










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.17 no.3 Ciudad de México Dez. 2007
Artículos
Morfometría y distribución de especies del género Ornithocercus (Dinophysiales: Dinophyta) del Pacífico Mexicano
Morphometry and distribution of species of the genus Ornithocercus (Dinophysiales: Dinophyta) from the Mexican Pacific
Adriana Hernández Rosas1, María Esther Meave del Castillo1, María Eugenia Zamudio-Resendiz1 y Manuel Castillo Rivera2
1 Lab. Fitoplancton Marino y Salobre. Dpto. de Hidrobiología, Universidad Autónoma Metropolitana Iztapalapa. Av. San Rafael Atlixco 186. Col. Vicentina. C.P. 09340. México, D.F. mem@xanum.uam.mx
2 Lab. Peces. Dpto. de Biología, Universidad Autónoma Metropolitana Iztapalapa.
Recibido: 8 de enero de 2007
Aceptado: 14 de septiembre de 2007
Resumen
Se describe, ilustra e indica la distribución de nueve especies de dinoflagelados del género Ornithocercus del Pacifico Mexicano (PM), dos de ellas nuevos registros. Se hace un estudio morfométrico de 7 especies de Ornithocercus (O. bilobatus, O. magnificus, O. heteroporus, O. galea, O. quadratus, O. steini y O. thumii), evaluándose 90 organismos colectados en el Pacifico Tropical Mexicano en los que se midieron 29 caracteres. Debido a la superposición morfológica entre O. steini-O. thumii, se enfatizó la evaluación en dichos taxa. Los datos se analizaron con técnicas estadísticas multivariadas: Análisis de Conglomerados (AC) y de Discriminantes (AD). El AC mostró la formación de dos grandes grupos, el primero con O. thumii, O. steini y O. quadratus (en subgrupos diferenciados) y el segundo con O. magnificus, O. heteroporus y O. galea. El AD mostró diferencias significativas entre las especies (hasta la cuarta función con λ Wilks < 0.162, p < 0.001), con una clasificación acertada del 97% y una varianza acumulada del 98.1%. Las variables de mayor importancia para la separación de los taxa fueron el largo total, el ancho del cíngulum y la longitud del radio R3, encontrándose en el conjunto de los datos diferencias significativas entre el área de la epiteca y el ancho a nivel del cíngulum. Se concluye que O. galea es suficientemente distinta de O. quadratus apoyándose así la teoría de que se trata de especies distintas y no de variedades. A pesar de la superposición entre O. steini y O. thumii, se encontraron caracteres merísticos para diferenciarlas.
Palabras clave: Morfometría, dinoflagelados, Ornithocercus, Pacífico Mexicano.
Abstract
It is described, illustrated and indicated the distribution of nine taxa of Ornithocercus; two of them being new records for the Mexican Pacific. The results of morphometric analyses over seven Ornithocercus species (O. bilobatus, O. magnificus, O. heteroporus, O. galea, O. quadratus, O. steini and. O. thumii) are presented. We evaluated 29 variables on 90 organisms of different species collected in the Mexican Tropical Pacific coasts. Due to the overlapping between O. steini-O. thumii, special analyses were done for this complex. Data were analyzed with multivariate statistical tools, Cluster Analysis (CA) and Discriminant Analysis (DA). CA resulted in two main and well differentiated groups, one of them with O. thumii, O. steini and O. quadratus (as differentiated subgroups), a second one with O. magnificus, O. heteroporus and O. galea. The DA showed significant differences up to fourth function (λ Wilks<0.162, p<0.0005) with 97.7% of cases correctly classified and accumulated variance of 98.1%. The most important variables for segregation were the total length, the cingulum width and the rib R3. Significant differences were found among epitecal area and cingulum width. We concluded that O. galea is different enough from O. quadratus so they are not varieties of a single species. Despite of the overlapping between O. steini and O. thumii we found meristic characters to differentiate them.
Keywords: Morphometry, Dinoflagellates, Ornithocercus, Mexican Pacific.
Introducción
Uno de los órdenes más importantes de dinoflagelados planctónicos es el de los Dinophysiales, para el cual hasta el momento se han reconocido entre 240 y 382 taxa (Sournia, 1986), la mayoría distribuidos en mares tropicales y varios sólo en aguas oligotróficas y con preferencia umbrófila (Sournia, 1982).
La clasificación de los miembros del orden Dinophysiales en tres a cinco familias, dependiendo de los autores, se basa en características morfológicas como son: la forma de la teca, la posición de los poros flagelar y ventral, el grado de desarrollo de las aletas cingulares y sulcales, y la presencia de cuernos o prolongaciones de la hipoteca (Balech, 1988; Fensome et al., 1993; Schiller, 1933; Sournia, 1986; Steidinger & Tangen, 1997). El género Ornithocercus se ha clasificado dentro de Dinophysiaceae junto con Dinophysis, Histioneis, Heteroschisma, Phalacroma y Sinophysis (Fensome et al., 1993).
Debido a la dificultad para cultivar a los organismos de Ornithocercus y en si de los organismos pertenecientes a los varios géneros del orden Dinophysiales, actualmente todavía su taxonomía descansa sobre caracteres meramente morfológicos, en Ornithocercus básicamente sobre las que presenta el cuerpo central y la aleta sulcal izquierda, incluyendo la forma, el número de lóbulos y número de radios que la sostienen (Taylor, 1976; Balech, 1988). Sin embargo algunos de esos caracteres pueden ser muy variables provocando traslape entre las especies y problemas de identificación y delimitación de especies, una abundante asignación de sinonimias (Abé, 1967; Balech, 1988) y discrepancias en la concepción de las especies válidas. Por todo lo anterior para este estudio se consideraron, además de las dimensiones de los organismos, la evaluación de la forma del cuerpo central, la densidad de poros de la teca y caracteres asociados a la epiteca, a los velos cingulares y a la aleta sulcal izquierda (Balech & Ferrando, 1964).
En especial debido a la superposición de la forma, parece problemático el reconocimiento entre O. thumii (Schmidt) Kofoid et Skogsberg y O. steini Schütt, y entre O. galea (Pouchet) Abé y O. quadratus Schütt, todas presentes en el Pacífico Mexicano (PM). Entre O. steini y O. thumii se menciona que las diferencias estriban en el tamaño de la aleta sulcal izquierda y el número de costillas que la sostienen (Abé, 1967; Taylor, 1976), caracteres que sin embargo parecen ser variables. Ornithocercus galea es una especie poco citada en la literatura, erigida por Pouchet (Pouchet, 1883, In: Abé, 1967) inicialmente como una especie del género Dinophysis y después transferida por Abé (1967) al género Ornithocercus, que comparte con O. quadratus el tener la aleta sulcal de forma cuadrada, razón por la cual es considerada actualmente una variedad de O. quadratus (Taylor, 1976). Gómez (2005) valida 15 taxa de Ornithocercus a nivel mundial, sin embargo tal número discrepa del considerado en el presente estudio (Tabla1), debido a que los autores de este trabajo, con base en los resultados obtenidos, conciben de manera distinta a algunas de las especies, reconociendo a O. galea como un taxón distinto de O. quadratus y por el contrario, considerando a O. heteroporoides un sinónimo de O. bilobatus Rampi. Hasta ahora han sido registrados 12 taxa en el PM, de los cuales 4 parecen ser exclusivos de la región del Pacífico Tropical Mexicano (PTM).
Dado que durante una revisión de la colección de muestras líquidas de fitoplancton de red del laboratorio de Fitoplancton Marino y Salobre de la UAM-Iztapalapa colectadas en el PTM, se reconoció la importancia de los organismos del género Ornithocercus, tanto en número de especies como de individuos, el objetivo del presente trabajo fue estudiar la morfometría de varios organismos del mismo taxón pertenecientes, cuando se pudiera, a distintas poblaciones, para reconocer de esta manera los caracteres que permitieran la diferenciación de las especies y entonces conocer, de manera más confiable, su distribución en la región de estudio.
Materiales y métodos
Se hizo una revisión de los caracteres incluidos en las diagnosis y descripciones de las especies del género Ornithocercus registradas en el PM (según Hernández-Becerril et al., 2003; Okolodkov & Gárate-Lizárraga, 2006). La literatura taxonómica especializada consultada fue: Abé (1967), Balech (1988), Halim (1965), Jörgensen (1923), Kofoid (1907), Kofoid y Skogsberg (1928), Matzenauer (1933), Norris y Berner (1970), Rampi (1950), Schiller (1933), Sournia (1967), Taylor (1976), Wood (1953, 1954).
Se revisaron 380 muestras de la colección de Fitoplancton Marino (FpM) de muestras líquidas fijadas en formol de fitoplancton colectado con red (luz de malla 54 µm) del Laboratorio de Fitoplancton Marino y Salobre del Departamento de Hidrobiología de la Universidad Autónoma Metropolitana Iztapalapa, las cuales fueron colectadas por arrastre vertical y horizontal en las costas del PTM en 22 localidades ubicadas en los estados de Sinaloa, Nayarit, Jalisco, Colima, Michoacán, Guerrero y Oaxaca (Fig.1) entre los años 1999 - 2004, así como arrastres verticales en el Golfo de Tehuantepec en 1989 (cruceros MIMAR V, mayo y FIQUIMBI I, noviembre) y 2000 (crucero PACMEX IV, abril), cuyos datos de georreferencia se encuentran en las bases de datos BIOTICA de los proyectos CONABIO BA008 y DJ022.

