










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.20 no.2 Ciudad de México may./ago. 2010
Artículos
Estructura comunitaria y trófica de las estrellas de mar (Echinodermata: Asteroidea) en arrecifes rocosos de Loreto, Golfo de California, México
Community and trophic structure of sea stars (Echinodermata: Asteroidea) in rocky reefs of Loreto, Gulf of California, Mexico
Betsabé Montserrat Luna Salguero y Héctor Reyes Bonilla
Universidad Autónoma de Baja California Sur. Departamento de Biología Marina. Apartado postal 19–B, CP 23080 La Paz, B.C.S., México e–mail: hreyes@uabcs.mx.
Recibido: 28 de octubre de 2009
Aceptado: 19 de julio de 2010
RESUMEN
Las estrellas de mar (Asteroidea) son invertebrados de gran importancia en hábitats rocosos y coralinos al ocupar diversos niveles de las cadenas tróficas y al actuar muchas veces como depredadores tope en esos ecosistemas. El conocimiento sobre este grupo en México es adecuado en los campos de la taxonomía y la biogeografía, pero existe muy poca información sobre la ecología y función de sus ensamblajes. El objetivo de este trabajo fue comparar la estructura comunitaria y trófica de los asteroideos de fondos rocosos en dos regiones del Golfo de California: Loreto y Ligüi (25.5° a 26.5°N). Los individuos fueron censados dentro de transectos de banda de 25 × 2 m (N = 106, a profundidades de 3 a 12 m) y se estimó la abundancia, riqueza de especies, diversidad (H'), y uniformidad (J'), así como el número de estrellas por gremio trófico (carnívoros, herbívoros y detritívoros), y de gremios presentes en cada transecto. Los resultados indican que las asociaciones de estrellas de mar de ambas zonas estuvieron dominadas por la especie Phataria unifascialis, y que la región de Loreto tuvo significativamente mayor riqueza, abundancia y diversidad de asteroideos que Ligüi, probablemente debido a que presenta mayor número de hábitat y recursos alimenticios. Con respecto a la composición trófica, en ambas localidades predominaron en número los herbívoros, luego los detritívoros y finalmente los carnívoros. La abundancia por transecto en cada uno de los niveles tróficos fue significativamente mayor en Loreto, y además se presentaron más grupos tróficos por transecto en esa localidad.
Palabras clave: Echinodermata, arrecifes rocosos, Parque Nacional Bahía de Loreto, índices ecológicos, gremios tróficos.
ABSTRACT
Sea stars (Asteroidea) are invertebrates with remarkable importance in rocky and coral habitats as they occupy several levels in trophic webs, and many times act as top predators in these ecosystems. The taxonomic and biogeographic knowledge about this group in México is adequate, but there is still limited information on the ecology and functioning of its assemblages. The objective of this study was to compare the community and trophic structure of rocky bottom asteroids in two regions of the Gulf of California: Loreto and Ligüi (25.5° to 26.5°N). Individuals were censused in belt transects 25 × 2 m (N = 106, at depths from 3 to 12 m), abundance, species richness, diversity (H') and evenness (J') was estimated, as well as the number of seastars per trophic guild (carnivores, herbivores and detritivores), and of guilds present per transect. The results indicate that starfish assemblages in both zones were dominated by the species Phataria unifascialis, and that Loreto had significantly higher richness, abundance and diversity of asteroids than Ligüi, probably as a consequence of higher number of habitats and food resources. In relation to the trophic composition, in both areas herbivores predominated, followed by detritivores and finally by carnivores. Abundance in each trophic level was statistically higher in Loreto, and also there were more trophic groups per transect at that location.
Key words: Echinodermata, rocky reefs, Bahía de Loreto National Park, ecological indices, trophic guilds.
INTRODUCCIÓN
Las estrellas de mar (Clase Asteroidea) son equinodermos que juegan papeles ecológicos de gran importancia, al aparecer en todas las franjas latitudinales, al ocupar diversos niveles de las cadenas tróficas y en especial actuar como depredadores tope en arrecifes rocosos y coralinos (Paulay, 1996). El éxito ecológico del grupo se atribuye a varias de sus características morfológicas y de historia de vida, incluyendo su crecimiento indeterminado, la ocurrencia de digestión extra e intra oral (que les da la posibilidad de acceder a una dieta muy diversa), la rapidez de detección y respuesta ante la presa, y la capacidad de sujeción al sustrato gracias a los discos suctores de los pies ambulacrales (Menge, 1982; Blake, 1989).
