional durante el verano (Cota-Meza et al., 1992). En el presente estudio también se observó un incremento de primavera a verano asociado a valores altos de biomasa zooplanctónica (Tabla 1). En esta época la biomasa del zooplancton alcanza el máximo y la temperatura es alta (Saldierna-Martínez et al., 1987). Alvariño (1992) atribuye el máximo de verano a la actividad reproductora de la especie.
Kotori (1975) reporta que la talla de eclosión de S. elegans Verrill, 1873, es de 1.3 mm y aun cuando esta información no ha sido documentada para S. euneritica se considera altamente probable que su talla sea de alrededor de 1 mm, que fue el tamaño mínimo de los organismos capturado durante el presente estudio. Por lo tanto, se asume que los organismos obtenidos entre marzo y agosto (1 mm), septiembre (2.5 mm) y noviembre (<1.0 mm) probablemente cercanos a la eclosión. Se observaron también otros de talla similar como apendicularias y diatomeas, en las mismas muestras. Las capturas de quetognatos inmaduros (estadios 0 y I) durante todos los meses del muestreo permiten suponer que la especie se reproduce todo el año y que en ciertas épocas estos ejemplares pequeños contribuyen a la biomasa zooplanctónica del CLBM.
Es importante mencionar que los resultados sobre estadios gonádicos encontrados para S. euneritica en las costas de Sinaloa en estadio I, II (Pantoja-Vega, 1973) y laguna de Agiabampo en los estadios juveniles I y II (Gómez-Aguirre & Rivero Beltrán, 1988), muestran proporciones similares de estadios de desarrollo gonádico.
De acuerdo a Russell (1927,1931), el alto porcentaje de animales pequeños puede relacionarse con varios factores: que sean más resistentes a la irradiación solar, que en los estratos superiores exista mayor disponibilidad de alimento que las temperaturas superficiales más cálidas favorezcan un desarrollo más rápido (McLaren, 1963). Por otro lado, los organismos con un desarrollo gonádico más avanzado son capaces de realizar movimientos mayores en la columna de agua, ubicándose en capas inferiores durante el día y cerca de la superficie durante la noche (siendo importante el factor hora-luz). La escasez de adultos también pudo deberse a su capacidad para evitar la red, ya que tienen una mayor eficiencia natatoria o bien a que se distribuían por debajo de la capa de muestreo superficial. En este estudio los arrastres fueron diurnos y cerca de la superficie, lo que posiblemente no permitió una mayor captura de los estadios III y IV y por tanto se considera que existió un sesgo metodológico en términos de la representación de la estructura de tallas y de los estadios gonádicos que pudieran habitar en cada zona, por lo que para estudios futuros se recomienda realizar muestreos las 24 horas del día con redes de cierre-apertura en las regiones profundas, particularmente en Bahía Magdalena.
En los quetognatos, como en muchos otros organismos, se requiere cierto tiempo para producir una nueva generación, lo que depende de las variaciones de la temperatura y de la latitud de cada área (Dunbar, 1962). Reeve & Walter (1972) reportan que en condiciones experimentales, para que se produzca una generación de Ferosagitta (Sagitta) hispida Conant 1895, una especie tropical-subtropical, se requiere un lapso de 18 a 51 días. En S. elegans, Verrill, 1873 considerada una especie circumboreal (con una distribución mundial en aguas árticas y subárticas), se producen una o dos generaciones por año dependiendo de la temperatura del agua (Pierce, 1941; Sherman & Shaner, 1968; Tande 1983; Øresland, 1985; Álvarez-Cadena, 1993). En este contexto se deben considerar no solamente las pequeñas diferencias de temperatura que influyen en la producción de una nueva generación, sino también en el efecto térmico acumulativo (Sameoto, 1971; Dunbar, 1962). Con base en esto, se asume que el tiempo necesario para que S. euneritica produzca una nueva generación es de algunas semanas o meses.