Debido a que en la región de estudio se encontraron varios ejemplares de las siguientes seis especies de Ornithocercus: O. galea, O. heteroporus Kofoid, O. magnificus Stein, O. quadratus, O. steinii y O. thumii, se decidió considerarlas para realizar los estudios morfométricos.
Para cada organismo se evaluaron 29 caracteres, incluyendo tanto cuantitativos como cualitativos, estos últimos manejados como variables clasificatorias, también llamadas "dummy" (Hair et al., 1988), entre los cuales se incluyeron todos aquellos que habían sido usados para la diferenciación de las especies por taxónomos reconocidos (Taylor, 1976; Balech, 1988).
Los organismos se fotografiaron en vista lateral con una cámara digital Canon Power Shot S40 AiAF y un microscopio óptico compuesto (ML) Leica-DMLB, realizando varios enfoques del mismo organismo para así obtener los mejores rasgos de todos los caracteres (Figs 2a-2b). La longitud total del organismo se midió considerando la máxima longitud incluyendo los velos cingulares y la aleta sulcal izquierda (Fig. 2c), de la misma forma se obtuvo el ancho máximo (Fig. 2d). El largo de la célula o cuerpo central se midió desde el extremo más alto de la epiteca hasta la base de la hipoteca (Fig. 2c) mientras que la medición del ancho máximo se hizo sobre la hipoteca (Fig. 2d); para garantizar la objetividad, todas las medidas se hicieron a través de trazos geométricos como se explica más adelante. Para el área de la hipoteca se consideró el enfoque donde se evidenciara perfectamente la forma de la misma para dibujar su perímetro y permitir que el software calculara el área (Fig. 2e). Los radios fueron numerados a partir del extremo más dorsal de la aleta. Es común que entre el R1 y R3 existan una serie de radios intermedios, pero en general es fácil reconocer al R3, valorado en el análisis morfométrico, debido a que es más evidente (más robusto y a veces bifurcado) que el resto e incluso llega a conservarse a pesar de que la aleta pueda estar incompleta.

Las mediciones de los organismos se realizaron usando el Módulo de Medición del Programa Leica IM1000 V1.20 (versión 2000). En la Figura 2 a-f, se muestra al mismo organismo con distintos enfoques y la manera en que se realizó la evaluación de los caracteres cuantitativos. Para la medición sistemática de los organismos se tomó como referencia el eje del cíngulum (línea recta siguiendo la inclinación del mismo), a partir del cual se trazaron líneas paralelas y tangentes a los extremos del organismo, hasta los bordes de los velos cingulares y de la aleta sulcal izquierda, para obtener así la longitud total y la longitud de la célula (Fig. 2c). A continuación se trazó un ángulo recto en cualquiera de las líneas anteriormente trazadas para ser tomada como referente y así poder trazar líneas perpendiculares que permitieran medir el ancho máximo del organismo y de la célula (Fig. 2d). La longitud de los radios de la aleta sulcal izquierda se midió desde el extremo proximal (es decir, donde nace la aleta cerca del cuerpo central) hasta la parte proximal siguiendo la dirección del radio (Fig. 2e). Para la densidad de poros ó areolas se marcó sobre la hipoteca, un área de 100 µm2 y en ella se contaron el número de poros o areolas presentes, rectificando el dato en la medida de lo posible, a través del uso de fotografías de barrido (MEB) (Fig. 2f); las áreas de la epiteca, hipoteca y aleta sulcal fueron calculadas a través del procesador de imágenes, después de marcar el perímetro de la célula siguiendo su contorno (Fig. 2e).
Los datos obtenidos se ingresaron en una base de datos analizándose los caracteres continuos con técnicas bivariadas y multivariadas a través del programa NCSS (versión 2000). Se utilizaron Análisis de Varianza (ANOVA) para determinar la significancia estadística de las variaciones merísticas de cada uno de los caracteres morfométricos cuantitativos entre las diferentes especies consideradas. Se utilizaron Análisis de Conglomerados (AC) para observar la agrupación que presentaban los organismos con base en los caracteres evaluados, y Análisis de Discriminantes (AD) para evaluar la significación estadística en la separación de especies, con base en el conjunto de los caracteres medidos, así como para evaluar la importancia de cada variable en la separación.

Se compararon las características de cada especie descritas por los diferentes autores y obtenidas en este análisis integrándolas en una descripción para cada especie y se indicó en un mapa la distribución la de las especies de Ornithocercus encontradas en el PM, usando los registros de la base de datos BIOTICA resultado de proyectos CONABIO.
Resultados
En 165 muestras colectadas en el PTM se reconocieron 9 especies de Ornithocercus: O. bilobatus, O. galea, O. heteroporus, O. magnificus, O. orbiculatus Kofoid et Michener, O. quadratus, O. splendidus Schütt, O. steini y O. thumii (Fig. 3). A pesar de que O. bilobatus (como O. heteroporoides), O. galea y O. orbiculatus, ya habían sido reportadas anteriormente en el PTM por autores del presente trabajo (Martínez-Jasso & Meave del Castillo, 2002; Hernández-Becerril et al., 2004; Hernández-Rosas et al., 2005; Meave del Castillo & Zamudio-Reséndiz, 2005; Meave del Castillo, 2006) es la primera vez que se ilustran en una publicación. Ornithocercus cristatus no fue encontrada en la revisión de las muestras del PTM que incluyó este estudio, pero ha sido reportada para el golfo de Tehuantepec (Meave del Castillo & Hernández Becerril, 1998; Hernández-Becerril et al., 2003; Okolodkov & Gárate-Lizárraga, 2006).

Análisis morfométricos. Para el estudio morfométrico fueron medidos un total de 91 organismos: 1 de O. bilobatus, 10 de O. galea, 10 de O. heteroporus, 14 de O. magnificus, 5 de O. quadratus, 22 de O. steini y 29 de O. thumii. 30 de los ejemplares incluidos en el análisis se ejemplifican en la Figura 4.
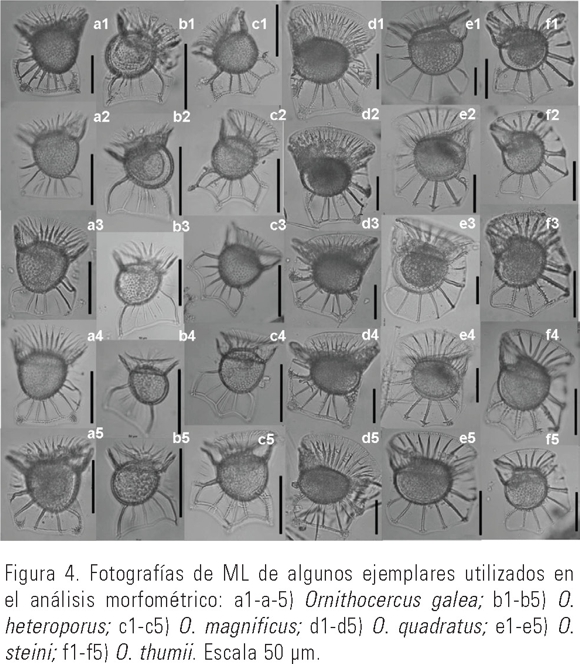
Todos los caracteres cuantitativos analizados presentaron diferencias significativas entre las especies (ANOVA), con una p<0.001. En la Tabla 3 se puede apreciar que las especies más pequeñas son O. galea, O. heteroporus, O. magnificus.
En el dendrograma resultado del Análisis de Conglomerados, de las seis especies estudiadas desde el punto de vista morfométrico, se diferencian dos grandes grupos (Fig. 5), los cuales a su vez presentan subgrupos bastante separados entre sí; el primero conteniendo a las especies O. thumii, O. steini y O. quadratus, y el segundo formado por O. magnificus, O. galea y O. heteroporus. Tanto O. thumii como O. galea se distribuyeron en diferentes subgrupos. Los subgrupos b1 y b2 de la especie O. galea (Fig. 6) se agruparon con la especie O. magnificus y O. heteroporus respectivamente, mientras que los subgrupos a1 y a2 de la especie O. thumii se agruparon con O. steini y O. quadratus respectivamente.


Los resultados del AD aplicado a las seis especies analizadas se presentan en la Figura 7, donde es evidente la no sobreposición entre las distintas especies, presentando diferencias significativas entre las especies hasta la cuarta función discriminante con p < 0.001, λ Wilks < 0.162 y un 97% de casos correctamente clasificados, traduciéndose en el hecho de que los caracteres evaluados en conjunto presentan características específicas para cada especie.