El conocimiento de los asteroideos en el Pacífico de México es amplio desde la perspectiva taxonómica (Caso, 1994; Cintra–Buenrostro, 2005), pero también hay buena información faunística, incluyendo listados de especies para muchas localidades (Bautista–Romero et al., 1994; Chávez–Dagostino et al., 2000; Zamorano y Leyte–Morales, 2005; Herrero–Pérezrul et al., 2008) y regiones (Solís–Marín et al., 2005; Honey–Escandón et al., 2008), así como estudios biogeográficos que han confirmado que las estrellas de mar siguen los patrones provinciales típicos del Pacífico oriental (Maluf, 1988), y además que su distribución está ligada a aspectos oceanográficos que limitan la presencia de ciertas especies (Cintra–Buenrostro et al., 2005). En años recientes, los aspectos de ecología comunitaria y trófica de este grupo han comenzado a recibir mayor atención; Reyes–Bonilla (1995) indicó que la abundancia de estrellas en las islas Socorro y San Benedicto (Archipiélago Revillagigedo, Colima; 18°N) era muy baja, y que los taxa más comunes fueron Mithrodia bradleyi Verrill, 1870 y Acanthaster planci (Linnaeus, 1758). Posteriormente, Caso et al. (1996) mencionaron que en la Bahía de Mazatlán (23°N) las zonas de fondo rocoso presentaron menor riqueza y abundancia de estrellas de mar que las de fondo blando y que la comunidad estuvo dominada en número por las especies Pharia pyramidatus (Gray, 1840) y Phataria unifascialis (Gray, 1840). Holguín–Quiñones et al. (2000) registraron que las especies dominantes en la Bahía de Loreto (25°N), en la costa oeste del Golfo de California fueron P. unifascialis, M. bradleyi y A. planci, mientras que Reyes–Bonilla et al. (2005) compararon la estructura comunitaria de asteroideos en cuatro zonas rocosas del oeste del golfo, indicando también que en Loreto se observó la mayor riqueza y abundancia de estrellas, y que P. unifascialis y P. pyramidatus fueron dominantes a lo largo de la región estudiada. González–Medina et al. (2006) encontraron 8 especies de estrellas de mar en la Isla Espíritu Santo, al suroeste del Golfo de California (24°N), e indicaron que P. unifascialis fue la especie más abundante (típica de zonas con baja intensidad de oleaje y alta cobertura de coral), seguida por Pentaceraster cumingi (Gray, 1840). Holguín Quiñones et al. (2008) observaron que en la Isla San José (24°N), al norte de La Paz, P. unifascialis también era la especie más común y abundante, seguida por P. cumingi y P. pyramidatus. Herrero–Pérezrul et al. (2008) confirmaron que dichos taxa también fueron dominantes en Bahía de Los Ángeles (29°N) y denotaron que los asteroideos tienen diferencias de abundancia y riqueza entre localidades, pero no en diversidad ecológica o distintividad taxonómica (Δ+). Finalmente, Ríos–Jara et al. (2008) mostraron que P. unifascialis y P. pyramidatus fueron también las estrellas más comunes en la Isla Isabel (20°N), seguidas por la corona de espinas A. planci, un famoso depredador coralino.
La mayoría de trabajos sobre ecología comunitaria de asteroideos se ha concentrado en localidades individuales, y no se han descrito los ensamblajes desde una óptica funcional. Por esa razón, el objetivo de este estudio fue comparar la estructura comunitaria y algunos aspectos de la ecología trófica de los asteroideos en hábitat rocosos de las secciones norte y sur de la Bahía de Loreto y sus alrededores, que es una de las regiones reconocidas como de mayor diversidad y abundancia de asteroideos en el Pacífico de México (Reyes–Bonilla et al., 2005). Sobre esta zona aún hay poca información disponible y además de las obras de Reyes–Bonilla et al. (2005) y Holguín–Quiñones et al. (2000) citadas anteriormente, Morgan y Cowles (1996) mostraron que P. unifascialis presentó tallas más pequeñas en Loreto que en Bahía de Los Ángeles (29°N), aunque en el primer sitio hay mayor presencia de juveniles; además los autores observaron que las bajas temperaturas (14 °C) generan estrés en este asteroideo, y sugieren que dicho factor regula su ocurrencia y abundancia en latitudes altas del golfo. Finalmente, Arizpe y Riosmena–Rodríguez (2007) mencionan la presencia de solo tres especies de este grupo en Loreto: P. cumingi, M. bradleyi y Nidorellia armata (Gray, 1840).
METODOLOGÍA
El presente estudio se realizó en zonas rocosas situadas en las cercanías de la ciudad de Loreto en octubre de 2007. Se efectuó un solo muestreo de alta intensidad en la época de otoño dado que estudios previos han indicado que las densidades poblacionales de los asteroideos son constantes e independientes de la temporada del año a lo largo del Golfo de California (Holguín–Quiñones et al., 2000, 2008; Reyes–Bonilla et al., 2005; Herrero Pérezrul et al., 2008), posiblemente debido a que son organismos de tiempo de vida largo y reclutamiento relativamente constante.