En general se conoce que en áreas con temperaturas elevadas, una misma especie alcanza la madurez a tallas menores, mientras que a temperaturas bajas los organismos alcanzarán la talla máxima antes de llegar a la madurez gonádica reproductiva (Dunbar, 1962; McLaren, 1963). En este estudio encontramos que las tallas de reproducción de S. euneritica también se pueden relacionar con la inmigración de quetognatos que habitan en las zonas adyacentes como la plataforma continental. Posiblemente los organismos provenientes de las áreas norteñas, influenciadas por la corriente de California (de menor temperatura), sean de mayor talla y con un desarrollo gonádico más lento. Esta situación puede ser inversa para individuos de S. euneritica provenientes de las aguas más cálidas. En los estudios realizados por Alvariño (1963) en el Golfo de California se reporta que los individuos más grandes tendieron a presentar un desarrollo gonádico menos avanzado. Estos y otros aspectos (tales como las tasas de alimentación y de crecimiento) en el ciclo de vida de S. euneritica necesitan ser estudiados a mayor detalle. En el CLBM, S. euneritica mostró una distribución heterogénea con predominancia alta de organismos inmaduros, posiblemente como una estrategia de sobrevivencia de la especie, bajo las condiciones hidrológicas que ofrece esta área de estudio, para la crianza de estos depredadores planctónicos. Dilucidar las estrategias que presenta este depredador es de suma importancia porque el CLBM representa una área de reproducción de las sardinas, un recurso pesquero de suma importancia para la región, ya que en su etapa larval, dichos peces son susceptibles a la depredación de los quetognatos (Alvariño 1985), sin olvidar que los propios quetognatos sirven de alimento a las sardinas en su etapa adulta.
Es evidente que las condiciones hidrográficas tales como mareas, surgencias y frentes de marea tienen una influencia determinante en las variaciones estacionales de las abundancias de los quetognatos, como se ha demostrado para otros grupos zooplanctónicos importantes estudiados en esta área. Se con-firma que S. euneritica es una especie euritérmica al encontrase en amplios intervalos de temperatura superficial. Su población en el CLBM es heterogénea, con una predominancia de organismos inmaduros recién eclosionados (estadios 0 y 1) presentes a lo largo de todo el estudio. Las capturas permiten suponer que este quetognato se reproduce todo el año manteniendo una alta abundancia.
AGRADECIMIENTOS
Deseamos dedicar el presente trabajo al Dr. José Álvarez Cadena, quien participó haciendo sugerencias y revisiones en una versión previa del presente manuscrito. En homenaje a su memoria y en reconocimiento al apoyo incondicional brindado al primer autor para la culminación de su tesis de maestría. ¡Gracias Maestro!, por sus enseñanzas y por dejarnos gratos recuerdos de su amistad. Así mismo agradecemos al personal del Departamento de Plancton y Ecología Marina del CICIMAR-IPN, por la colaboración en la realización del presente trabajo. A los Drs. Jaime Gómez Gutiérrez y Ellis Glazier por sus valiosas sugerencias para mejorar este documento y al proyecto SIP 20120343 del IPN.
REFERENCIAS
Álvarez-Borrego, S. L., A. Galindo-Bect & A. Chee-Barragán. 1975. Características hidroquímicas de Bahía Magdalena, Baja California Sur. Ciencias Marinas 2: 94-110. [ Links ]
Álvarez-Cadena, J. N. 1993. Life cycle, abundance, gonadic stages and size frequency distribution of the chaetognath Sagitta elegans Verrill in the North-eastern Irish Sea. Estuarine Coastal Shelf Science 37: 15-25. [ Links ]
Alvariño, A. 1963. Quetognatos epiplanctónicos del Mar de Cortés. Re-vista de la Sociedad Mexicana de Historia Natural 24: 97-203. [ Links ]
Alvariño, A. 1965a. Chaetognaths. Oceanography and Marine Biology Annual Review 3: 115-194. [ Links ]
Alvariño, A. 1965b. Distributional atlas of chaetognatha in the California Current Region. During The CalCOFI monthly cruises of 1954 and 1958. California Cooperative Fisheries Investigations Atlas 3 (I-XIII): 1-291. [ Links ]
