










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.18 no.2 Ciudad de México Ago. 2008
Monitoreo del crecimiento del Cocodrilo americano Crocodylus acutus, durante su primer año de vida en condiciones de cautiverio
Growth monitoring during captivity of the American Crocodile Crocodylus acutus along its first year
Juan Meraz1*,2, José Alberto Montoya Márquez1, Edith Ávila Nahon3, y Laurentino Reyes Sánchez4
1 Instituto de Recursos, Universidad del Mar, Puerto Ángel, Oaxaca, México. **E–mail: j.meraz–hernando.1@research.gla.ac.uk
2 Institue of Biomedical and Life Sciences, University of Glasgow. Graham Kerr Building, Room 517. G12 HQQ Glasgow, UK.
3 Biología Marina, Universidad del Mar, Puerto Ángel, Oaxaca, México.
4 Sociedad Cooperativa La Ventanilla. Tonameca, Oaxaca, México.
Recibido: 20 de diciembre de 2007.
Aceptado: 30 de julio de 2008.
RESUMEN
Un aspecto a considerar durante el cautiverio de crías de cocodrilo americano (Crocodylus acutus), es el relacionado con el tipo de crecimiento de los individuos. Para evaluarlo, se seleccionaron cuatro nidadas naturales en el estero La Ventanilla, Oaxaca, las cuales se mantuvieron en cautiverio a lo largo de un año. Con base en mediciones mensuales de longitud y peso se aplicaron varias ecuaciones de desarrollo, observándose el tipo de alométria, toda vez que se encontró una relación positiva entre la talla y el peso. De los modelos de crecimiento de dos parámetros empleados el que mejor se ajustó a los datos fue el lineal, mientras que de las igualdades de 3 parámetros el que presentó mejor ajuste fue el de von Bertalanffy. El incremento promedio para los organismos estudiados fue de 0.614 mm día–1, o 1.84 cm mes–1, lo cual es bajo comparado con estudios realizados en Jalisco, México. y Brasil, por lo que se considera que es importante revisar las condiciones prevalecientes durante el cautiverio.
Palabras clave: Crocodylus acutus, von Bertalanffy, La Ventanilla, crecimiento.
ABSTRACT
An aspect to consider during the captivity of the American crocodile (Crocodylus acutus), is that related to the individuals growth. In order to evaluate this issue, four natural nests from La Ventanilla, Oaxaca were selected, and kept in captivity one year. Based on monthly measurements of length and weight several growth models were applied. Allometric growth was observed due to the positive relationship between length and weight. Of the two parameter growth models used, the lineal type was the most appropriated. Of the three parameter growth models used, the von Bertalanffy type was the most adequate. Mean growth for the studied individuals was 0.614 mm day–1, or 1.84 cm month–1. This represents low values in comparison to other studies from Jalisco, Mexico and Brasil. Therefore, we consider it important to check the conditions that prevail during captivity.
Key words: Crocodylus acutus, von Bertalanffy, La Ventanilla, growth.
INTRODUCCIÓN
El cocodrilo americano Crocodylus acutus (Cuvier 1807), también conocido como cocodrilo de río, es una de las especies de cocodrilianos más estudiados (Alvarez del Toro & Sigler, 2001). A pesar de ello, en los años noventas en México se desconocía la distribución y abundancia de sus poblaciones (Méndez–de la Cruz & Casas–Andreu, 1992), existiendo numerosas localidades donde no se tenían registros sobre su presencia lo que aunado a la cacería furtiva (Thorbjarnarson, 1992) y la modificación de su hábitat, hizo de esta una especie susceptible, por lo que está sujeta a protección especial (Casas–Andreu et al., 2004). Estudios recientes para estimar sus poblaciones utilizan diferentes técnicas como: el conteo de ojos durante censos nocturnos, empleando la técnica de lampareo (Cupul–Magaña et al., 2002a; Escobedo–Galván, 2003; Andrade–Esquivel et al., 2004; Hernández–Jiménez et al., 2004; Huerta–Ortega et al., 2004), observaciones directas de ejemplares en sus asoleaderos (Méndez–de la Cruz & Casas–Andreu, 1992) y conteo de nidos (Platt & Thorbjarnarson, 2000; López–Osuna et al., 2004; Platt et al., 2004).
Un tema que ha adquirido importancia en el estudio actual de los cocodrilos, lo representa el mantenimiento de ejemplares en cautiverio (Piedra et al., 1997; Cupul–Magaña et al., 2002b) y su producción bajo estas condiciones, con la finalidad básica de repoblar las zonas donde la especie ha sido diezmada, ya sea por su cacería o por la modificación del su hábitat (Escobedo–Galván & Mejía–Vargas, 2003).
A fin de incrementar la sobrevivencia de los ejemplares en las etapas tempranas (crías y juveniles), los nidos puestos por hembras en vida libre son trasladados a encierros artificiales, sitio en el que se reproducen las condiciones naturales (Bolton, 1994); y una vez eclosionados los huevos, las crías son mantenidas en cautiverio hasta que alcanzan tallas que les permiten buscar su propio alimento y sobrevivir a depredadores tales como mapaches (Platt & Thorbjarnarson, 2000), coyotes, aves de presa (Romeu, 1998) y garzas. Entonces los ejemplares son liberados al medio natural donde se espera tengan altas tasa de sobrevivencia.