Se estableció un total de 18 sitios de muestreo, los cuales se arreglaron en dos localidades mayores: Loreto (10 sitios) y Ligüi (8 sitios; Fig. 1). La primera comprende el frente costero de San Nicolás hasta el área del Juncalito en su extremo sur, así como la Isla Coronados y la parte interior de Isla Carmen. La segunda abarca desde Puerto Escondido hasta Puerto Agua Verde, incluyendo las islas Danzante, Monserrat, Santa Catalina y la parte exterior de la Isla Carmen.

En cada sitio, empleando buceo autónomo se hicieron entre 5 y 6 censos de todos los asteroideos observados en transectos de banda de 25 × 2 m colocados en dirección paralela a la costa (50 m2 de superficie en cada uno; N = 106; Loreto N = 61 y Ligüi N= 45). Los conteos se realizaron a profundidades de 3 a 12 m ya que ahí se encuentran las mayores abundancias de asteroideos en el Golfo de California (Reyes Bonilla et al., 2005). La identificación de las especies se hizo siguiendo los criterios de Brusca (1980), Caso (1992) y Hickman (1998), y apoyados por ilustraciones de Gotshall (1987) y Kerstitch y Bertsch (2007).
Para el análisis comunitario, los datos de abundancia se analizaron en forma de número de individuos por transecto (equivalentes a ind/50 m2), y para cada unidad de muestreo se cuantificó la riqueza total de las especies de estrellas de mar. A partir de ahí se calculó el índice de diversidad de Shannon–Wiener (H' en base 10), y el de uniformidad o equidad de Pielou (J') el cual expresa qué tan homogéneamente se encuentran distribuidos los individuos de las diferentes especies en cada muestreo (Clarke y Warwick, 2001). Los valores de los índices fueron revisados con las pruebas de Kolmogorov–Smirnoff y Levene para determinar si mostraban normalidad y homoscedasticidad. El índice de uniformidad fue el único que cumplió con estos términos, mientras que el resto no mostraron estas condiciones, para ello se hizo la prueba t de Student para varianzas iguales y desiguales (o prueba de Welch), respectivamente, con el fin de determinar diferencias de los índices ecológicos entre las localidades de Loreto y Ligüi (Zar, 2009).
Posteriormente, se estimó el grado de semejanza entre las localidades de muestreo utilizando el índice de similitud de Bray–Curtis, el cual considera en conjunto la presencia y abundancia de las especies (Krebs, 1999). Luego, con base en la matriz se efectuó un análisis de ordenación con la técnica de escalamiento multidimensional no métrico (nMDS), la cual muestra los sitios de muestreo en un mapa de dos dimensiones y nos indica la distancia relativa entre dichos sitios. En el caso que nos atañe, para determinar la validez del arreglo se tomó en cuenta el nivel de stress de 0.10 (Mc. Cune et al., 2002). En adición, la matriz de Bray–Curtis se utilizó en un análisis de similitudes (ANOSIM; Warwick y Clarke, 2001), con el fin de denotar si la composición faunística de las estrellas de mar difería significativamente entre localidades.
La última parte del estudio se enfocó al análisis funcional. Para ello se clasificó cada especie dependiendo de su gremio trófico (carnívoro, herbívoro o detritívoro), usando los criterios de Cintra–Buenrostro et al. (2005). Hecho esto, se compararon las abundancias de individuos entre localidades pertenecientes a cada gremio trófico, así como el número de grupos tróficos presentes por transecto. En todos los casos, las comparaciones estadísticas entre localidades se hicieron por medio de la prueba t de Student, o su versión para varianzas desiguales (prueba de Welch), dependiendo si los datos eran homoscedasticos y normales.
RESULTADOS
Estructura comunitaria. Se identificaron un total de 1740 especímenes en los 106 censos, pertenecientes a 9 especies de asteroideos, incluidas en 9 géneros y 6 familias. De ellas, P. unifascialis fue la especie más abundante (52% del total de individuos observados) seguida por P. cumingi (22%) y P. pyramidatus (17%). Caso contrario, para las especies Asteropsis carinifera Lamarck, 1816 y Narcisia gracilis Clark, 1916, las cuales fueron representadas en los transectos por un individuo y un par de ejemplares, respectivamente (Tabla 1).