Alvariño, A. 1972. Zooplancton del Caribe, Golfo de México y regiones adyacentes del Pacífico. Memorias del IV Congreso Nacional de Oceanografía, México, 24 p. [ Links ]
Alvariño, A. 1985. Predation in the plankton realm; mainly with reference to thefish larvae. Investigaciones Marinas CICIMAR 2 (1): 72-77. [ Links ]
Alvariño, A. 1992. Distribución batimétrica, diurna y nocturna, de diez y siete especies de quetognatos, durante las cuatro estaciones del año 1969, en aguas de California y Baja California. Investigaciones Marinas CICIMAR 7: 1-169. [ Links ]
Cambrón, M. 1981. Estudio preliminar de las especies planctónicas del Phylum Chaetognatha en el Golfo de Tehuantepec, México. Tesis Profesional, Facultad de Ciencias, UNAM, México. 78 p. [ Links ]
Cervantes-Duarte, R., S. López-López, E. González-Rodríguez & S. Futema-Jiménez. 2010. Ciclo estacional de nutrientes, temperatura, salinidad y clorofila a en Bahía Magdalena, Baja California Sur, México. (2006-2007). CICIMAR Oceánides 25 (2): 111-120. [ Links ]
Cota-Meza, M. S. & M. A. Fernández-Álamo. 1998. Range extension for Sagitta peruviana (Chaetognatha: Sagittoidea) in Bahía Magdalena, Baja California Sur, México. Revista de Biología Tropical 46 (3): 847-848. [ Links ]
Cota-Meza, M. S., M. J. Haro-Garay & V. Massé-Zendejas. 1992. Distribución y abundancia de quetognatos en el complejo lagunar de Bahía Magdalena, Baja California Sur. México, durante el ciclo estacional 1988-1989. Investigaciones Marinas CICIMAR 7 (2): 47-59. [ Links ]
Cota Meza, M. S. 2011. Chaetognatha in the Bahía Magdalena lagoon complex, Baja California Sur, México: Species composition and assemblages. Journal of Environmental Biology 32 (4): 401-406. [ Links ]
Dilling, L. & A. L. Alldredge. 1993. Can chaetognath faecal pellets contribute significantly to carbon flux? Marine Ecology Progress Series 92: 51-58. [ Links ]
Dunbar, M. J. 1962. The life cycle of Sagitta elegans in Arctic and subarctic seas and the modifying effects of hydrographic differences in the environment. Journal of Marine Research 20: 76-91. [ Links ]
Feigenbaum, D. L. & R. C. Maris. 1984. Feeding in Chaetognatha. Annual Review of Oceanography and Marine Biology 22: 343-392. [ Links ]
Fleminger, A. 1964. Distributional Atlas of Calanoid Copepods in the California Current Region, part 1, California Cooperative Oceanic Fisheries Investigations Atlas 2: 1- 213. [ Links ]
Funes-Rodríguez, R., J. Gómez-Gutiérrez & R. Palomares-García (eds.). 2007. Estudios Ecológicos en Bahía Magdalena. Instituto Politécnico Nacional México, México, D. F. 311 p. [ Links ]
Gómez-Aguirre, S. & C. Rivero-Beltrán. 1988. Variación estacional de Sagitta euneritica (Chaetognatha) en la laguna de Agiabampo, México. Anales del Instituto de Biología Universidad Nacional Autónoma de México Serie Zoología 2: 697-706. [ Links ]
Gómez-Gutiérrez, J., S. Martínez & C. J. Robinson. 2007. Influence of tidal fronts on surface zooplankton aggregation and community structure in a subtropical bay, Bahia Magdalena, Mexico. Marine Ecology Progress Series 346: 109-125. [ Links ]
Kotori, M. 1972. Vertical distribution of chaetognaths in the northern North Pacific Ocean and Bering Sea. In: Takenouti, A. Y. (Ed.). Biological oceanography of the northern North Pacific Ocean. Idemitsu Shoten, Tokio. pp. 291-308. [ Links ]
Kotori, M. 1975. Morphology of Sagitta elegans (Chaetognatha) in early larval stages. Journal of Oceanography Society of Japan 31: 139-144. [ Links ]
McLaren, I. A. 1963. Effects of temperature on the growth of zooplankton, and the adaptive value of vertical migration. Journal of the Fisheries Research Board Canada 20: 685-727. [ Links ]
Øresland, V. 1985. Temporal size and maturity-stages distribution of Sagitta elegans and occurrence of other chaetognath species in Gullmarsfjorden, Sweden. Sarsia 70: 95-101. [ Links ]
Pantoja-Vega, A. 1973. Abundancia de quetognatos en las Bocas de Barrón y Chametla, Sinaloa, México, Tesis Profesional, Facultad de Ciencias, UNAM, México. 32 p. [ Links ]