Previo a la colecta de los huevos, es importante conocer la biología reproductiva en las poblaciones silvestres (Mazotti, 1989, citado en Casas–Andreu, 2003), ya que es de estas de donde se obtienen los nidos que se habrán de trasladar a los encierros artificiales. Adicionalmente, deben conocerse los hábitos alimenticios (Piedra et al., 1997; Casas–Andreu & Barrios–Quiroz, 2003) así como el alimento natural disponible para cada población. Esto a fin de garantizar que los ejemplares liberados cuenten con alimento suficiente, máxime cuando se ha encontrado que los cocodrilos liberados presentan tasas de crecimiento menores a aquellos que permanecieron en vida libre (Arteaga, 1997).
El mantenimiento de cocodrilianos en condiciones controladas ha permitido estudiar su desarrollo, con relación al tipo de encierro, alimentación y condiciones naturales (Piedra et al., 1997; Parra–Miranda et al., 2002; de la Ossa–Velásquez, 2002; Escobedo–Galván, 2003). Con ello, se han propuesto condiciones de cautiverio adecuadas para obtener mayores tasas de sobrevivencia. Evaluar el crecimiento de los ejemplares cautivos, es una medida que puede emplearse como indicador de la calidad del manejo controlado. Por consiguiente, es esencial determinar la tasa de crecimiento de los ejemplares durante el periodo en cautiverio, así como reconocer aquellos factores que afectan el propio desarrollo de las crías, a fin de que se alcancen tallas adecuadas de liberación. Esto último es muy importante ya que en algunos lugares, como en La Ventanilla, el periodo en que se mantienen las crías en cautiverio es limitado y se espera que crezcan en óptimas condiciones para que puedan sobrevivir al reintegrarse en su hábitat.
Investigaciones recientes han incluido el análisis de tallas de las crías en cautiverio, aplicando para ello modelos de crecimiento (Cupul–Magaña, 2002). En los estudios del crecimiento, es necesario el análisis de las relaciones morfométricas, ya que permiten conocer el tipo de proporción que existe entre el crecimiento relativo de dos o más estructuras corporales de un organismo (McMahon & Tyler–Bonner, 1986), predecir la talla o peso de algún individuo, o incluso mostrar cambios en el proceso ontogénico de los cocodrilianos (Verdade, 2000). Si bien el crecimiento puede emplearse para determinar la edad de un cocodrilo (Webb et al., 1983; Verdade, 2000), también puede ser una herramienta útil para establecer el tiempo que una cría debería permanecer en cautiverio antes de ser soltada, debido a la densidad de ejemplares en los encierros (Cupul–Magaña & Hernández–Hurtado, 2002) u otras condiciones como la calidad de los encierros o del alimento suministrado.
Las condiciones de encierro difieren entre localidades, principalmente porque se emplean los recursos naturales disponibles en cada lugar. La fuente principal de agua y los materiales para construir los encierros, así como el diseño de éstos, son particulares de cada criadero, aunque el entorno en general debe ser similar y apegado a programas de manejo adecuados.
En el estero La Ventanilla se han mantenido cocodrilos en cautiverio por varios años, y los escenarios de encierro han presentado modificaciones. Evaluar la eficiencia del criadero en general permite conocer el grado de éxito obtenido, así como aporta conocimiento sobre la biología de los cocodrilos en general, y su ecología en el estado de Oaxaca. Por ello, el presente trabajo tiene como objetivo evaluar el crecimiento de las crías de cocodrilo bajo las condiciones de cautiverio prevalecientes en el estero La Ventanilla, desde su eclosión hasta el primer año de vida cuando son liberadas.
MATERIALES Y MÉTODOS
El trabajo se desarrolló durante el periodo comprendido entre los meses de mayo del 2003 y junio del 2004, en el estero La Ventanilla, Oaxaca, localizado en la parte oriental de la laguna de Tonameca. Está limitado al sur por una barra arenosa que se abre por unos días durante la época de lluvias, usualmente en mayo o junio, existiendo en el estero un ambiente preponderantemente dulceacuícola (Becerril–Morales, 2001). En ella la comunidad asentada ha establecido una UMA (Unidad de Manejo y Aprovechamiento Sustentable de la Vida Silvestre) que realiza recorridos ecoturísticos en embarcaciones de remos para observar aves y cocodrilos principalmente (Ávila–Foucat, 2002). La reintroducción al medio silvestre de crías de cocodrilo mantenidas en cautiverio ha sido una experiencia exitosa en La Ventanilla (García–Grajales et al., 2007). Dentro del estero hay asoleaderos donde se les provee de alimento (pollo y pescado), que complementa su dieta natural y consiste de animales que capturan en el estero como peces, invertebrados y las crías de aves acuáticas que anidan en las ramas de la vegetación circundante (Meraz & Ávila–Nahón, 2006).
Se identificaron 4 nidos (nidadas) en las inmediaciones del estero. La incubación se realizó en los nidos naturales (construidos por las madres en libertad) a fin de mantener el desarrollo embrionario con las variaciones naturales de temperatura de incubación, cosa que no ocurre en nidos artificiales (Parra–Miranda et al., 2002). Las crías eclosionaron los días 24, 27 y 29 de mayo, así como 4 de junio, y fueron trasladadas para su mantenimiento en tinas de fibra de vidrio de 150 X 45 cm, con una profundidad de 35 cm, durante los primeros tres meses, y trasladadas a corrales temporales (acuaterrarios) con un área de 2 m2, con el fin de mantenerlas hasta alcanzar el año de vida, cuando fueron liberadas para continuar su desarrollo en vida libre. El traslado de las crías a corrales de crianza forma parte de las actividades de la UMA encaminadas a incrementar la sobrevivencia de los ejemplares, manteniéndose el siguiente régimen de alimentación: desde la eclosión hasta los 3 meses de edad se les suministra hígado de res 30 g individuo–1 cada tercer día, conservándolos en las tinas de fibra de vidrio dentro de una cabaña de madera, provista de un foco que sirve para atraer insectos que complementan la dieta. La temperatura promedio dentro de la cabaña fue de 25–30 °C durante el día y de unos 18–27 °C durante la noche a lo largo del año. Posteriormente y hasta su liberación, se les proporcionó pollo (incluyendo pechuga) ad libitum, alcanzando aproximadamente los 32 g individuo–1. El método de suministrar el alimento es dándoles aproximadamente 4 Kg por nidada por la tarde y de consumirse todo hasta la mañana siguiente, se les ofrece una ración menor y así sucesivamente hasta que se observa que no se consume todo el pollo (retirando el excedente del acuaterrario). Este procedimiento se repitió cada tercer día. La temperatura promedio en el acuaterrario osciló entre 27–35 °C a la sombra durante el día, y entre15–25 °C durante la noche a lo largo del año. Por su parte, el agua que se obtiene del estero presenta una temperatura promedio de 27 °C.