Analizando los índices ecológicos, la riqueza promedio para ambas localidades de estudio en conjunto fue de 2.96 ± 0.13 especies por transecto, y hubo diferencias entre ellas (t2,102.6 = 4.84, p < 0.001), siendo significativamente más rica la localidad de Loreto que la de Ligüi (Fig. 2a). Por otro lado, la abundancia promedio general de los asteroideos fue de 16.42 ± 1.29 ind/transecto, y también se dieron diferencias significativas de los valores de abundancia entre localidades (t2,100 8 = 3.74, p < 0.001), siendo Loreto donde hubo números más altos (Fig. 2b). En el análisis de diversidad (H') se obtuvo un valor promedio general de 0.32 ± 0.01 decits/ind y hubo diferencias significativas entre localidades (t2 102 6 = 3.1 1 , p = 0.002), ya que la localidad de Loreto fue la que tuvo mayor diversidad (Fig. 2c). Por último, la uniformidad (J') tuvo un promedio general de 0.67 ± 0.03, pero en dicho índice no se encontraron diferencias significativas entre localidades (t2,104 = 0.13, p < 0.898); esto indica que la abundancia relativa de individuos encontrados por especie está distribuida de manera similar en toda el área de estudio (Fig. 2d).
El análisis de ordenación no evidenció una separación clara de grupos de estaciones de muestreo (stress = 0.15; Fig. 3), y el ANOSIM tampoco fue significativo (R = 0.02; p = 0.356), todo lo cual corrobora la ausencia de separaciones entre las estaciones de muestreo y confirma que las asociaciones de asteroideos no presentaron diferencias cualitativas entre las dos localidades de estudio.
Grupos Tróficos. Como se observa en la Figura 4, la abundancia de carnívoros, herbívoros y detritívoros fue mayor en la localidad de Loreto que en Ligüi. No obstante, en ambos sitios las estrellas de mar con hábitos herbívoros fueron los que predominaron (Loreto 71% y Ligüi 68% del total de la abundancia), seguidas de las detritívoras (Loreto 27% y Ligüi 26%) y finalmente las carnívoras (Loreto 5% y Ligüi 3%).
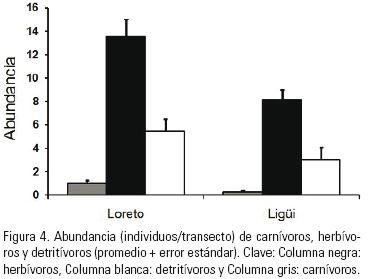
El valor promedio de abundancia de carnívoros en ambas localidades fue de 0.75 ± 0.14 ind/transecto, y hubo diferencias significativas entre Loreto y Ligüi (t2,90.3 = 3.07, p = 0.002. Fig. 4). Para los herbívoros la abundancia general fue de 11.26 ± 0.95 ind/transecto, y también se presentaron diferencias significativas (t2,941 = 3.21, p < 0.001. Fig. 4), mientras que el promedio para los detritívoros fue de 4.41 ± 0.78 ind/transecto y ambas localidades no difirieron entre sí (t2,104 = 1.56, p < 0.122. Fig. 4). Aunque, se encontró una clara diferencia en el número de grupos tróficos presentes por transecto (t21038 = 6.07, p < 0.001), el cual fue mayor en Loreto (2.19 ± 0.09) que en Ligüi (1.42 ± 0.08).
DISCUSIÓN
La información presentada en el presente estudio exhibe por vez primera las tendencias de abundancia y ocurrencia de distintos gremios tróficos de asteroideos. Según Cintra–Buenrostro (2001) y Solís–Marín et al. (2005), en el Golfo de California existen 47 especies de estrellas de mar de aguas someras (menores a 200 m), pero en este trabajo sólo fueron encontradas 9 especies (Tabla 1). La diferencia se explica porque más de la mitad de los asteroideos de este mar interior aparecen en fondos arenosos o limosos (Caso et al., 1993; Cintra–Buenrostro et al., 2005), mientras que los de zonas rocosas, algunos son endémicos del Alto Golfo y otros son específicos de la zona tropical del mismo (al sur de los 24°N; Hendrickx et al., 2005), de modo que no ocurren en Loreto. Asimismo, cabe señalar que Sagarin et al. (2008) enseñan que la riqueza y abundancia local de asteroideos y de equinodermos en general, ha disminuido sensiblemente en el golfo desde los 1940, posiblemente debido a efectos sinérgicos de menor calidad de hábitat y eventos de calentamiento oceánico (en especial El Niño y la fase cálida de la Oscilación Decadal del Pacífico).