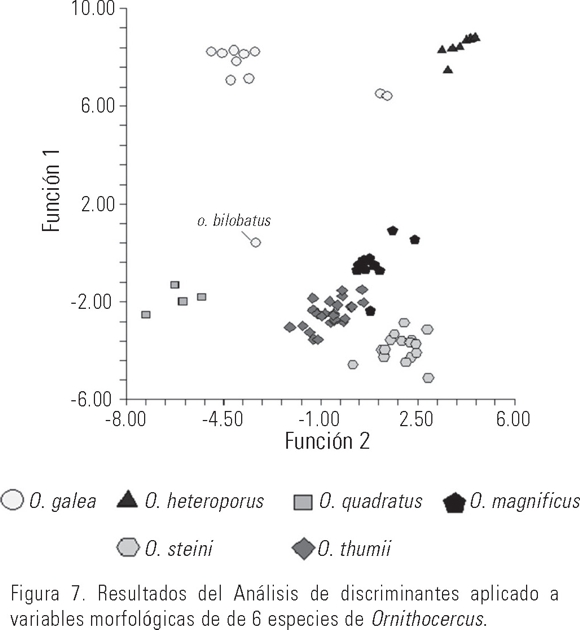
La matriz de estructura, que comprende las correlaciones de las variables morfológicas con las primeras tres funciones discriminantes (con un porcentaje acumulado del 81%) derivadas del análisis, mostró que las variables más importantes para la separación de las seis especies analizadas son: la forma asimétrica de la célula (fa), el número de lóbulos de la aleta sulcal (nls), la longitud del cíngulum (c) y la altura de los velos precingulares (avpr).
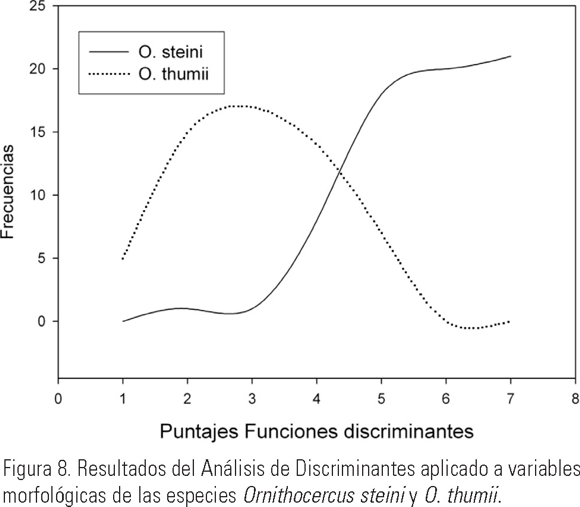
Por otro lado, en la Figura 7 se observa que el mayor grado de confusión podría presentarse entre O. thumii y O. magnificus, ya que incluso llegaron a traslaparse en el gráfico. A pesar de la semejanza morfológica entre O. thumii y O. steini, existen diferencias en las dimensiones que provoca su separación (Fig. 7). Lo anterior fue corroborado con el AD particular ambas especies (Fig. 8), encontrándose diferencias significativas con una p =0.005, λ Wilks =0.415 y un 81.4% de los casos correctamente clasificados. De acuerdo a la matriz de estructura, las variables más importantes para su separación de O. steini y O. thumii son el área de la epiteca y la longitud del cíngulum.
Distribución de las especies de Ornithocercus en el Pacífico Tropical Mexicano. Para facilitar la visualización de la distribución de las distintas especies de Ornithocercus en el PTM se muestran las Figuras 9a-d (b), donde además de los datos de distribución de las especies de Ornithocercus encontradas en las muestras colectadas y estudiadas por los autores, también se muestran todos aquellos registros a lo largo del PM, generados por otros autores y vertidos en las bases BIOTICA de proyectos CONABIO S151 y BA008. En total se tuvieron 508 registros geo-referenciados para Ornithocercus.

Las localidades donde se presentó el mayor número de especies (riqueza) de Ornithocercus se localizan en las costas de Guerrero y Oaxaca, en localidades como Acapulco, Vicente Guerrero, Punta Maldonado, Puerto Escondido, Huatulco y Golfo de Tehuantepec. La mayoría de las especies de Ornithocercus se encontraron ampliamente distribuidas en el PTM, a excepción de las raras como O. bilobatus, O. cristatus, O. splendidus y O. orbiculatus. Ornithocercus galea parece estar restringida a la porción tropical del PM y O. cristatus al Golfo de Tehuantepec (Meave del Castillo & Hernández-Becerril, 1998; Hernández-Becerril et al., 2003; Okolodkov & Gárate-Lizárraga, 2006).
Descripciones de especies del género Ornithocercus estudiadas en el Pacífico Tropical Mexicano.
Ornithocercus bilobatus Rampi (Fig. 3b)
Referencias: Rampi, 1950, p.3, Fig.12; Abé, 1967, p. 83, Fig. 29 a-c.
Sinonim. O. heteroporoides Abé.
Descripción: El único ejemplar encontrado midió 101 µm de largo total y 82.5 µm de ancho máximo, con forma intermedia entre O. galea y O. heteroporus, pero con cuerpo celular más esférico: de 42 µm de largo y 45.5 µm de ancho. Altura del velo precingular: 28 µm y del postcingular: 22 µm. La aleta sulcal, aunque también oblicua y triangular como en O. heteroporus, alcanzaba el borde dorsal de la célula y presentó una costilla longitudinal muy evidente desde el R3 hasta el R1, sin embargo, localizada más internamente que en O. heteroporus. R3 de 22 µm de largo. Borde superior de la epiteca bastante más inclinado que en O. galea y que en O. heteroporus. Entre el R1 y el R3 se presentaban 3 radios intermedios y otro más incompleto. La densidad de poros fue aproximadamente de 14 en 100 µm2.
Distribución: Este taxón es raro en el PTM, únicamente se encontró en la localidad de Puerto Escondido, Oax., muestra líquida FpM1002, en una temperatura de 27°C en el mes de septiembre. La especie también parece ser muy rara a nivel mundial, ya que únicamente se había reportado para la bahías de Saruga y Sagami (Abé, 1967).
Comentarios: Taylor (1976) propone que O. heteroporoides es sinónimo de O. bilobatus Rampi (1950) aspecto que aceptan los autores de este trabajo. Abé (1967) señala que el cuerpo celular de O. heteroporoides mide de 50-65 µm de largo y Rampi (1950) da una medida de 34 µm para O. bilobatus, por lo que el largo del cuerpo central del ejemplar encontrado en el PM (42 µm), está dentro del intervalo de la especie; también el ancho máximo de la célula se correspondió porque el ejemplar encontrado midió 55 µm y para la especie se da un intervalo de 34-55 µm (Abé, 1967; Rampi, 1950). Hay que mencionar que las dimensiones de O. bilobatus se traslapan con las de los ejemplares de O. magnificus del PTM, la otra especie que puede presentar la costilla longitudinal en situación interna, como se muestra en la Figura 4 (C1-C5). No obstante, Kofoid y Skogsberg (1928) señalan que en ocasiones pueden faltar uno e incluso dos de los tres lóbulos que O. magnificus presenta en su aleta sulcal izquierda, en este trabajo se validó al taxón O. bilobatus utilizando además como criterio la densidad de poros que presenta la teca, ya que en esta especie el número fue de 14 en 100 µm, mientras que en los ejemplares de O. magnificus del PTM, fue prácticamente de 10.
Ornithocercus galea (Pouchet) Abé (Figs. 3 a, 4 a1-a5, 6 a-i, 11 C)


Referencias: Pouchet, 1883, p. 426, Fig.6 (In: Abé 1967); Schütt, 1900 (partim), Figs. 3, 5-6; Kofoid y Skogsberg, 1928 (partim f. simplex f. intermedia), p. 561, Figs. 86-87; Schiller, 1933 (partim), p. 204, Figs.194 c-f; Abé, 1967, p. 89, Fig. 33a-b; Taylor, 1976 (partim O. quadratus var. assimilis, O. quadratus f. simplex), p. 50-51, Lám. 8, Figs. 79, 81- 82.
Sinonim. Dinophysis galea Pouchet (partim); O. quadratus Schütt (partim). O. quadratus var. assimilis (Jörgensen) Taylor; O. quadratus var. quadratus f. simplex Kofoid et Skogsberg, O. assimilis Jörgensen.
Descripción: Organismos de 89-163 µm de largo total y 74-129 µm de ancho máximo. Cuerpo celular ovoide con la parte posterior agudizada y el ancho máximo ubicado en la parte superior cerca del cíngulum, célula asimétrica (inclinada), de 41-83 µm de largo y 45-88 µm de ancho; en el PTM se encontraron dos conjuntos de organismos con dimensiones distintas. Epiteca ancha con borde superior recto o ligeramente convexo. Velos cingulares de 17 a 37 µm de altura, sostenidos por radios fuertes. Aleta sulcal izquierda cuadrada, no muy grande pero que alcanza a llegar al borde dorsal del organismo. La anchura de dicha aleta sulcal, en el lado ventral del organismo, disminuye hacia la parte superior a partir del R3 y está sostenida por un número variable de radios (3-7), localizados entre R1 y R3; en ocasiones se observan ramificaciones en las puntas de los radios que forman las esquinas del borde posterior de la aleta, o también suele presentarse una costilla longitudinal que corre paralela y muy pegada al margen. El R3 mide de 17 a 38 µm de largo y es variable, en ocasiones robusto y en posición casi horizontal, otras veces más delicado y con posición más inclinada, en ambos casos se desvían abruptamente hacia la parte posterior para formar la costilla longitudinal. Poros evidentes, densidad 10-12 en 100 µm2.