A fin de no interferir en los trabajos realizados en la UMA, así como evaluar las condiciones de cautiverio prevalecientes, no se modificaron las condiciones de cautiverio o alimentación.
Durante el periodo en que se encontraron en cautiverio se registraron datos morfológicos y merísticos mensuales, consistentes en la longitud total (desde la punta del hocico hasta el final de la cola) y el peso.
A fin de determinar el tipo de relación morfométrica existente entre la longitud total y el peso, se utilizó el análisis de regresión lineal con los valores transformados a logaritmo natural. La pendiente determinó la razón de la proporción de cambio del peso en gramos dividida por la razón de la talla en centímetros (McMahon & Tyler–Bonner, 1986). Este análisis incluyó las tallas y los pesos registrados mensualmente para las cuatro nidadas.
La ecuación que describe la relación talla–peso es:
Peso = a x (Talla)b
Donde: b = Pendiente o coeficiente de alometría
a = Valor de la ordenada (cuando la talla =0)
Para corroborar los supuestos de linealidad, normalidad, homocedasticidad e independencia de los datos del análisis de regresión, se utilizó el análisis de residuales (Mendenhall & Sincich, 1996), para detectar posibles puntos aberrantes, los residuales se estandarizaron, siendo aberrantes aquellos cuyos valores fueron mayores a 2 y menores a –2 (Zar, 1996).
Para conocer el tipo de crecimiento relativo (alometría) que presentaron los cocodrilos durante su primer año de vida, se utilizó la prueba t de student con un nivel de confianza del 95% (Zar, 1996):
t = b1 – 3 /Sb1
Donde: t = Estadístico de prueba
b1 = Pendiente de la relación morfométrica
Sb1 = Error estándar de la pendiente.
De obtenerse una pendiente igual a tres entonces la relación sería isométrica, esto significa que las proporciones del organismo se mantienen constantes a través de su crecimiento (McMahon & Tyler–Bonner, 1986).
Se probaron diferentes modelos, a fin de describir el crecimiento del cocodrilo americano durante su primer año de vida en cautiverio, empleando las mediciones mensuales de los organismos para las cuatro nidadas, las que se tomaron como réplicas. Para evaluar el crecimiento se usó la longitud y peso promedio por nidada.
Las ecuaciones probadas se dividieron en aquellas con 2 y 3 parámetros. Entre los primeros se contrastaron los del tipo lineal, logarítmico, potencial y exponencial; haciendo las estimaciones por medio del método de mínimos cuadrados. Entre los segundos se aplicaron, el de von Bertalanffy (vB), el cual permite calcular el crecimiento en longitud y peso a través del tiempo (von Bertalanffy, 1938), y el de Gompertz (G) (Beverton & Holt, 1957).
El modelo vB para la longitud se expresa:
Lt = L∞ (1 – ek(t–to))
Donde: Lt = Longitud al mes t
L∞ = Longitud máxima que los organismos pueden alcanzar al año de edad
k = Coeficiente de conversión catabólica
t0 = Edad hipotética que el cocodrilo debiera tener para que su longitud fuese cero.
Para estimar los parámetros adecuados se utilizaron los métodos propuestos por Ford Walford Gulland–Holt y Levenberg–Marquardt. Para valorar qué método se ajustó mejor a los valores observados, se calculó el coeficiente de determinación y la suma de las diferencias al cuadrado (SD2), considerándose como mejor aquél con el valor más alto del coeficiente de determinación y el valor más bajo de la suma de las diferencias al cuadrado.
El modelo de crecimiento en longitud de G se expresa (Ricker, 1975):
Lt = L0e G (1–e –gt )
Donde Lt= Longitud al mes t
L0= Longitud al tiempo t= 0, en este caso es la talla al cual emergen del huevo
G= Tasa instantánea de crecimiento en t0 y donde L= L0,
g= Tasa instantánea de decremento de G.
La curva de Gompertz se usa para describir curvas sigmoides mientras que la de von Bertalanffy es una curva monotónica desacelerada.
Se calcularon las tasas instantáneas de crecimiento absoluto (TCA) por mes que se expresa en dL/dt, y por día (TCDA), así como la de crecimiento promedio mensual. Dado que la TCA depende del tamaño que ha alcanzado el organismo, una estimación más apropiada es la tasa instantánea de crecimiento específico: gL= dL / Ldt (Wootton, 1991).
Sobre un periodo de tiempo definido, la gL es calculada como:
GL = LogeP2 – LogeP1 / t2 – t1
y frecuentemente se expresa en porcentaje por unidad de tiempo: GL= 100 gL .
Para convertir las tasas por mes a día, y viceversa, se dividieron o multiplicaron por 30, según el caso, los valores obtenidos.