El hallazgo que P. unifascialis fue la especie más abundante (52 % del total de individuos observados) y frecuente (88% de los transectos) en Loreto y Ligüi (Tabla 1), concuerda con lo descrito en los estudios de Morgan y Cowles (1996); Holguín–Quiñones et al. (2000) y González–Medina et al. (2006) para la Bahía de Loreto, y las observaciones y datos de Reyes–Bonilla et al. (2005) y Herrero–Pérezrul et al. (2008) para el resto del Golfo de California, por lo que se concluye que es el asteroideo dominante en aguas someras y arrecifales de este mar interior. Es posible que esta afirmación pueda extenderse al resto del Pacífico mexicano ya que Zamorano y Leyte–Morales (2005), Benítez–Vil la lobos (2008) y Ríos–Jara etal. (2008) coinciden en que P. unifascialis también es el asteroideo más numeroso en Nayarit (20°N) y Oaxaca (15°N).
Respecto al orden de abundancia de las especies en Loreto, Holguín–Quiñones etal. (2000) indicaron que la estrella de mar corona de espinas, Acanthaster planci, estaba en los tres primeros lugares. La diferencia respecto a nuestros datos (Tabla 1) puede ser evidencia de fluctuaciones en la abundancia de esta especie a escala local, pues censos recientes en Loreto (2008–2009) mostraron que A. planci ha tenido un repunte de abundancia en relación con lo observado entre 2004 y 2007 (Reyes–Bonilla etal. 2005; presente trabajo). Los cambios de abundancia de esta especie han sido observados en el Pacífico central, y oeste, en el Océano Indico, y también en América Central (Uthicke et al., 2009), por lo que será interesante hacer un seguimiento a largo plazo de la población de este importante depredador en el Golfo de California. Por último, Brusca (1980) y Sagarin et al. (2008) indican que antes de la década de los 1980s, el asteroideo más común del Golfo de California era el carnívoro Heliaster kubiniji (Xantus, 1860), el cual tuvo una caída poblacional severa causada por un patógeno a principios de los 1980 (Dungan et al., 1982). Dicha especie sigue siendo escasa no sólo en Loreto sino en todo el golfo (Reyes–Bonilla et al., 2005; Herrero–Pérezrul et al., 2008), lo que muestra que su recuperación no ha sido exitosa.
Comparando las localidades de estudio se observó que Loreto tuvo mayor riqueza, abundancia y diversidad de asteroideos que Ligüi (Fig. 2). Este resultado se explica porque la localidad de Loreto presenta gran variedad de hábitat (fondos rocosos, manglares, humedales, mantos de rodolitos; Arizpe y Riosmena–Rodríguez, 2007), los cuales ofrecen refugio y exportan carbono. Además, la parte norte de la bahía mantiene una mayor productividad oceánica debido a surgencias estacionales en la parte noreste de la Isla Carmen (Fig. 1). El aporte de energía hacia las comunidades bénticas de los arrecifes debe estar favoreciendo la biodiversidad en Loreto respecto a Ligüi.
El único índice ecológico que no cambió significativamente entre localidades fue la uniformidad de Pielou (J'), un indicador de la homogeneidad en la distribución de la abundancia de las especies de un sitio, independientemente de su composición (Krebs, 1999). El resultado indica que la abundancia relativa de individuos encontrados por especie de estrella de mar está distribuida de manera similar tanto a escala local (Loreto y Ligüi; Fig. 1) como regional (Bahía de Loreto y sus alrededores). Es importante remarcar que nuestros resultados no pudieron compararse directamente con los obtenidos por otros autores que aplicaron índices ecológicos debido principalmente a que no estimaron los valores para los asteroideos como tal, sino para invertebrados o equinodermos en general (Holguín–Quiñones et al., 2000; Benítez–Villalobos, 2001; Mille–Pagaza et al., 2002; Holguín Quiñones et al., 2008). Se sugiere que futuros trabajos traten de ofrecer información más detallada sobre cada taxón.
En el análisis de ordenación (Fig. 3) no observamos separaciones entre los sitios de trabajo, y el stress del NMDS fue relativamente alto (0.15). Estas tendencias fueron confirmadas con el ANOSIM, y demuestran que las asociaciones de asteroideos no presentan diferencias cualitativas entre las dos localidades de estudio, sólo cuantitativas. Reyes–Bonilla et al. (2005) tampoco mostraron agrupaciones claras al revisar las asociaciones de asteroideos de arrecifes del oeste del Golfo de California, aun cuando la riqueza y abundancia fueron mayores en Loreto y Bahía de Los Ángeles que en las otras localidades estudiadas. Tomando en cuenta toda esta información, se sugiere que aunque haya marcadas diferencias en la abundancia de las estrellas de mar en el golfo a escalas locales o regionales (las cuales como se dijo, deben estar señalando aspectos de disponibilidad de alimento), la homogeneidad en el índice de uniformidad indica que la abundancia relativa de las especies dominantes (siempre P. unifascialis, P. pyramidatus y P. cumingi) es muy similar a escalas locales, lo que se traduce a escalas regionales. Además, estas especies pueden habitar diferentes tipos de sustratos (rocosos, coralinos, arenosos, etc.).