Distribución: La especie se encontró en las localidades de: San Blas, Nay., Punta Mita, Nay., Cabo Corrientes, Jal., El Faro, Mich., Caleta Campos, Mich., Zihuatanejo, Gro., Barra de Potosí, Gro., Acapulco, Gro., Punta Maldonado, Gro., Chacahua, Oax., Puerto Escondido, Oax., Huatulco Oax.
Muestras líquidas (FpM): 463, 503, 505, 517, 519, 537, 684, 874, 878, 669, 868, 873, 882, 884, 957, 961, 962, 964, 984, 1001, 1002, 1005, 1027, 1029, 1031, 1060 y 1143, 1239, 1242, 1389, 1390, 1391; con un intervalo de temperatura entre los 22-29 °C, en los meses de marzo, mayo, julio, septiembre y noviembre.
Comentarios: La especie fue reconocida tanto por Pouchet (1883, como Dinophysis galea) como por Jörgensen (1923, como Ornithocercus assimilis). Y fue Abé quien hizo una nueva combinación: O. galea (Pouchet) Abé. Sin embargo Kofoid y Skogsberg (1928) sobrevaloraron la forma de la aleta y consideraron que todos los organismos que tenían una aleta sulcal cuadrada pertenecían a O. quadratus reconociendo así a O. assimilis como una forma de O. quadratus, posición que adoptaron la mayoría de los taxónomos posteriores. La var. assimilis estuvo presente en varias localidades del Océano Índico estudiadas por Taylor (1976). Dicho autor menciona que al principio reconoció a O. assimilis como un taxón distinto, pero después fue encontrando formas intermedias entre O. assimilis y O. quadratus que lo hicieron reconsiderar su idea inicial y concebirlo como una variedad de O. quadratus y a la forma simplex de O. quadratus var. quadratus como un probable sinónimo de la var. assimilis, aunque también señala que ambos taxa requieren de una revisión más profunda para esclarecer su estatus. Probablemente Taylor (1976) no le da validez a O. galea porque el esquema de Pouchet (1883) parece ser una combinación de distintos taxa. Abé (1967) señala que sólo la cuarta figura de Pouchet (1883, In: Abé 1967) debe considerarse el iconotipo de O. galea. Taylor (1976) señala sin embrago, que ciertas características tanto de la var. assimilis como de la forma simplex de O. quadratus, realmente se corresponden con las de O. galea y mismas que fueron encontrados en los ejemplares del PTM, como es el hecho de que la parte superior de la aleta sulcal en el lado dorsal esté incompletamente desarrollada. El velo cingular de O. galea puede ser variable: algo bilobulado como se muestra en la Figura 11-c, o bien mucho más reducido semejando sólo superficialmente a O. cristatus, como en la Figura 6g-h. La consideración de los autores del presente trabajo para descartar la posibilidad de que se tratara de O. cristatus se basó en el hecho de que tales ejemplares no presentaban las costillas del velo precingular bifurcadas como ocurre en O. cristatus y su velo cingular surgía en una dirección más dorsal que lo señalado para esta última (Matzenauer, 1933). El hecho de que O. galea sea muy común en la porción tropical del Pacífico Mexicano y que no haya sido encontrada por Licea et al. (1995) o por Hernández-Becerril (1988a, b) indica que su distribución está asociada a la porción tropical del PM.
Ornithocercus heteroporus Kofoid (Figs. 3c, 4b1-b5)
Referencias: Kofoid, 1907, p. 206, Lám. 12, Fig. 70; Jörgensen, 1923, p. 38, Fig. 54; Kofoid & Skogsberg, 1928, p. 517, Fig. 75, Lám. 18, Figs. 1-3; Schiller, 1933, p. 195, Fig. 178a-d; Rampi, 1940; p. 265, Fig. 53; Wood, 1953, Fig. 58 a-c; Wood, 1954, p. 210-211, Fig. 65-66; Abé, 1967, p. 85-86, Fig. 28a-b; Taylor, 1976, p. 48-49, pl. 8 Fig. 83; Balech, 1988, p. 59, Lám. 14, Fig. 4; Hernández-Becerril, 1988b, p. 427, Fig. 9; Licea et al., 1995, p. 23, Lám. 21, Fig. 11.
Sinonim. O. biclavatus Wood; O. triclavatus Wood.
Descripción: Especie pequeña de 67-90 µm de longitud total y 57-69 µm de ancho máximo. Cuerpo celular elíptico, con el ancho máximo cerca de la porción media, asimétrico muy inclinado, de 31-40 µm de largo y 33-41 µm de ancho. Epiteca pequeña y convexa, con borde superior reducido dando al velo precingular una ligera apariencia de embudo. Velos cingulares de 13-23 µm de altura. Ancho a nivel del cíngulum (en vista lateral) de 24-31 µm. La aleta sulcal izquierda oblicua con forma triangular y extendida solamente hasta la mitad del polo posterior de la célula, presenta dos lóbulos en las esquinas correspondientes a los radios R1 y R3 y entre ellos se presenta un número variable de radios (4-6); en el borde posterior de la aleta sulcal se forma una costilla longitudinal evidente. Longitud de R3 13-15 µm y 20 poros en 100 µm2.
Distribución: La especie se encontró en las localidades de: Chacala, Nay., Punta Mita, Nay., Bahía Manzanillo, Col. Caleta Campos, Mich., Playa Azul, Mich., Zihuatanejo, Gro., Acapulco, Gro., Punta Maldonado, Gro., Puerto Escondido, Oax. y Golfo de Tehuantepec. Muestras líquidas (FpM): 40, 41, 51, 55, 94, 107, 110, 512, 517, 519, 537, 538, 544, 868, 885, 984, 1005, 1058, 1060, 1076, 1081, 1128, 1132, 1143, 1147, 1226, 1283, 1285, 1387, 1432; con una temperatura entre los 22-28 °C en los meses de marzo, abril, mayo, julio, septiembre, octubre y noviembre.
Comentarios. Taylor (1976) hace sinónimos de Ornithocercus heteroporus a O. biclavatus y O. triclavatus, evidenciando también las diferencias de este conjunto de especies con O. bilobatus. Ya que en esta última la aleta sulcal izquierda se extiende hasta la parte dorsal de la hipoteca, como se observa en la Figura 3b.
Ornithocercus magnificus Stein emend. Schütt (Figs. 3d, 4c1-c5, 11 a-b)
Referencias: Stein, 1883, partim, Lám. 23, figs. 1-2; Schütt, 1900, Figs. 8, 10; Jörgensen, 1923, p. 35, Fig. 48; Kofoid & Skogsberg, 1928, p. 529, Fig. 79, Schiller, 1933, p. 301, Fig. 190a-b, Wood, 1954, p. 203, Fig. 60a-b; Abé, 1967, p. 88, Fig. 32 a-d; Balech, 1988, p. 61, Lám. 14, Figs. 7-8; Taylor, 1976, p. 49, Lám. 7. Figs. 67-69, Lám.42, Fig. 505a-b; Hernández-Becerril, 1988c, p.193, Figs. 9-10; Licea et al.,1995, p. 23-24, Lám. 8, Fig. 3.
Descripción: Especie pequeña semejante a O. thumii por la aleta lobulada, de 66-104 µm de largo total y 59-85 µm de ancho máximo. Cuerpo celular redondeado de 33-45 µm de largo y 33-47 µm de ancho. Epiteca con borde apical recto, algo inclinado. Ancho a nivel del cíngulum 29-40 µm (en vista lateral). La altura de los velos cingulares es de 14-29 µm, siendo más alto el precingular. Aleta sulcal trilobulada debido a que se extienden hasta el margen, de sus tres radios. Generalmente se presenta una costilla submarginal. Longitud de R3 14-24 µm. Entre el R1 y el R3 pueden existir de 4-5 radios adicionales. La teca tiene una densidad promedio de 10 poros en 100 µm 2.
Distribución: Mazatlán, Sin., San Blas, Nay., Chacala, Nay., Punta Mita, Nay., Cabo Corrientes, Jal., El Faro, Mich., Caleta Campos, Mich., Playa Azul, Mich., Ixtapa, Gro., Zihuatanejo, Gro., Barra Potosí, Gro., Vicente Guerrero, Gro., Acapulco, Gro. Punta Maldonado, Gro., Chacahua, Oax., Puerto Escondido, Huatulco, Oax., y Golfo de Tehuantepec. Muestras líquidas (FpM): 40, 55, 79, 82, 83, 84, 94, 98, 105, 107, 110, 111, 112, 463, 503, 507, 509, 512, 517, 519, 521, 529, 537, 539, 544, 546, 548, 550, 552, 553, 554, 556, 669, 671, 684, 775, 868, 874, 876, 878, 882, 883, 884, 885, 886, 927, 930, 938, 939, 940, 941, 943, 947, 950, 952, 953, 954, 957, 958, 961, 962, 964, 978, 984, 986, 988, 1002, 1005, 1006, 1008, 1025, 1026, 1027, 1029, 1030, 1031, 1076, 1078, 1080, 1083, 1105, 1128, 1131, 1132, 1191, 1283, 1189, 1240, 1242, 1243, 1244, 1285, 1226, 1240, 1244, 1280, 1281, 1370, 1377, 1379, 1382, 1384, 1389, 1390, 1420, 1424, 1426, 1427, 1428, 1429, 1431; 1432, 1507, 1571, 1634; con un intervalo de temperatura entre 22-27 °C, en los meses de febrero, marzo, abril, mayo, julio, septiembre, octubre y noviembre.
