










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.17 no.2 Ciudad de México ago. 2007
Artículos
Morfología del sistema digestivo de la berrugata Menticirrhus saxatilis (Teleostei: Sciaenidae)
Morphology of the digestive system of Menticirrhus saxatilis (Teleostei: Sciaenidae)
Abraham Kobelkowsky y Orlando Pantoja-Escobar
Laboratorio de Peces. Departamento de Biología. Universidad Autónoma Metropolitana, Unidad Iztapalapa. Av. San Rafael Atlixco 186, Col. Vicentina, Iztapalapa, 09340, México D.F. E-mail: akd@xanum.uam.mx
Recibido: 6 de octubre de 2006
Aceptado: 4 de junio de 2007
Resumen
Como caracteres morfológicos relacionados con la bentofagia, Menticirrhus saxatilis muestra el tentáculo y los orificios mentonianos, el pliegue supraoral, la boca ventral. El esqueleto visceral presenta una notable anchura de los canales mandibular y preopercular del sistema sensorial de la línea lateral. Se reconoce una articulación especial del palatino con el paraesfenoides. Como caracteres sobresalientes de la musculatura visceral se encuentran: la sección tentacular del intermandibularis, la múltiple división del adductor mandibulae, la inserción del retractor dorsal en los costados de las dos primeras vértebras. Son notables los ligamentos rostrales y el que se origina del suborbital 3 y que separa la sección A3 del adductor mandibulae. La morfología de los componentes del tracto digestivo y la relación de la longitud tracto digestivo-longitud del pez, corresponde a la de un pez carnívoro. Se reconocen el ciego estomacal y siete ciegos pilóricos. La dentición de las mandíbulas y de los huesos faríngeos consiste de dientes cónicos puntiagudos y curvados, de varios tamaños, mientras que los dientes de los restantes arcos branquiales son pequeños y en placas.
Palabras clave: Branquiocráneo, musculatura visceral, tracto digestivo, dentición, bentofagia.
Abstract
As morphological characters related to bentophagy, Menticirrhus saxatilis shows the mental tentacle and pores, the supraoral fold, and the ventral position of mouth. The visceral skeleton shows notably wider the mandibular and preopercular lateral line channels; an additional condyle is found in the palatine for a special articulation with the parasphenoid. As conspicuous characters of the visceral musculature are: the tentacular section of intermandibularis, the multiple division of the adductor mandibulae, and of the insertion of the retractor dorsalis on the sides of the two first vertebrae. As notably ligaments are the rostrals and the one located between sections A3 and the rest of the adductor mandibulae. The morphology of the digestive tract is of typical carnivore. The intestine has the 70% of the standard length. This species has a big stomach caecum and seven pyloric caeca. Mandibular and pharyngeal dentition is of curved, conical and pointed teeth of various sizes, while teeth of the rest of branchial arches are fine, and on dental plates.
Key words: Branchiocranium, visceral musculature, digestive tract, dentition, bentophagy.
Introducción
De acuerdo con Gerking (1994) los teleósteos depredadores bentónicos son más variados morfológica y fisiológicamente que cualquier otro grupo trófico. En la comunidad de peces bentónicos de los sistemas estuarino lagunares del Golfo de México se reconocen dos formas del cuerpo, la aplanada y la pisciforme. La forma aplanada es en el sentido dorsoventral o deprimida como en las rayas, y la lateral o comprimido como en los lenguados.
Las especies de Sciaenidae tienen el cuerpo fusiforme, similar a los teleósteos nectónicos, sin embargo, numerosas especies son bentófagas y muestran la boca en posición ventral, con estructuras sensoriales que seleccionan el alimento, como tentáculos y orificios de la línea lateral.
La berrugata Menticirrhus saxatilis (Bloch & Schneider, 1801) se distribuye en el Atlántico Occidental, desde Massachussets hasta el sur de Florida, Estados Unidos de Norteamérica; y por el Golfo de México hasta Yucatán, México. Es una de las especies bentónicas del litoral y de los sistemas estuarino lagunares del Golfo de México (Reséndez-Medina & Kobelkowsky, 1991).