Los valores reportados en el presente trabajo son los promedios ± desviación estándar (S).
RESULTADOS
De las 4 nidadas colectadas eclosionaron 35, 34, 29 y 32 crías en cada una, sobreviviendo hasta el final del periodo 25, 21, 25, y 24 cocodrilos respectivamente. En las tinas de fibra de vidrio se mantuvieron, durante los primeros tres meses, a densidades de 45 ind m–2, 43 ind m–2, 37 ind m–2 y 41 ind m–2 respectivamente. En los corrales, las densidades fueron aproximadamente de 13 ind m–2, 11 ind m–2, 13 ind m–2 y 12 ind m–2 hasta la liberación. Con estos datos se calculó el éxito de sobrevivencia por nidada, para el periodo de un año, que fueron: 0.71 (24 de mayo), 0.61 (27 de mayo), 0.86 (29 de mayo) y 0.75 (4 de junio). Las longitudes promedio mensuales variaron de manera similar para las 4 nidadas (F33,132= 1.020; p > 0.444), a lo largo del año de estudio, teniendo la nidada del 4 de junio las tallas promedio mayores (Fig. 1).
La talla promedio de nacimiento, considerando las 4 nidadas, fue de 29.98 (±1.49 cm), con un mínimo de 27.00 y un máximo de 33.5 cm.
Se encontró una relación positiva significativa entre la longitud total y el peso de los organismos (p < 0.0001, R= 0.97, R2ajs= 0.946), indicando que los mayores pesos se encontraron en individuos con tallas mayores (Fig. 2), lo que se expresa en la ecuación:
Peso = 0.0016 × Talla3.21
El tipo de crecimiento relativo entre estas dos variables fue alométrico positivo (t = 9.600, gl = 1270, p <0.0001) de acuerdo al valor de la pendiente de la relación (3.21 > 3). Esto implica que los organismos crecieron más en peso que en longitud dentro del intervalo de tallas considerado.
Se calcularon las tallas medias de cada mes, así como su error estándar (Fig. 3), con lo cual se obtuvo la relación edad (en meses) y longitud promedio de los organismos estudiados (Tabla 1).
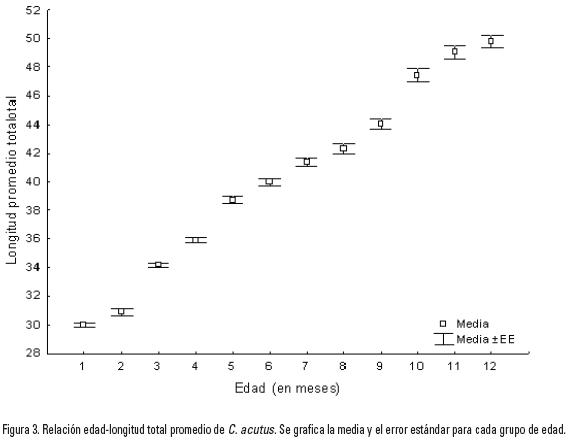
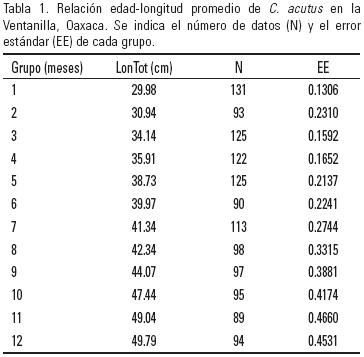
Con base en los datos de los grupos mensuales y la longitud promedio se calcularon los diferentes modelos planteados para describir el crecimiento de los cocodrilos en su primer año de vida, y con esto estimar la tasa de crecimiento de los mismos. De aquéllos con 2 parámetros el que mejor se ajustó a los datos fue el modelo lineal (R2= 0.989, SD2= 5.42) (Tabla 2).
Con respecto a los modelos con 3 parámetros usados, el que presentó el mejor ajuste fue el de vB (R2= 0.99, SD2= 4.75). El método que mejor se ajustó a los datos observados fue el de Levenberg–Marquard (Tabla 3).
La curva calculada para el crecimiento en cautiverio de C. acutus, durante su primer año de vida en el estero La Ventanilla, tuvo los siguientes parámetros: L∞ = 115.20, K = 0.0246, t0 = –11.13, siendo la ecuación de vB para crecimiento en longitud:
Lt = 115 x (1–e–0.0246(t+11.13))
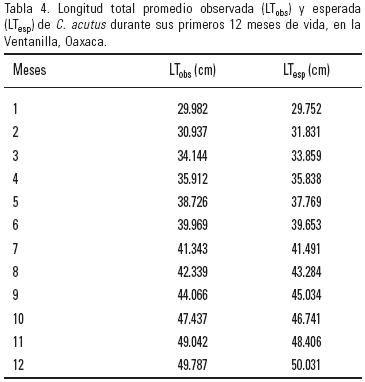
Empleando los valores obtenidos del modelo de vB se calcularon las longitudes teóricas de la muestra estudiada para el primer año de vida (Tabla 4). Con estas longitudes se generó la curva de crecimiento (Fig. 4). El crecimiento promedio para los organismos estudiados fue de 0.614 mm día–1, o 1.84 cm mes–1, lo cual es bajo comparado con los 4.15 cm reportados por De la Ossa–Velásquez y Sampedro–Marín (2002), o lo citado por Álvarez del Toro y Sigler (2001) además de otros estudios (Tabla 5).
En lo que respecta al crecimiento instantáneo los organismos presentaron una tasa de 0.047 por mes (Fig. 5), con una tendencia a disminuir con el tiempo.