Comentario: Es una de las especies menos conflictivas en su reconocimiento, aunque algunos autores mencionan que podría estar relacionada con O. thumii (Taylor 1976; Balech, 1988); sin embargo, esta última se diferencia de O. magnificus porque no presenta un radio en el lóbulo medio, además de que O. magnificus es de tamaño menor. Además se ha mencionado que en O. magnificus no sólo uno, sino hasta dos de sus lóbulos pueden faltar (Kofoid & Skogsberg, 1928).
Ornithocercus orbiculatus Kofoid et Michener (Fig. 3e).
Referencias: Kofoid & Michener, 1911, p. 300, Kofoid & Skogsberg, 1928, p. 559, Lám. 17, Fig. 7; Balech, 1988, p. 61, Lám 15, Fig. 2.
Descripción: Especie de 118 µm de largo máximo y 108 µm de ancho máximo, cuerpo celular ensanchado de 64 µm de largo y 75 µm de ancho. Velos cingulares de 13 a 23 µm de altura, semejantes a los de O. steini, pero soportados por un número de radios que va de 12 a 15 y muchos más radios pequeños hacia la orilla. La aleta sucal izquierda terminando hasta la porción dorsal de la célula, con un margen subcircular y pequeños lóbulos dorsales, con 5 costillas que pueden terminar en reticulaciones marginales. Densidad de poros de la teca: 6.5 en 100 µm2.
Distribución: Se encontró un solo ejemplar en el PTM, en la localidad Golfo de Tehuantepec, estación 45 (14º 32.782' N, 93º 36.232') Crucero PACMEX III realizado en el mes de abril; muestra líquida FpM939. La especie parece ser realmente rara, Kofoid y Skogsberg (1928) sólo encontraron un ejemplar en el Pacífico en el área de Panamá (estación 4617) y Balech (1988) sólo encontró dos ejemplares en el Atlántico sudoccidental.
Comentarios: Taylor (1976) hace sinónimo a O. orbiculatus de O. steini. Kofoid y Michener (1911) consideran que "este taxón" puede corresponder a un fase de reconstrucción de la teca de O. steini y sin embargo la erigen como especie. Las medidas del ejemplar del PTM concuerdan con las descritas por Kofoid y Skogsberg (1928) y con las de Balech (1988), sin embargo también están dentro del intervalo de medidas señalado para O. steini. Los autores del presente trabajo validaron a la especie por el número de radios en los velos cingulares, los lóbulos presentes en la parte ventral superior de la aleta sulcal izquierda y porque la densidad de poros de la teca también fue menor a lo que presenta O. steini.
Ornithocercus quadratus Schütt (Figs. 2 a-f, 3f, 4 d1-d5, 11 d)
Referencias: Schütt, 1900 p. 5, Figs.1-4, 12-13; Jörgensen, 1923, Fig. 50; Kofoid & Skogsberg, 1928, partim, p. 463, Fig. 86; Kofoid & Skogsberg, 1928, partim, p. 567, Fig. 87; Schiller, 1933 partim, p. 204, Fig. 194 a-b; Wood, 1954, p. 209, Fig. 63 a-c; Abé, 1967, p. 89-90, Fig. 33 a.b; Taylor, 1976, p. 50, Lám 8, Fig. 77-78, 80; Balech, 1988, p. 60, Lám. 14, Fig. 10; Hernández-Becerril, 1988b, p. 427, Figs. 11, 41.
Sinonim. O. quadratus f. schütti Kofoid et Skogsberg, O. quadratus f. intermedia Kofoid et Skogsberg.
Descripción: Organismos grandes de 133-171 µm de largo total y 104 a 135 µm de ancho máximo. Célula más ancha que larga de 59-66 µm de ancho y 71 a 80 µm de largo. Epiteca muy aplastada e inclinada hacia la porción ventral. Velos cingulares prácticamente del mismo tamaño, 42 a 55 µm de altura. Con radios fuertes en la base y costillas más débiles en la porción distal que no llegan a la mitad de los velos. Aleta sulcal izquierda bastante extendida, llegando hasta la parte media de la hipoteca en el lado dorsal, con esquinas cuadradas, frecuentemente sin lóbulos, aunque a veces si lo presenta, como en las Figura 3f y 4d1-4d2, 4d4; generalmente el R3 más largo dándole mayor anchura a la aleta justo en ese sitio y presenta una costilla longitudinal en el borde posterior de la aleta. Longitud del R3: 29-40 µm, en ocasiones en posición casi horizontal y en otras inclinado, frecuentemente el R3 se ve más grueso, como si estuviera bifurcado desde la base y comúnmente formando una reticulación en la parte distal. Es común que se formen en las esquinas reticulaciones por debajo de la costilla longitudinal, semejando borlas. Constantemente se presentan 5 radios entre R1 y R3, y es característico que después del R1 se presentan 3 radios divergentes, justo en la porción basal de la célula. Poros grandes, 4.5-6 poros en 100 µm2, en ocasiones poco evidentes en microscopio de luz. Es frecuente la presencia de feosomas (cianobacterias simbiontes) en los velos cingulares.
Distribución. La especie se encontró en pocas muestras, en las localidades de: Punta Mita, Nay., Barra de Navidad, Nay., Bahía Manzanillo, Col., El Faro, Mich., Caleta de Campos, Mich., Playa Azul, Mich., Zihuatanejo, Gro., Barra de Potosí, Gro., Acapulco, Gro. Punta Maldonado, Gro., Puerto Escondido, Oax., Huatulco, Oax., y Golfo Tehuantepec. Muestras líquidas (FpM): 107, 519, 529, 539, 544, 756, 769, 780, 868, 870, 884, 886, 952, 961, 962, 964, 1005, 1027, 1076, 1078, 1080, 1082, 1131, 1143, 1147, 1214, 1229, 1239, 1243, 1244, 1283, 1388, 1428, 1431; con temperaturas de 22-25 °C, en los meses de marzo, abril, mayo, junio, julio, agosto, septiembre, octubre y noviembre.
Comentarios. Las dimensiones de los organismos concuerdan con el intervalo señalado por Abé (1967) y Balech (1988) para la longitud de la célula.
Sin embargo, los esquemas de Balech (1988) y Abé (1967) muestran organismos con la aleta sulcal izquierda perfectamente cuadrada, mientras que los organismos del PTM se parecen más a los esquematizados por Taylor (1976) como O. quadratus var. quadratus, asimismo la célula es mucho más ancha que larga, como lo muestran los mismos esquemas de Taylor, a diferencia de la descripción de Balech (1988), quien menciona que el cuerpo celular de O. quadratus prácticamente es isodiamétrico, y que sólo en algunos organismos el largo de la célula puede ser menor.
Ornithocercus steini Schütt (Figs. 3h, 4e1-e5, 10b, 11e)
Referencias: Schütt, 1900, Figs. 5-7; Kofoid, 1907, p. 206, Lám.15, Fig. 93; Jörgesen, 1923, p. 32, 38, Figs. 49, 52; Schiller, 1933, p. 202, Fig. 192; Kofoid & Skogsberg, 1928, p. 551, Figs. 83:1-12; Wood, 1953, p. 203, Fig. 62; Taylor, 1976, p. 52-53, Lám. 7, Fig. 72; Balech, 1988, Lám. 15, Fig. 1; Licea et al., 1995, p. 24, Lám. 8, Fig. 4.
Sinonim. O. serratus Kofoid.
Descripción: Organismos de (92) 120-184 µm de largo total y 56.5-84.1 µm de ancho máximo. Cuerpo celular subesférico ligeramente más ancho que largo, de 54-98 µm de largo y 57-102 µm de ancho; ancho a nivel del cíngulum 53-85.5 µm, epiteca chata e inclinada hacia la porción ventral, velos cingulares de 28 a 53 µm de altura, el precingular con radios muy fuertes, el postcingular con radios más numerosos que en ocasiones se anastomosan por pares en la parte media para formar uno sólo en la porción distal (Fig. 11), aleta sulcal izquierda muy amplia, que en ocasiones sobrepasa la mitad de la hipoteca del lado dorsal, generalmente presenta cuatro lóbulos posteriores poco marcados, cada uno sostenido por un radio y el cual, principalmente en el borde de la porción distal de la aleta, presenta un ramillete de ramitas. R3 de 25-50 µm de largo, en posición inicialmente inclinada y después vertical, 3-7 radios entre R1 y R3 , costilla longitudinal generalmente ausente, rara vez presente. Densidad de poros en la teca 8 en 100 µm2. La especie comúnmente presenta feosomas en los velos cingulares.