El estudio de las tendencias tróficas de los asteroideos es importante dado que ellos pueden obtener alimento de muy diversas formas dependiendo la especie, y por ende el grupo representa una serie de eslabones importantes en los ecosistemas arrecifales. La clasificación empleada en este trabajo (Cintra Buenrostro et al., 2005; Tabla 1) se basó en una revisión sobre la dieta de las especies del golfo, donde se asignó cada especie a un gremio trófico con base en datos de literatura, observaciones de campo y en algunos casos, extrapolaciones de las características de especies afines. Este es el mismo método que se ha seguido desde hace ya varios años para revisar la condición de peces (Pauly et al., 1998) y moluscos (Valentine et al., 2002), por lo que se consideró que la clasificación es confiable.
Según la Figura 4, el tipo de alimentación más común en las dos localidades estudiadas fue el herbivorismo, lo cual ocurre porque las dos especies dominantes pertenecen a este gremio (Tabla 1). Este patrón puede tener su origen en la alta biomasa de algas frondosas y coralinas que caracteriza a Loreto (Arizpe & Riosmena–Rodríguez, 2008), y en general el Golfo de California (Brusca, 1980), y que son capaces de mantener cantidades importantes de peces e invertebrados. Además de esto, los números de los detritívoros y los carnívoros fueron más altos en la localidad de Loreto, quizá resultado de la alta productividad de la columna de agua en la bahía, que se manifiesta como carbono orgánico particulado que llega al fondo, y como energía transmitida dentro de la red trófica y que eventualmente alcanza el tercer nivel. Es factible que la mayor abundancia de elementos en todos los gremios tróficos sea la causa por la cual también en la localidad de Loreto se halló un mayor número de grupos tróficos por transecto que en Ligüi. Pero es interesante anotar que no obstante a ello, la proporción de la abundancia total de cada uno de los gremios se mantuvo independiente de la localidad. Esto puede indicar que la función de los asteroideos como unidad es similar en ambas localidades, y que sus números cambian solo en respuesta a la cantidad de recursos disponibles.
En conclusión, nuestro trabajo muestra que la estructura comunitaria y trófica de los asteroideos residentes en zonas rocosas de la región de estudio difiere cuantitativamente, debido a que en la localidad de Loreto existe una mayor riqueza, abundancia y diversidad de asteroideos y mayor número de gremios tróficos por transecto. Esto puede deberse a que dicha localidad presenta ambientes muy variados con fondos rocosos y abundante alimento para los asteroideos. Sin embargo, la composición de especies se mantuvo constante, debido a que las especies dominantes (P. unifascialis, P. cumingi y P. pyramidatus) pueden colonizar diferentes tipos de hábitats.
AGRADECIMIENTOS
Esta investigación se realizó con fondos del programa PRODERS otorgados a las autoridades del Parque Nacional Bahía de Loreto (Yrvin Ramírez, Director del Parque), y la información de campo se obtuvo con la colaboración de Andrés González y Arturo Ayala (UABCS). Juan José Alvarado Barrientos (Universidad de Costa Rica) revisó versiones preliminares del manuscrito. Luis Drew (UABCS) y Leonardo Vázquez Vera (COBI A. C.) ayudaron en la edición de las figuras. Agradecemos a los dos árbitros anónimos y al editor que revisaron el documento y ofrecieron comentarios que mejoraron sensiblemente la versión final.