Es importante mencionar que, por errores de manejo, a partir del tercer y cuarto mes se anexaron dos organismos de vida libre a la primera nidada; como no se pudo determinar con exactitud cuáles eran, se mantuvieron por el resto del periodo. Lo mismo ocurrió en la nidada 3 durante el séptimo mes (cuando se anexó un organismo) y en la cuarta nidada durante el mes quinto y séptimo (también un organismo para cada caso).
Por otro lado, durante el segundo mes en la segunda nidada se detectó un organismo con crecimiento anormal (presentando una joroba notable).
En el octavo mes se escaparon 10 organismos de la primera nidada y adicionalmente se detectaron organismos que presentaban hongos durante el tercer mes en la cuarta nidada por lo que se les cambio la fuente de agua.
Para corroborar que la adición de organismos de vida libre, muerte de organismos, organismos con deformaciones y enfermedades durante el presente trabajo no afectaron la estimación del crecimiento, se hizo una comparación entre el crecimiento de las cuatro nidadas, mediante un análisis de medidas repetidas a un nivel de confianza del 95 % (Tabla 6). No se encontraron diferencias significativas entre las nidadas durante los meses de estudio (F33,132=1.02; p=0.444), lo que indica que las situaciones antes descritas no afectaron la estimación de los modelos.
DISCUSIÓN
La talla promedio de nacimiento calculada, fue mayor a la reportada por Cupul–Magaña et al. (2004) en Jalisco (26.32 ± 0.50 cm) dichos autores sólo midieron una nidada, lo que pudo resultar en la diferencia entre ambos trabajos.
El tipo de crecimiento relativo encontrado coincide con lo descrito para cocodrilianos, dado que se ha reportado crecimiento alométrico para el caimán de nariz ancha (Caiman latirostris, Daudin 1801) en Brasil (Verdade, 2000). Los cambios alométricos son compensaciones de los organismos frente a necesidades dependientes del área superficial (difusión, fricción y pérdida del calor) (McMahon & Tyler–Bonner, 1986), respondiendo a cambios fisiológicos durante su desarrollo, mismos que están relacionados tanto con variables ambientales como con cambios en la ración alimenticia y específicamente en su dieta (McMahon & Tyler–Bonner, 1986; Wotton, 1991). Estos cambios en tamaño pueden darse por simples bases genéticas y ontogenéticas (Gould, 1971) y, para el caso de organismos adultos, pueden estar relacionados con las épocas reproductivas.
En cuanto al ajuste de los modelos probados, los que mejor se ajustaron fueron aquellos con mayor número de parámetros (von Bertalanffy y Gompertz). A pesar de que los modelos de dos parámetros fueron significativamente satisfactorios (p < 0.05) y explicaron un porcentaje alto de varianza (todos alrededor del 90%) tuvieron un menor ajuste, esto se debe por lo menos a dos razones: a) matemáticamente los que tienen mayor número de parámetros se ajustan mejor (Reyes–Victoria, 1996), b) éstos violan los supuestos de independencia de los residuales y la no correlación entre los parámetros, esto puede ocasionar inconsistencias en la estimación de parámetros resultando en valores erróneos. Estas irregularidades son relevantes principalmente en la estimación de la tasa de crecimiento. En el caso de vB y Gompertz dichos supuestos no son aplicables. Es importante mencionar que el modelo de Gompertz describe adecuadamente el crecimiento en peso o talla en los primeros años de vida, contrario a lo encontrado en el presente trabajo, probablemente debido a que se usaron datos de talla y no de peso, sin embargo diversos autores concuerdan que el modelo de vB describe de mejor forma el crecimiento de los organismos, no sólo de vertebrados sino también de invertebrados (Beverton & Holt, 1957; Ricker, 1975; Frazer & Ehrhart, 1985; Moreau, 1987, Valiela, 1995).
Para escoger un modelo biológico debe buscarse que sea sencillo (principio de parsimonia), que describa de forma razonable las características del fenómeno estudiado, que el número de supuestos sea el menor (Lackey & Hubert, 1977), que los parámetros presentes sean confirmables por experimentos independientes y que de la teoría puedan derivarse predicciones de hechos aún no observados (von Bertalanffy, 1938). Esto implica que los parámetros tengan significado biológico. El modelo de vB cumple con estas características, ya que se ha comprobado la función biológica de sus parámetros (Beverton & Holt, 1954, Valiela, 1995).
También es importante que las constantes del modelo se calculen fácilmente; en el caso de vB existen diversos métodos de estimación, el que mejor se ajustó fue el de Levenberg–Marquardt (mínimos cuadrados no lineales) el cual produce, en general, mejores ajustes para ecuaciones no lineales (Statsoft, 2001), lo cual es el caso.
La curva de crecimiento estimada indica mayor crecimiento en los dos primeros meses de vida de los cocodrilos, manteniéndose más o menos constante para el resto del periodo de estudio.
La longitud máxima (L∞) para este caso parece no representar un valor práctico, en teoría indica el tamaño máximo que pueden alcanzar los organismos de una población. En este sentido sería entonces la longitud máxima que podría alcanzar un cocodrilo en su primer año de vida (115 cm). Sin embargo, cabe destacar que vB sobreestima los valores de L∞, principalmente cuando se hacen determinaciones durante los primeros años de vida o cuando se trata de organismos de crecimiento lento (Moreau, 1987).
K es una tasa constante con unidades de tiempo recíproco, en este caso mes–1. von Bertalanffy (1938) comprobó que esta es igual a la tasa catabólica en los organismos, y se refiere a la tasa de degradación de las substancias del cuerpo, principalmente proteínas. Se ha visto que si la tasa de catabolismo es baja entonces el crecimiento es más lento y los organismos tienen una longevidad mayor. Dado que en la presente investigación el valor de K es pequeño, se puede inferir que los cocodrilos en La Ventanilla crecerían lentamente (y tendrían una alta longevidad) de mantenerse en las condiciones de cautiverio.