Distribución: Bahía de Mazatlán, Sin., San Blas, Nay., Punta Mita, Nay., Manzanillo, Col., El Faro, Mich., Caleta Campos, Mich., Zihuatanejo, Gro., Acapulco, Gro., Barra Potosí, Gro., Vicente Guerrero, Gro., Acapulco, Gro., Punta Maldonado, Gro., Chacahua, Oax., Puerto Escondido, Oax., Huatulco, Oax., y Golfo de Tehuantepec. Muestras líquidas: (FpM): 41, 51, 79, 91, 102, 107, 111. 503, 505, 519, 522, 539, 544, 548, 566, 731, 736, 882, 868, 874, 882, 883, 884, 885, 937, 940, 946, 957, 958, 962, 984, 1001,1002, 1008, 1076, 1078, 1079, 1983, 1128, 1130, 1143, 1191, 1277, 1281, 1283, 1384, 1391, 1420, 1422, 1423, 1426, 1432, 1519; con temperaturas entre los 22-29 °C., en los meses de marzo, abril, mayo, junio, julio, septiembre y noviembre.
Comentarios. Abé menciona que el nombre de la especie es O. steini. En la literatura más reciente es común encontrar el nombre de esta especie escrito con doble i (O. steinii). Según Abé (1967) parece que este fue un error introducido por Schiller (1933), que fue heredado por los autores subsiguientes.
Ornithocercus splendidus Schütt (Fig. 3g)
Referencias: Schütt, 1893, p. 272, Fig. 82; Schütt, 1896, p.10, Fig. 13B; Kofoid & Skogsberg, 1928, p. 521, Fig. 77:1-11, 85:3, Lám. 16, Fig. 2, 4; Schiller, 1933, p. 196, Fig. 189; Abé, 1967, p. 81, Fig. 27a-c; Taylor, 1976, p. 52, Lám. 8, Figs. 85-86, Lám. 40, Fig. 486, Lám. 42, Fig. 504; Balech, 1988, p. 59, Lám. 14, Figs. 5-6; Licea et al., 1995, p. 24.
Sinonim: O. splendens Schütt.
Descripción: Organismos grandes, de 133-165 µm de largo máximo y 122-126 µm de ancho máximo. Aleta sulcal bilobulada que no llega a alcanzar ni siquiera la mitad de la parte posterior de la célula. Cuerpo celular ovoide (con extremo apical ensanchado) de 43-48 µm de largo y 46-47 µm de ancho. Aletas cingulares muy desarrolladas de 34-45 µm de altura sostenida por numerosos radios fuertes que frecuentemente se ramifican formando una compleja reticulación. Aleta sulcal izquierda oblicua y margen posterior cóncavo, los extremos de los lóbulos puntiagudo-redondeados con fuerte reticulación semejando pequeñas borlas. R3 de 20-23 µm de largo, 4 radios entre R1 y R3. Teca con pequeños poros dispersos difíciles de medir en microscopio óptico.
Distribución: San Blas, Nay., Bahía Manzanillo, Col. y Acapulco, Gro. Muestras líquidas (FpM): 503, 522 y 1370; con temperatura promedio de 28 °C, en los meses de mayo, julio y octubre.
Ornithocercus thumii (Schmidt) Kofoid et Skogsberg. (Figs. 3i, 4f1-f5, 10a, 11f)
Referencias: Schmidt, 1888, In: Schmidt 1874-1902, Lám. 144, Figs. 59-61; Stein, 1883, partim, Lám. 23, Figs. 4-5; Jörgensen, 1923, p. 36, Fig. 49; Kofoid & Skogsberg, 1928, partim, p. 540, Fig. 81-82, Lám. 18, Figs. 4-6; Schiller, 1933, p. 200, Fig.191; Taylor, 1976, p. 53, Lám. 7, Fig. 71-72, 74; Balech, 1988, p. 61, Lám. 14, Fig. 9; Hernández-Becerril, 1988b, p. 427, Fig. 12; Licea et al., 1995, p. 24, Lám 8, Fig. 5.
Sinonim. Parelion thumii Schmidt, O. magnificus Stein.
Descripción: Especie semejante a O. magnificus por tener la aleta lobulada, pero de mayores dimensiones, 108-184 µm de largo máximo y 56-139 µm de ancho máximo. Con epiteca más grande y chata, cuerpo celular de 48-90 µm de largo, y 53-97 µm de ancho. Velos cingulares de 25-50 µm de altura, más ornamentados que en O. magnificus. Ancho en vista lateral a nivel del cíngulum 45-78. La aleta sulcal izquierda frecuentemente llega hasta la porción media de la hipoteca en el lado dorsal, formando tres lóbulos; uno posteroventral, otro antapical y otro posterodorsal. El R3 mide 22-44 µm de longitud. Entre R1 y R3 se presentan 3-5 radios. Densidad promedio de poros de la teca: 5-6 en 100 µm2.
Distribución: Bahía Mazatlán, Sin., San Blas, Nay., Punta Mita, Nay., Bahía Manzanillo, Col., El Faro, MIch., Caleta Campos, Mich., Playa azul, Mich., Ixtapa, Gro., Zihuatanejo, Gro., Barra de Potosí, Gro. Acapulco, Gro., Punta Maldonado, Gro., Chacahua, Oax., Puerto Escondido Oax., Huatulco, Oax. y Golfo Tehuantepec. Muestras líquidas (FpM): 40, 41, 43, 51, 55, 70, 79, 83, 102, 107, 111, 463, 503, 504, 505, 507, 509, 512, 517, 518, 519, 521, 522, 537, 548, 553, 565, 684, 736, 866, 868, 882, 883, 884, 886, 870, 874, 878, 932, 936, 937, 938, 939, 940, 941, 943, 944, 946, 947, 950, 952, 954, 956, 957, 958, 959, 861,978, 984, 985, 1002, 1006, 1008, 1027, 1029, 1048, 1060, 1076, 1078, 1079, 1080, 1083, 1103, 1128, 1130, 1131, 1143, 1147, 1189, 1213, 1224, 1239, 1241, 1243, 1275, 1276, 1277, 1278, 1280, 1281, 1282, 1283, 1285, 1370, 1373, 1376, 1377, 1378, 1379, 1383, 1384, 1387, 1388, 1389, 1390, 1391, 1419, 1420, 1422, 1423, 1424, 1426, 1427, 1571, 1634; con temperaturas de 27 a 30 °C, en los meses de marzo, abril, mayo, junio, julio, septiembre, octubre y noviembre.
Discusión
Validación de especies controvertidas. Los organismos del PTM nombrados como O. galea presentaron variación en las dimensiones y en la forma de la aleta sulcal izquierda, la presencia o no de lóbulos con ramificaciones en los extremos de la aleta, la cuadratura de la misma y la presencia de la costilla longitudinal. Abé (1967) ha mencionado que esa aleta era sumamente variable en la especie y que inclusive podía estar ausente. El mismo autor señala que para O. galea, la relación entre el ancho de la epiteca con respecto al ancho de la parte superior de la hipoteca es cercana a 0.7, lo cual se cumplió para los organismos encontrados en el PTM (con valores de 0.6-0.7). Las dimensiones de los organismos de O. galea del PTM, considerando ambos grupos, fueron los siguientes: largo célula (41) 48-57 (83) y ancho célula 45-70 (88). Basándonos en el dendograma de la Figura 5 se obtuvieron por separado, los intervalos por cada uno de los subgrupos diferenciados de O. galea (Tabla 4), cuyos esquemas se muestran en la Figura 6. Los del tipo b1 (más relacionados con O. magnificus) tuvieron las siguientes dimensiones del cuerpo celular: 47-57 (83) µm de largo y 47-70 (88) µm de ancho máximo; mientras que los organismos del tipo b2, más relacionados con O. heteroporus, fueron más pequeños: 41-52 µm de largo y 45-54 µm de ancho máximo. Estos últimos coinciden mejor con la descripción original de la especie (Abé, 1967). No obstante, las diferencias en las dimensiones de los subgrupos de O. galea (Tabla 3) no fueron valoradas a nivel taxonómico, toda vez que otras especies de fácil reconocimiento, como el caso de O. magnificus, también son muy variables. Se concibió la existencia de los dos grupos de O. galea en el PTM, como formas distintas para la especie, las cuales podrán explicarse en un estudio posterior.
Ornithocercus assimilis, es considerada por varios taxónomos un sinónimo de O. quadratus (Kofoid & Skogsberg, 1928; Abé, 1967; Taylor, 1976; Balech, 1988); sin embargo, en este estudio es considerado sinónimo de O. galea porque se valoró la forma asimétrica del cuerpo celular y también la forma y dirección del R3 ligeramente posterior y menos desarrollado que los siguientes radios originando que la aleta disminuya su anchura justo a la altura del R3 y diferenciándola así de las de O. quadratus, quien presenta el R3 muy grueso e incluso en ocasiones en posición prácticamente horizontal, provocando que la aleta se ensanche a ese nivel, o mantenga la forma totalmente cuadrada desde la porción superior hasta la inferior.
Como puede observarse en la Figura 7, el AD para el conjunto de especies ayudó también a rectificar la identificación de algunos ejemplares, así un ejemplar originalmente identificado como O. galea, que el AC ubicó fuera del grupo e incluso alejado de cualquier otro (señalado en la Figura 7 con una flecha, e ilustrado en la Figura 3b), guió a revisar nuevamente la morfología del organismo y su identificación, encontrándose que la aleta sulcal era asimétrica y que la costilla longitudinal estaba situada más adentro que el extremo de la aleta, además de que presentaba ramificación del R1, todo lo cual coincidió con la descripción tipo de O. bilobatus (Abé, 1967).