REFERENCIAS
Arizpe, O. & R. Riosmena Rodríguez. 2007. Flora y fauna marina y costera. In: Ganster, P., O. Arizpe & A. Ivanova (Eds.). Loreto; el futuro de la primera capital de las Californias. San Diego State University Press, San Diego, pp. 31–48. [ Links ]
Bautista–Romero, J. J., H. Reyes–Bonilla, D. Lluch–Cota & S. Lluch–Cota. 1994. Aspectos generales de la fauna marina, In: Ortega–Rubio, A. & A. Castellanos–Vera (Eds.). La Isla Socorro, Reserva de la Biosfera Archipiélago de Revillagigedo. CIBNOR, La Paz, pp. 247–275. [ Links ]
Benítez–Villalobos, F. 2001. Comparación de la comunidad de equinodermos asociados a dos arrecifes, en dos localidades de las Bahías de Huatulco, Oaxaca, México. Ciencia y Mar 5 (13): 19–24. [ Links ]
Benítez–Villalobos, F. 2008. Listado taxonómico de los equinodemos (Echinodermata: Asteroidea y Echinoidea) de la costa de Oaxaca en el Pacífico sur mexicano. Revista de Biología Tropical 56 (3): 75–81. [ Links ]
Blake, D. B. 1989. Asteroidea: functional morphology, classification and phylogeny. In: Jangoux, M & J.M. Lawrence (Eds.). Echinoderm studies 3. Balkema, Amsterdam, pp. 179–223. [ Links ]
Brusca, R. C. 1980. Common intertidal invertebrates of the Gulf of California. 2nd Ed. University of Arizona Press, Tucson. 320 p. [ Links ]
Caso, M. E. 1992. Los equinodermos (Asteroideos, Ofiuroideos y Equinoideos de la Bahía de Mazatlán, Sinaloa. Publicación Especial de los Anales del Instituto de Ciencias del Mar y Limnología UNAM 11 (1): 1–214. [ Links ]
Caso, M. E. 1994. Estudio morfológico, taxonómico, ecológico y distribución geográfica de los asteroideos colectados durante las campañas oceanográficas Cortés 1, 2, 3. Publicación Especial de los Anales del Instituto de Ciencias del Mar y Limnología UNAM 12 (1): 1–111. [ Links ]
Caso, M. E., A. Laguafda–Figuefas, F. A. Solís–Marín, A. Ortega–Salas & A. de la Luz Dufán–González. 1996. Contribución al conocimiento de la ecología de las comunidades de equinodermos de la Bahía de Mazatlán, Sinaloa, México. Anales del Instituto de Ciencias del Mar y Limnología UNAM 22 (1): 101–119. [ Links ]
Chávez–Dagostino R. M., A. C. Nepote–González, P. Medina– Rosas & F. Solís–Mafín. 2000. Listado preliminar de echinoideos (sic) y asteroideos (Echinodermata: Echinoidea y Asteroidea) de las Islas Marietas, Nayarit, México. Mexicoa 2 (1): 69–72. [ Links ]
Cintra–Buenrostro, C. E. 2001. Los asteroideos (Echinodermata: Asteroidea) de aguas someras del Golfo de California, México. Oceánides 16 (1): 49–90. [ Links ]
Cintra–Buenrostro, C. E., H. Reyes–Bonilla & M. D. Herrero–Pérezrul. 2005. Oceanographic conditions and diversity of seastars (Echinodermata: Asteroidea) in the Gulf of California, Mexico. Revista de Biología Tropical 53 (3): 245–261. [ Links ]
Clarke, K.R & R. M, Warwick. 2001. Change in marine communities: an approach to statistical analysis and interpretation. 2nd edition. PRIMER–E: Plymouth Marine Lab. 171 p. [ Links ]
Dungan, M. T., T. E. Miller & D. A. Thomson. 1982. Catastrophic decline of a top carnivore in the Gulf of California rocky intertidal zone. Science 216 (4549): 989–991. [ Links ]
González–Medina, F. J., O. E. Holguín–Quiñones & G. De la Cruz–Agüero. 2006. Variación espacio temporal de algunos macroinvertebrados (Gastropoda, Bivalvia y Echinodermata) de fondos someros del Archipiélago Espíritu Santo, B.C.S., México. Ciencias Marinas 32 (1A): 33–44. [ Links ]
Gotshall, D. W. 1987. Marine animals of Baja California. Sea Challengers, Monterey. 104 p. [ Links ]
Herrero–Pérezrul, M. D., H. Reyes–Bonilla, A. González–Azcárraga, C. E. Cintfa–Buenfostfo & A. Rojas–Sierra. 2008. Equinodermos. In: G. D. Danemann y E. Ezcurra (Eds.). Bahía de Los Ángeles: recursos naturales y comunidad: línea base 2007. SEMARNAT/INE/PRONATURA Noroeste, San Diego Natural History Museum, pp. 339–357. [ Links ]
Hendrickx, M. E., R. C. Brusca & L. T. Findley. 2005. Listado y distribución de la macrofauna del Golfo de California, México. Parte 1, Invertebrados. Arizona–Sonora Desert Museum, Tucson. 429 p. [ Links ]
Hickman, C. P. 1998. A field guide to sea stars and other echinoderms of Galápagos. Sugar Spring, Lexington. 83 p. [ Links ]