La baja tasa de crecimiento observada, en comparación con las reportadas para la especie, podría deberse a la presencia de hongos en algunos individuos, la deformidad de otros y, sobretodo, las condiciones de cautiverio; sin embargo, no se contó con datos que sustentaran esta idea. No obstante, se ha considerado que las condiciones inadecuadas (Cupul–Magaña et al., 2002a) son un factor importante que debe ser evaluado en estudios sobre crecimiento del cocodrilo de río.
Es substancial considerar que algunos factores como la densidad de ejemplares por encierro y la temperatura ambiental, pudieron incidir de alguna manera en el bajo crecimiento de los cocodrilos estudiados. En el primer caso, se considera que el desarrollo puede verse afectado por la densidad poblacional y la presencia de enfermedades (Moreau, 1987). La densidad de ejemplares en tinas o corrales es un factor importante para el incremento de los organismos, así como el crecimiento diferencial, ya que las crías que crecen con mayor rapidez pueden evitar que las pequeñas se alimenten adecuadamente (Bolton, 1994). Sin embargo, existen ejemplos de hacinamiento extremo donde se han podido criar cocodrilos (Bolton, 1994). Por su parte, la inhibición del crecimiento de las crías pequeñas puede compensarse con el intercambio de ejemplares, procurando que los encierros contengan individuos de tallas similares en lugar de mantenerlos en sus nidadas originales.
Por otra parte, dada la condición ectotérmica de la especie, la temperatura ambiental es un factor clave en el desarrollo y crecimiento de los organismos, Diferencias significativas se han documentado entre los crecimientos a temperaturas con y sin control (de la Ossa–Velásquez, 2002). De igual manera, se ha establecido la importancia de la densidad de manejo en organismos en cautiverio, así como su relación con el alimento (de la Ossa–Velásquez & Sampedro–Marín, 2001). No existen protocolos estrictos de alimentación en La Ventanilla, ya que el alimento se suministró en pequeñas cantidades hasta que los ejemplares dejaron de consumirlo. Con este método se garantizó que los cocodrilos reciben la cantidad deseada de alimento (Bolton, 1994), pero no permitió saber con certeza la cantidad de alimento consumida por ejemplar.
El crecimiento observado en la nidada de junio, notablemente superior al de las nidadas anteriores, las cuales presentaron valores muy semejantes, podría estar relacionado con la experiencia de la madre, toda vez que las hembras primerizas pueden tender a anidar temprano.
Si bien la introducción de nuevos ejemplares en los encierros no afectó los resultados obtenidos, pudo haber modificado la conducta de algunos organismos al incrementar la competencia por comida y espacio. Esto se pudo haber evitado marcando a los ejemplares. Para ello existe un método estandarizado de marcaje, consistente en el corte de las crestas caudales, que no fue empleado debido a que el marcaje definitivo se hace hasta el momento de la liberación. A pesar de que se colocaron marcas de colores en los ejemplares, muchas de ellas se perdieron a lo largo del periodo de estudio.
En el caso del organismo deforme, siendo el de menor tamaño de su nidada durante los primeros meses, presentó a partir del quinto mes una talla similar a la del resto de la nidada (su talla se encontraba dentro de los intervalos de confianza (95%) de los meses indicados).
Se concluye que el crecimiento de las crías de cocodrilo americano, bajo las condiciones de cautiverio prevalecientes en el estero La Ventanilla, es lento en comparación con lo reportado para Colombia (de la Ossa–Velásquez y Sampedro–Marín, 2001; de la Ossa–Velásquez, 2002), Puerto Vallarta, Jalisco, México (Cupul–Magaña, 2002), Quintana–Roo, México (Merediz–Alonso, 1999, citado en Cupul–Magaña, 2002), y Panamá (Thorbjarnarson, 1997 en Cupul–Magaña, 2002). Ello puede deberse a varias razones, como las altas densidades durante los primeros meses de vida, o la falta de adición de vitaminas en el alimento suministrado. No obstante, la descripción detallada de las condiciones de cautiverio habrá de ser explicada en trabajos posteriores.
Sin embargo estas condiciones han cambiado últimamente en el lugar, con la construcción de cuatro nuevos acuaterrarios (con un área aproximada de 10 m2 cada uno), el constante cambio de agua en éstos (hasta cada 3–4 días) y la aplicación de mejores medidas de vigilancia, lo cual habrá de repercutir no sólo en el mejor crecimiento de los organismos, sino también en el incremento en la sobrevivencia en esta etapa de vida.
Por último, conviene señalar que el modelo de crecimiento obtenido en el presente estudio, describe sólo el crecimiento en el primer año de vida, cuando aún el organismo no destina energía a la reproducción, por lo que este modelo no puede ni debe ser usado para modelar el crecimiento de juveniles y mucho menos de adultos.
AGRADECIMIENTOS
Este trabajo no hubiera sido posible sin el apoyo de la Sociedad Cooperativa La Ventanilla, especialmente del señor Perfecto Escamilla. El equipo y materiales empleados para el manejo y medición de los organismos fueron adquiridos con el apoyo del PIFI–FOMES. Finalmente los autores agradecen a Atanasio Martínez, Alberto Escamilla y Betzabeth González su colaboración en la medición de los organismos, a Beny Gueck la revisión del abstract, así como a Jesús García–Grajales por la valiosa información proporcionada. Este trabajo se enriqueció notablemente con los comentarios y observaciones de dos revisores anónimos.