Pearre, S. Jr. 1980. Feeding by Chaetognatha. The relation of prey size to predator size in several species. Marine Ecology Progress Series 3: 125-134. [ Links ]
Pierce, E. L. 1941. The occurrence and breeding of Sagitta elegans Verrill and Sagitta setosa J. Müller in parts of the Irish Sea. Journal of the Marine Biological Association of the United Kingdom 25: 113-124. [ Links ]
Reeve, M. R. 1970. The biology of Chaetognatha I.Quantitative aspects of growth and egg production in Sagitta hispida. In: J. H. (Ed.). Marine Food Chains. Steele, Oliver and Boyd, Edinburgh, pp. 168-189. [ Links ]
Reeve, M. R. & Walter, M. A. 1972. Conditions of culture, food size selection and the effects of temperature and salinity on growth rate and generation time in Sagitta hispida Conant. Journal of Experimental Marine Biology and Ecology 9: 191-200. [ Links ]
Russell, F. S. 1927. The vertical distribution of marine macroplankton V. The distribution of animals caught in the ring-trawl in the daytime in the Plymouth area. Journal of the Marine Biological Association of the United Kingdom 14: 557-608. [ Links ]
Russell, F. S. 1931. The vertical distribution of marine macroplankton. X. Notes on the behaviour of Sagitta in Plymouth area. Journal of the Marine Biological Association of the United Kingdom 17: 391-414. [ Links ]
Saldierna-Martínez, R., C. Sánchez & G. R. Vera. 1987. Estudios sobre los primeros estadios de vida de las sardinas crinuda (Opisthonema libertate), y Monterrey (Sardinops sagax) en Bahía Magdalena, Baja California Sur. I Descripción de huevos, II Distribución y Abundancia de huevos y larvas, III Mortalidad larval. Tesis Profesional, Universidad Autónoma de Baja California Sur, México. 217 p. [ Links ]
Sameoto, D. D. 1971. Life history, ecological production, and an empirical mathematical model of the population of Sagitta elegans in St. Margaret's Bay. Nova Scotia. Journal of the Fisheries Research Board of Canada 28: 971-985. [ Links ]
Sánchez-Montante, O. 2004. Hidrodinámica y transporte de masa en el sistema lagunar de Bahía Magdalena-Bahía Almejas, Baja California México: modelación y experimentación Tesis de Doctorado, Centro Interdisciplinario de Ciencias Marinas, La Paz, Baja California Sur, México. 237 p. [ Links ]
Sánchez-Montante, O., O. Zaitsev & M. Saldívar-Reyes. 2007. Condiciones hidrofísicas en el sistema lagunar Bahía Magdalena-Bahía Almejas. In: Funes-Rodríguez, R., J. Gómez-Gutiérrez & R. Palomares-García (Eds.).Estudios Ecológicos en Bahía Magdalena. Centro Interdisciplinario de Ciencias Marinas, La Paz Baja California Sur, México, pp. 1-28. [ Links ]
Sherman, K. & E. G. Shaner. 1968. Observations on the distribution and breeding of Sagitta elegans (Chaetognatha) in coastal waters of the Gulf of Maine. Limnology and Oceanography 13: 618-625. [ Links ]
Steedman, H . F. 1976. General and applied data on formaldehyde fixation and preservation of marine zooplankton. In: Steedman, H. F. (Ed.). Zooplankton Fixation and Preservation, Monographs on oceanographic methodology 4, Unesco Press, Paris, pp. 103-154. [ Links ]
Tande, K. S. 1983. Ecological investigations of the zooplankton community of Balsfjorden, northern Norway: Population structure and breeding biology of the chaetognath Sagitta elegans Verril. Journal of Experimental Marine Biology and Ecology 68: 13-24. [ Links ]
Thuesen, E. V. & J. J. Childress. 1993. Enzymatic activities and metabolic rates of pelagic chaetognaths: Lack of depth-related declines. Limnology and Oceanography 38 (5): 935-948. [ Links ]
Vázquez, C. M. 1973. Quetognatos en dos localidades de la costa mexicana del océano Pacífico. Su relación con elementos abióticos del ambiente y con postlarvas de camarón. Tesis Profesional, Universidad Autónoma de Morelos, México, 27 p. [ Links ]
Villate, F. 1991. Annual cycle of zooplankton community in the Abra Harbour (Bay of Biscay): abundance, composition and size spectra. Journal of Plankton Research 13 (4): 691-706. [ Links ]
Zo, Z. 1973. Breeding and growth of the chaetognath Sagitta elegans in Bedford Basin. Limnology and Oceanography 18: 750-756. [ Links ]