Holguín–Quiñones, O., H. Wright–López & F. A. Solís–Marín. 2000. Asteroidea, Echinoidea y Holothuroidea en fondos someros de la Bahía de Loreto, Baja California Sur, México. Revista de Biología Tropical 48 (4): 749–757. [ Links ]
Holguín–Quiñones, O., E., F. J. González–Medina, F. Solís–Marín & E. Félix–Pico. 2008. Variación espacio–temporal de Scleractinia, Gorgonacea, Gastropoda, Bivalvia, Cephalopoda, Asteroidea, Echinoidea y Holothuroidea de fondos someros de Isla San José, Golfo de California. Revista de Biología Tropical 56 (3): 1189–1199. [ Links ]
Honey–Escandón, M., F. A. Solís–Marín & A. Laguarda–Figueras. 2008. Equinodermos (Echinodermata) del Pacífico mexicano. Revista de Biología Tropical 56 (3): 57–73. [ Links ]
Kerstitch, A. & H. Bertsch. 2007. Sea of Cortez marine invertebrates. 2nd. ed. Sea Challengers, Monterey. 106 p. [ Links ]
Krebs, C. 1999. Ecological methodology. 2nd. Ed, Harper & Row. New York. 620 p. [ Links ]
Maluf, L. Y. 1988. Biogeography of the central Eastern Pacific shelf echinoderms. In: Burke, R.D., P.V. Mladenov, P. Lambert & R.L. Parsley (Eds.). Echinoderm Biology. Balkema, Rotterdam, pp. 389–398. [ Links ]
McCune, B., J. B. Grace & D. L. Urban. 2002. Analysis of ecological communities. MJM Software Desig. 300 p. [ Links ]
Menge, B. A. 1982. Effects of feeding on the environment. In: Jangoux, M. & J. M. Lawrence (Eds.) Echinoderm nutrition. Balkema, Rotterdam, pp. 521–551. [ Links ]
Morgan, M. B. & D. L. Cowles. 1996. The effects of temperature on the behavior and physiology of Phataria unifascialis (Gray, 1840) (Echinodermata, Asteroidea): implications for the species distribution in the Gulf of California, Mexico. Journal of Experimental Marine Biology and Ecology 208 (1): 13–27. [ Links ]
Mille–Pagaza, S., J. Carrillo–Laguna, A. Pérez–Chi & M. E. S ánchez–Salazar. 2002. Abundancia y diversidad de los invertebrados litorales de Isla Socorro, Archipiélago Revillagigedo, México. Revista de Biología Tropical 50 (1): 97–105. [ Links ]
Paulay, G. 1996. Diversity and distribution of reef organisms. In: C. Birkeland (ed.). Life and death in coral reefs. Academic Press, San Diego, pp. 298–353. [ Links ]
Pauly, D., V. Chfistensen, vf J. Dasgaafd, R. Ffoese & F. Torres. 1998. Fishing down marine food webs. Science 279 (5352): 860–863. [ Links ]
Solís–Marín, F. A., A. Laguarda–Figueras, A. Durán–González, C. G. Ahearn & J. Torres–Vega. 2005. Equinodermos (Echinodermata) del Golfo de California, México. Revista de Biología Tropical 53 (3): 123–137. [ Links ]
Reyes–Bonilla, H. 1995. Asteroidea and Echinoidea (Echinodermata) from Isla San Benedicto, Revillagigedo Archipelago, México. Revista de Investigación Científica UABCS 6 (1): 29–38. [ Links ]
Reyes–Bonilla, H., González–Azcárraga A & A. Rojas–Sierra A. 2005. Estructura de las asociaciones de las estrellas de mar (Asteroidea) en arrecifes rocosos del Golfo de California, México. Revista de Biología Tropical 53 (3): 233–244. [ Links ]
Ríos–Jara, E., C. M. Galván–Villa & F. A. Solís–Marín. 2008. Equinodermos del Parque Nacional Isla Isabel, Nayarit, México. Revista Mexicana de Biodiversidad79 (1): 131–141. [ Links ]
Sagarin, R. D., W. F. Gilly, C. H. Baxter, N. Burnett & J. Christensen. 2008. Remembering the gulf: changes to the marine communities of Sea of Cortez since the Steinbeck and Ricketts expedition of 1940. Frontiers in Ecology and the Environment 6 (7): 372–379. [ Links ]
Uthicke, S., B. Schaffelke & M., Byrne. 2009. A boom–bust phylum? Ecological and evolutionary consequences of density variations in echinoderms. Ecological Monographs 79 (1): 3–24. [ Links ]
Valentine, J. W., K. Roy & D. Jablonski. 2002. Carnivore & non carnivore ratios in northeastern Pacific marine gastropods. Marine Ecology Progress Series 228:153–163. [ Links ]
Warwick, R. M. & K. R. Clarke. 2001. Practical measures of marine biodiversity based on relatedness of species. Oceanography and Marine Biology Annual Review 39 (1): 207–231. [ Links ]
Zamorano, P & G. E. Leyte–Morales. 2005. Cambios en la diversidad de equinodermos asociados al arrecife coralino de La Entrega, Oaxaca, México. Ciencia y Mar 9 (27):19–28. [ Links ]
Zar, J. H. 2009. Biostatisticalanalysis. 5th Ed. Prentice Hall, Upper Saddle River. 944 p. [ Links ]