REFERENCIAS
Andrade–Esquivel, A., H. Hernández–Hurtado & O. Barragán Cuencas. 2004. Diagnóstico poblacional del cocodrilo de río (Crocodylus acutus) en el área natural protegida Estero El Salado, Puerto Vallarta, Jalisco. Memorias de la 6ª Reunión COMACROM. Centro Universitario de la Costa, Universidad de Guadalajara, pp. 24. [ Links ]
Alvarez del Toro, M. & L. Sigler. 2001. Los Crocodylia de México. IMENAR – PROFEPA, México. 134 p. [ Links ]
Arteaga, A. 1997. Crecimiento, sobrevivencias y uso de hábitat de Crocodylus acutus introducidos en el embalse de Tacarigua, Edo. Falcón, Venezuela. Crocodile Specialist Group Newsletter, 16(2):16–17. [ Links ]
Avila–Foucat, S.V. 2002. Community–based ecotourism management moving towards sustainability, in Ventanilla, Oaxaca, Mexico. Ocean and Coastal Management, 45:511–529. [ Links ]
Becerril–Morales, F. 2001. Algunos aspectos sobre la ecología y conservación en el Estero La Ventanilla, Oaxaca, México. Ciencia y Mar, 15(5):37–45. [ Links ]
Beverton, R.J. & S.J. Holt. 1957. On the Dynamics of Exploited Fish Populations. Facsimile reprinted, 1993. Chapman y Hall. London UK. 533 p. [ Links ]
Bolton, M. 1994. La explotación del cocodrilo en cautividad. Guía FAO conservación 22. Roma. 80 p. [ Links ]
Casas–Andreu, G. 2003. Ecología de la anidación de Crocodylusacutus (Reptilia: Crocodylidae) en la desembocadura del río Cuitzmala, Jalisco, México. Acta Zoológica Mexicana, 89:111–128. [ Links ]
Casas–Andreu, G. & G. Barrios–Quiroz. 2003. Hábitos alimenticios de Crocodylus acutus (Reptilia: Crocodylidae) determinados por el análisis de sus excretas en la costa de Jalisco, México. Anales del Instituto de Biología de la UNAM, Serie Zoológica. 74(1):35–42. [ Links ]
Casas–Andreu, G., F.R. Méndez de la Cruz & X. Aguilar–Miguel. 2004. Anfibios y Reptiles. In A.J. García–Mendoza, M.J. Ordóñez & M. Briones–Salas (eds.), Biodiversidad de Oaxaca. Instituto de Biología, UNAM – Fondo Oaxaqueño para la Conservación de la Naturaleza – World Wildlife Found, México. pp 375–390. [ Links ]
Cupul–Magaña, F. G. 2002. Edad del cocodrilo de río Crocodylus acutus, usando el modelo de Von Bertalanffy. Boletín de la Sociedad Herpetológica Mexicana. 10(2):47–50. [ Links ]
Cupul–Magaña, F. G. & H. Hernández–Hurtado. 2002. Nota sobre el crecimiento de Crocodylus acutus en cautiverio. Revista Biomédica. 13(1):69–71. [ Links ]
Cupul–Magaña, F. G., A. Rubio–Delgado & A. Reyes–Juárez. 2002a. Nota sobre el crecimiento en cautiverio y libertad de crías de Crocodylus acutus en Puerto Vallarta, Jalisco. Ciencia y Mar, 17(VI):45–50. [ Links ]
Cupul–Magaña, F. G., A. Rubio–Delgado, A. Reyes–Juárez & H. Hernández–Hurtado. 2002b. Sondeo poblacional de Crocodylus acutus en el estero Boca Negra, Jalisco. Ciencia y Mar, 16(VI):43–44. [ Links ]
Cupul–Magaña, F. G., A. Niz–Villaseñor, A. Reyes–Juárez & A. Rubio–Delgado. 2004. Historia natural del cocodrilo americano (Crocodylus acutus) en el estero Boca Negra, Jalisco, México: anidación y crecimiento de neonatos. Ciencia y Mar, 23(VII):31–42. [ Links ]
De la Ossa–Velásquez, J. L. 2002. Efecto de la temperatura de manejo sobre el crecimiento de Crocodylus acutus (Crocodylia:Crodylidae). Revista Biología, 16(1):8–13. [ Links ]
De la Ossa–Velásquez, J. L. & A. Sampedro–Marín. 2001. Densidad de manejo y alimentación adecuados para la cría en cautiverio de Crocodylus acutus (Crocodylia:Crocodylidae). Revista Biología, 15 (2):105–110. [ Links ]
Escobedo–Galván, A.H. 2003. Periodos de actividad y efecto de las variables ambientales en cocodrilos (Crocodylus acutus Cuvier 1807): evaluando los métodos de determinación de la fracción visible. Ecología Aplicada 2(1):136–140. [ Links ]
Escobedo–Galván, A.H. & F. Mejía–Vargas. 2003. El "cocodrilo de tumbes" (Crocodylus acutus Cuvier 1807): estudio preliminar de su estado actual en el norte de Perú. Ecología Aplicada 2(1):133–135. [ Links ]
Frazer, N.B. & L.N. Ehrhart. 1985. Preliminary growth models for green, Chelonia mydas and loggerhead, Caretta caretta, turtles in the wild. Copeia 1: 5–79 [ Links ]
García–Grajales, J., G. Aguirre–León & A. Contreras–Hernández. 2007. Tamaño y estructura poblacional de Crocodylus acutus (Cuvier 1807)(Reptilia:Crocodylidae) en el estero La Ventanilla, Oaxaca, México. Acta Zoológica Mexicana, 23(1):53–71. [ Links ]
Gould, S.J. 1971. Geometric similarity in allometric growth: a contribution to the problem of scaling in the evolution of size. The American Naturalist 942(105):113–136. [ Links ]