Aunque en este estudio se reconoció a O. orbiculatus, debido a las semejanzas del único ejemplar encontrado en el Pacífico con la descripción de Kofoid y Skogsberg (1928), es necesario reconocer que puede tratarse de un taxón inválido, toda vez que se ha mencionado que podría ser una etapa de vida de O. steini (Kofoid & Skogsberg, 1928). Sin embargo, este ejemplar tuvo una densidad de poros de la teca menor que lo que presenta O. steini (6.5 vs. 8.0). Para aclarar completamente su situación se tendría que realizar un estudio morfométrico que incluyera una mayor cantidad de ejemplares de O. orbiculatus, lo cual parece una tarea difícil debido a la rareza de la especie.
Morfometría de O. steini y O. thumii. En lo que respecta a los resultados del análisis morfométrico de ambas especies se encontró que efectivamente son taxa muy parecidas que además comparten el mismo ambiente, toda vez que frecuentemente estuvieron presentes en la misma muestra, lo que incluso puede explicar el hecho de que hayan llegado a ser confundidas, como le ocurrió a Jörgensen (1923). Sin embargo, el AD mostró que ambos taxa se diferenciaban bien (Fig. 8), toda vez que sólo 4 de los 43 organismos estuvieron mal clasificados desde el punto de vista estadístico. En la Figura 10 se observa además, que la forma de la aleta sulcal izquierda es bastante diferente en ambas especies, redondeada para O. steini (Fig. 10a) y angulosa para O. thumii (Fig. 10b), como lo señalan Steidinger y Tangen (1997). Dado que en ocasiones dicha aleta está ausente o se presenta de manera muy reducida, fue interesante el que este estudio evidenciara que otro carácter, no relacionado con la aleta, tuviera diferencias significativas entre ambas especies y además estuviera altamente correlacionado con la anchura de la célula a nivel del cíngulum (carácter que es más fácil de medir), por lo que al parecer es posible diferenciar a ambas especies por ciertas dimensiones, de tal forma sí el cíngulum es <60 µm el organismos corresponde a O. thumii, mientras que sí el área de la epiteca es >800 µm2 corresponde a O. steini.
Distribución de las especies. Prácticamente en el PM se han encontrado ya a todas las especies posibles del género Ornithocercus. Ornithocercus quadratus parece ser una especie ampliamente distribuida ya que ha sido encontrada en la porción subtropical templada del PM (Okolodkov & Gárate-Lizárraga, 2006) y ahora se encontró prácticamente a todo lo largo del PTM. Se han encontrado pocos ejemplares de O. splendidus en el PTM y posiblemente eso se deba a que la especie sea más oceánica y se han estado trabajando estaciones costeras; de cualquier manera en la Figura 9b se observa que se trata de una especie con distribución mas bien tropical, desde la boca del Golfo de California (Okolodkov & Gárate-Lizárraga, 2006) hasta Huatulco, Oax. y que al parecer está ausente en el Golfo de Tehuantepec. Ornithocercus splendidus fue registrada por Kofoid y Skogsberg (1928) a lo largo de todo el Pacífico Oriental, tanto en la corriente de California, como en la corriente Oeste Mexicana, el Área Panámica y la corriente de Perú.
Ornithocercus galea también parece estar restringida al PTM y O. cristatus al Golfo de Tehuantepec. Hasta el momento sólo se ha encontrado un ejemplar de O. bilobatus y otro de O. orbiculatus, pero la ausencia de estos taxa en la porción subtropical templada, cuyo fitoplancton ha sido más estudiado, hace pensar que también estén restringidos a la porción tropical del PM.
Otras especies tales como O. skogsbergii y O. formosus, no encontradas en este estudio, parecen estar presentes únicamente en la porción subtropical templada del PM (Martínez-López, 1993; Martínez-López & Verdugo-Díaz, 2000; Hernández Becerril et al., 2003; Licea et al., 1995, Lám. 8, Fig. 4; como O. steini). Una de las especies faltantes en el PM es O. francescae, la cual fue descrita primeramente en el mar Mediterráneo y la cual definitivamente no parece ser nativa de la región, toda vez que Kofoid y Skogsberg (1928) tampoco la encontraron en su estudio del Pacífico Oriental, que abarcó desde California hasta la corriente de Perú.
De acuerdo con lo anterior se concluye:
La flora del género Ornithocercus es rica en el PTM.
Se confirma la presencia de O. galea en el PTM, considerando a O. assimilis y a O. quadratus var. assimilis como sus sinónimos.
Se observa que O. quadratus se diferencia claramente de O. galea. Por ello a pesar de que ambas especies presenten la aleta sulcal izquierda de forma cuadrada, se valoró la forma de la célula (ovoide asimétrica o esférica ensanchada) y de la epiteca (convexa redondeada ó chata inclinada) para concluir que O. galea es un taxón independiente de O. quadratus.
Se registra por vez primera las especies O. bilobatus y O. orbiculatus en el PM.
Para la evaluación de las especies estudiadas y con base en los resultados morfométricos obtenidos, se propone que para reconocer a las especies de Ornithocercus tratadas en este estudio, es suficiente medir los siguientes seis caracteres: largo total de organismo, largo de la célula, área de la hipoteca, número de radios y dirección de R1 y R3, que además son caracteres que se pueden obtener sin dificultad de un mismo organismo. Por el contrario la aleta sulcal izquierda suele estar incompleta y con frecuencia falta la porción próxima al R3, debido a la forma en que estos organismos completan sus tecas al realizar la reproducción asexual (Reguera & González, 2001).
Existen diferencias morfológicas que nos permiten diferenciar a O. steini de O. thumii, a pesar de sus semejanzas morfológicas a primera vista. Los caracteres importantes para su diferenciación son: la forma de la aleta sulcal, el número de sus radios y lóbulos, así como la posición que ocupa el R1, el cual se prolonga en O. thumii hasta la costilla marginal, mientras que en O. steini sólo llega hasta el borde de la aleta. El AD mostró diferencias significativas entre ambos taxa para el conjunto de los caracteres evaluados, siendo los más importantes: el área de la epiteca y el ancho a nivel del cíngulum.
Agradecimientos
Los fondos para el desarrollo de esta investigación fueron obtenidos de los proyectos CONACYT 37560-V "Biología y Filogenia Molecular de Dinoflagelados del Pacifico Mexicano" y CONABIO: S151 y BA008. Se agradece la ayuda de Isabella Percopo para la obtención de literatura. Agradecemos los comentarios de Ismael Gárate Lizárraga y David Hernández Becerril para mejorar el manuscrito.