Hernández–Jiménez, L., M. Balcázar–Lara, M. Rivera–Rodríguez & H. Hernández–Hurtado. 2004. Censo poblacional del cocodrilo de río (Crocodylus acutus Cuvier 1807) en el cuarto vaso de la Laguna de Cuyutlán, Armería, Colima, México, en dos meses de lluvias y dos de secas, 2003. Memorias de la 6ª Reunión COMACROM. Centro Universitario de la Costa, Universidad de Guadalajara, pp 28. [ Links ]
Huerta–Ortega, S., P. Ponce–Campos & J. Perran–Ross. 2004. Actualización del estudio de poblaciones del caimán (Crocodylus acutus) en Jalisco. Memorias de la 6ª Reunión COMACROM. Centro Universitario de la Costa, Universidad de Guadalajara, pp. 38 [ Links ]
Lackey, R.T. & W. A. Hubert. 1977. Analysis of exploited fish populations. Sea Grant. Virginia Polytechnic Institute and State University. Blacksburg, Virginia. USA. 172 p [ Links ]
López–Osuna, L., F. León–Ojeda & P. L. Arredondo–Ramos. 2004. Ecología poblacional de Crocodylus acutus en la laguna de Chiricahueto, Sinaloa. Memorias de la 6ª Reunión COMACROM. Centro Universitario de la Costa, Universidad de Guadalajara, pp. 33. [ Links ]
McMahon, T. A. & J. T. Tyler–Bonner. 1986. Tamaño y vida. Biblioteca Scientific American. Prensa Científica Editorial Labor, Barcelona España. 255 p. [ Links ]
Mendenhall, W. &T. Sincich. 1996. A second course in statistics, Regression Analysis. Fifth edition. Prentice–Hall, Inc. Simon & Shuster. A Viacom Company Upper Saddle River, New Yersey. USA. 899 p. [ Links ]
Méndez–De la Cruz, F. R. & G. Casas–Andreu. 1992. Status y distribución de Crocodylus acutus en la costa de Jalisco, México. Anales del Instituto de Biología de la UNAM, Serie Zoológica, 63(1):125–133. [ Links ]
Meraz, J. & E. Ávila–Nahón. 2006. Observations on young captive American Crocodiles (Crocodylus acutus) at La Ventanilla, Oaxaca, Mexico. Crocodile Specialist Group Newsletter, 25(2):4–5. [ Links ]
Moreau, J. 1987. Mathematical and biological expression of growth in fishes: Recent trends and further developments. In R.C. Summerfelt & G.E. Hall (eds.), Age and Growth of Fish. Iowa State University Press/Ames. USA. p. 81–113. [ Links ]
Parra–Miranda, M., G.V. de Moraes, E. Nunes–Martins, L.C. Pinto–Maia & O. Rus–Barbosa. 2002. Thermic variation in incubation and development of pantanal caiman (Caiman crocodilus yacare) (Daudin, 1802) kept in metabolic box. Brazilian Archives of Biology and Technology, 3(45):333–342. [ Links ]
Piedra, L., J.R. Bolaños & J. Sánchez. 1997. Evaluación del crecimiento de neonatos de Crocodylus acutus (Crocodylia: Crocodylidae) en cautiverio. Revista de Biología Tropical, 45(1): 289–293. [ Links ]
Platt, S.G. & J.B. Thorbjarnarson. 2000. Nesting ecology of the American Crocodile in the coastal zone of Belize. Copeia, 3:869–873. [ Links ]
Platt, S.G., T.R. Rainwater & S. Nichols. 2004. A recent population assessment of the American crocodile (Crocodylus acutus) in Turneffe Atoll, Belize. Herpetological Bulletin, 89:26–32. [ Links ]
Reyes–Victoria, J. 1996. Cálculo diferencial para las ciencias naturales. Edit. Trillas. México. 314 p. [ Links ]
Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish populations. Bulletin of the Fisheries Research Board of Canada. 191: 382 p. [ Links ]
Romeu, E. 1998. Cocodrilos mexicanos. Biodiversitas, 17(4):1–8. [ Links ]
Statsoft, 1999. Electronic Statistics Textbook. Tulsa, OK: StatSoft. WEB: http://www.statsoft.com/textbook/stathome.html. [ Links ]
Thorbjarnarson, J.B. 1992. Crocodiles an action plan for their conservation. UICN, Switzerland. 136 p. [ Links ]
Valiela, I. 1995. Marine Ecological Processes. Springer, New York. Segunda Edición. 686 p. [ Links ]
Verdade, L.M. 2000. Regression equations between body and head measurements in the broad–snouted caiman (Caiman latirostris). Revista Brasileira de Biología, 60(3):469–482. [ Links ]
Von Bertalanffy, L. 1938. A quantitative theory of organic growth (inquiries on growths laws. II). Human Biology 10(2):181–213. [ Links ]
Webb, G.J.W., R. Buckworth & S.C. Manolis. 1983. Crocodylus johns–toni in the Mc Kinaly River area, N.T.III. Growth, movement and the population age structure. Australian Wildlife Research. 10:383–401. [ Links ]
Wootton, R.J. 1991. Ecology of Teleost Fishes. Chapman & Hall. Fish and Fisheries Series, USA, 404 p. [ Links ]
Zar, J.H. 1996. Biostatistical Analysis. 3ª ed. Prentice Hall, USA, 662 p. [ Links ]