Referencias
ABÉ, T.H. 1967. The armored Dinoflagellata. II. Prorocentridae and Dinophysidae (B) - Dinophysis and its allied genera. Publish Seto marine Biology Laboratory 15: 37-68. [ Links ]
BALECH, E. 1988. Los Dinoflagelados del Atlántico Sudoccidental. Publicaciones Especiales Instituto Español de Oceanografía 1: 1-310. [ Links ]
BALECH, E. & H. FERRANDO. 1964. Fitoplancton Marino. Editorial Universitaria de Buenos Aires, Buenos Aires, 155 p. [ Links ]
FENSOME, R.A., F.J.R. TAYLOR, G. NORRIS, W.A.S. SARJEANT, D.I. WHARTON & G.L. WILLIAMS. 1993. A Classification of Living and Fossil Dinoflagellates. Micropaleontology, Special Publishers. No. 7, Sheridan Press, Hanover: 1-351. [ Links ]
GÓMEZ, F. 2005. A list of free-living dinoflagellate species in the world's oceans. Acta Botánica Croatica 64 (1): 129-212. [ Links ]
HAIR, J.F., R.E. ANDERSON, R.L. TATHAM & W.C. BLACK. 1998. Multivariate Data Analysis. 5th. Ed. Prentice Hall. Upper Saddle River, New Jersey, 730 p. [ Links ]
HALIM, Y. 1965. Microplancton des eaux egyptiennes. II.-Chrysomonadines; Ebriediens et Dinoflagellés noveaux ou d'intérêt biogéographique. Rapports et Procès-verbaux des Réunions de la Commission internationale de la Exploration Scientifique de la Mer Méditerranée 17: 495-502. [ Links ]
HERNÁNDEZ-BECERRIL, D.U. 1988a. Observaciones de algunos dinoflagelados (Dinophyceae) del Pacífico Mexicano con microscopios fotónico y electrónico de barrido. Investigación Pesquera 52 (4): 517-531. [ Links ]
HERNÁNDEZ-BECERRIL, D.U. 1988b. Planktonic dinoflagellates (except Ceratium and Protoperidinium) from the Gulf of California. Botánica Marina 1: 423-435. [ Links ]
HERNÁNDEZ-BECERRIL, D.U. 1988c. Especies de fitoplancton tropical de Pacífico mexicano II. Dinoflagelados y Cianobacterias. Revista Latinoamericana de Microbiología 30 (2): 187-196. [ Links ]
HERNÁNDEZ-BECERRIL, D.U., M.E. MEAVE DEL CASTILLO & C. FLORES-GRANADOS. 2003. Dinoflagelados del orden Dinophysiales en las Costas Mexicanas. In: Barreiro-Güemes M.T., M.E. Meave del Castillo y M. G. Figueroa-Torres (Eds.). Planctología Mexicana. Sociedad Mexicana de Planctología A. C. México, pp. 19-42. [ Links ]
HERNÁNDEZ-BECERRIL D.U., G. CEBALLOS-CORONA, K. ESQUEDA-LARA, M. TOVAR-SALAZAR & D. LEÓN-ÁLVAREZ. 2004. Dinoflagelados del Orden Dinophysiales (Dinophyta) de las costas del Pacifico Tropical Mexicano. Resúmenes XIII Reunión Nacional SOMPAC y VI Reunión Internacional de Planctología. Nuevo Vallarta, Nayarit, México, p. 90 [ Links ]
HERNÁNDEZ-ROSAS, A., M.E. MEAVE DEL CASTILLO & M. CASTILLO RIVERA. 2005. Morfometría de dinoflagelados del género Ornithocercus del Pacífico tropical mexicano. Resúmenes VII Congreso de Ficología de Latinoamérica y el Caribe y V Reunión Iberoamericana de Ficología. La Habana, Cuba, 57 p. [ Links ]
JÖRGENSEN, E. 1923. Mediterranean Dinophysiaceae. Report of the Danish Oceanographic Expedition Mediterranean 2 (Biological Journal) 2: 1-48. [ Links ]
KOFOID, C.A. 1907. Dinoflagellata of the San Diego region. III. Descriptions of new species. University of California Publications in Zoology 3 (13): 299-340. [ Links ]
KOFOID, C.A. & J.R. MICHENER. 1911. Reports on the scientific results of the expedition to the eastern tropical Pacific, in charge of Alexander Agassiz, by the U. S. Fish commission steamer "Albatross" from October 1904 to March 1905. L. M. Garrett, U. S. N., commanding 22. New genera and species of Dinoflagellates. Memoirs of the Museum of Comparative Zoology at Harvard College 54 (7): 267-302. [ Links ]
KOFOID, C.A. & T. SKOGSBERG. 1928. The Dinoflagellata: the Dinophysoidae. Memoirs of the Museum of Comparative Zoology at Harvard 51: 1-766. [ Links ]
LICEA, D.S., J.L. MORENO, H. SANTOYO & G. FIGUEROA. 1995. Dinoflagelados del Golfo de California. Universidad Autónoma de Baja California Sur. 165 p. [ Links ]
MARTÍNEZ-JASSO, E. & M.E. MEAVE DEL CASTILLO. 2002. Dinoflagelados del orden Dinophysiales Lindemann en el Pacífico Tropical Mexicano. Resúmenes VI Congreso Latinoamericano de Ficología y IV Reunión Iberoamericana de Ficología. Ponce, Puerto Rico, p. 33. [ Links ]
MARTÍNEZ-LÓPEZ, A. 1993. Efectos del evento "El Niño" 1982-83 en la estructura del fitoplancton en la costa occidental de Baja California Sur. Tesis de Maestría en Ciencias. Centro Interdisciplinario de Ciencias Marinas- IPN. La Paz, BCS, México, 83 p. [ Links ]
MARTÍNEZ-LÓPEZ, A. & G. VERDUGO DÍAZ. 2000. Composición y dinámica del fitoplancton en el BAC de Bahía Magdalena, Baja California Sur. Capitulo 9. In: Lluch-Belda, D., J. Elorduy-Gray, S.E. Lluch-Cota & G Ponce-Díaz (Eds.). BACs, Centro de actividad Biológica del Pacífico Mexicano. La Paz, México, pp: 125-142. [ Links ]
MATZENAUER, L. 1933. Die Dinoflagellaten des In Dischen Ozeans (mit Austnahme der Gatting Ceratium). Botanisches Archive 35: 437-510. [ Links ]
MEAVE DEL CASTILLO, M. E. 2006. Diatomeas (Bacillariophyta), Dinoflagelados (Dinophyta) y Silicoflagelados (Dictyochophyceae) marinos del Pacífico Mexicano con énfasis en la porción tropical. Reporte final del proyecto CONABIO BA008, México, 80 p. [ Links ]
MEAVE DEL CASTILLO, M. E. & D.U. HERNÁNDEZ-BECERRIL. 1998. Fitoplancton. In: M. Tapia García (Ed.). El Golfo de Tehuantepec: El Ecosistema y sus Recursos. Universidad Autónoma Metropolitana Iztapalapa, pp. 59-74. [ Links ]
MEAVE DEL CASTILLO, M.E. & M.E. ZAMUDIO-RESÉNDIZ. 2005. Biodiversidad de Dinoflagelados en el Pacífico Mexicano. Resúmenes del VII Congreso de Ficología de Latinoamérica y el Caribe y V Reunión Iberoamericana de Ficología. La Habana, Cuba, p.134. [ Links ]
NORRIS, D.R. & L.D. BERNER. 1970. Thecal morphology of selected species of Dinophysis (Dinoflagellata) from the Gulf of Mexico. Contributions on Marine Science 15: 145-192. [ Links ]
OKOLODKOV, Y.B. & I. GÁRATE-LIZÁRRAGA. 2006. An annotated checklist of Dinoflagellates (Dinophyceae) from the Mexican Pacific. Acta Botánica Mexicana 74: 1-154. [ Links ]
RAMPI, L. 1940. Ricerche sil microplancton del mare ligure 2. Le Tecatali e le Dinofisiali delle acque di Sanremo. Bollettino di Pesca, Piscicoltura e di Idrobiologial 16: 243-274. [ Links ]
RAMPI, L. 1950. Péridiniens rares ou nouveaux pour le Pacifique Sud-Équatorial. Bulletin de L'Institut Océanographique 974: 1-12. [ Links ]
REGUERA, B. & G.S. GONZÁLEZ. 2001. Small cell and intermediate cell formation in species of Dinophysis (Dinophyceae, Dinophysiales). Journal Phycology 37: 318-333. [ Links ]
SCHILLER, J. 1933. Dinoflagellatae (Peridineae) In: Rabenhorst, L. (Ed.) Kryptogamen-Flora von Deuschland, Österreich und der Schweiz, (Flagellatae), Akad. Verlagsges, Leipzig, Sect. 3, fasc. 1-3: 1-617. [ Links ]
SCHMIDT, A. 1874-1902. Atlas der Diatomaceen-Kunde in Verbindung mit den Herrn Grüdler, Granow, Janisch, Weissflog und Witt. Herausgegeben von Adolf Schmidt, Archdia conus in Aschersleben. Aschersleben, Leipzip, Achegel, 240 pls. [ Links ]
SCHÜTT, F. 1893. Das Pflanzenleben der Hochsee. Ergebn Plankton Expedition 1A, pp. 243-314. [ Links ]
SCHÜTT, F. 1896. Peridiniales. In: Engler & Prantl (Eds.) Die Natürlichen Pflanzefamilien, Leipzig, W. Engelmann 1(1b): 1-30. [ Links ]
SCHÜTT, F. 1900. Centrifugale und simultane Membranverdickungen. Jahrbucher fur Wissenschaftlichi Botanik 35 (3): 470-534. [ Links ]
SOURNIA, A. 1967. Contribution a la connaissance des Péridiniens Microplanctoniques du Canal de Mozambique. Bulletin Du Museum National D'Histoire Naturelle 2e Série 39 (2): 417-438. [ Links ]
SOURNIA, A. 1982. Is there a shade flora in the marine plankton? Journal of Plankton Research 4: 391-399. [ Links ]
SOURNIA, A. 1986. Atlas du Phytoplancton Marin. Volume I. Introduction, Cyanophycées, Dictyochophycées, Dinophycées, Raphidophycées. Éditions du Centre National de la Recherche Scientifique, Paris, 216 p. [ Links ]
STEIDINGER, K.A. & K.TANGEN.1997. Dinoflagellates. In: Tomas, C.R. (Ed.) Identifying Marine Phytoplankton. Academic Press, San Diego, pp. 387-584. [ Links ]
STEIN, F.R. 1883. Der organismus der arthrodelen Flagellaten II. Hälfte (Die Naturgeschichte der arthrodelen. Der organismus der Infusionthie nach eigenen Forschungen in systematischer Reihenfolge bearbeitet, Leipzig :1-30, pl. 1-42. [ Links ]
TAYLOR, F.J.R. 1976. Dinoflagellates from the International Indian Ocean Expedition. A report on material collected by the R.V. "Anton Bruun" 1963-1964. Biblotheca Botanica, Stuttgart 132: 1- 234, pl. 1-45. [ Links ]
WOOD, E.J.F. 1953. Dinoflagellates in the Australian region. Australian II. Recent collections. Division Fish and Oceanography CSIRO Australian 14: 1-55. [ Links ]
WOOD, E.J.F. 1954. Dinoflagellates of the Australian Region. Australian Journal of Marine & Freshwater Research 5 (2): 171-351. [ Links ]