Entre los trabajos morfológicos de especies de Sciaenidae se encuentran el de Topp y Cole (1968) sobre el neurocráneo de Sciaenops; el de Castillo et al. (1988) sobre el sistema digestivo de Bairdiella ronchus; el de Ruiz et al. (2001) sobre el aparato bucofaríngeo de Micropogonias furnieri y el de Grubich (2005) sobre los mecanismos mandibulares de Pogonias cromis. Entre los trabajos que se han realizado sobre la morfología de especies de Menticirrhus, se encuentran el de Kong y Irachet (1995) sobre el neurocráneo. Jardim (1988) describe el neurocráneo y el esqueleto visceral de Menticirrhus littoralis y Menticirrhus americanus.
La dieta de las especies de Menticirrhus ha sido registrada por McMichel Jr. y Ross (1987), Bowman et al. (2000) y Chaves y Umbria (2003).
Debido a que M. saxatilis es parte de la comunidad de peces bentónicos de las lagunas costeras del Golfo de México, se tiene como objetivo la descripción de su sistema digestivo y de las estructuras sensoriales que participan en la selección del alimento.
Materiales y métodos
Se colectaron ejemplares de Menticirrhus saxatilis en las lagunas costeras de Pueblo Viejo, Tampamachoco y Alvarado, en el estado de Veracruz, mediante red de arrastre camaronera de prueba. Los ejemplares se fijaron en formaldehído al 10% y se conservaron en alcohol etílico al 70%.
Se realizó el análisis anatómico de 10 ejemplares cuya longitud patrón varió de 158 a 210 mm con un promedio de 186.5 mm. Se registró el número de branquiespinas del primer arco branquial. Se contaron los ciegos pilóricos y se registró la longitud intestinal, excluyendo al estómago. La longitud intestinal se relacionó con la longitud patrón del pez.
Se observó y describió la morfología externa de la boca y de sus estructuras asociadas. Se retiró la piel de la región cefálica con el objeto de exponer el neurocráneo y el esqueleto visceral. Se analizó y describió el arco mandibular, así como también su dentición. Se extrajeron el globo ocular y la serie de huesos suborbitales, para exponer los músculos viscerales más superficiales.
Se retiraron el hueso nasal y el órgano del olfato para exponer el conjunto de ligamentos y cartílagos rostrales. Se analizó y retiró el músculo aductor mandibular, para evidenciar los músculos más internos y las series de huesos del paladar y la suspensión mandibular. Se retiraron los huesos del arco mandibular, la serie palatina, la suspensión mandibular y la serie opercular, para exponer los aparatos hioideo y branquial y su musculatura. Se desprendieron las laminillas branquiales con el objeto de exponer los arcos branquiales.
Se retiraron los músculos intrínsecos de la cintura escapular y se observaron los músculos que la relacionan con los arcos branquiales y con el neurocráneo. A continuación se cortó el ligamento de Baudelot y se retiró la cintura escapular, para exponer los músculos que relacionan las vértebras con los arcos branquiales. Se retiró el aparato branquial y se expuso su esqueleto para contar y describir las branquiespinas y la dentición.
Se retiró la piel de la región entre la cintura escapular y el nivel del ano. Se extrajeron los músculos axiales de dicha región, así como también las costillas dorsales (epipleurales) y ventrales (pleurales), con el objeto de exponer la cavidad visceral. Se describió la organografía de esta cavidad. Se retiraron las gónadas y la vejiga gaseosa o natatoria. Mediante un bisturí se seccionó transversalmente el esófago y las venas hepáticas, así como también el recto, con el propósito de extraer el tracto digestivo con el hígado. Se describieron éstos, y se registraron las medidas de longitud de los componentes del tracto digestivo y el número de ciegos pilóricos.
La terminología del endoesqueleto siguió el criterio de Gregory (1959) y la de la musculatura el criterio de Winterbottom (1974).
Las ilustraciones se realizaron mediante una camera lucida acoplada a un microscopio estereoscópico Wild M3Z.
Resultados
Morfología externa. La berrugata Menticirrhus saxatilis tiene la boca relativamente pequeña, en posición ventral y sub-terminal (Fig. 1 a). Sobre el labio superior se forma un pliegue rostral con dos pares de lóbulos. Se reconocen poros rostrales anteriores y sobre el pliegue rostral.

En la mandíbula inferior se localizan dos pares de orificios mentales y un tentáculo mental impar (Fig. 1 b). Los orificios mentales se comunican con las ramas mandibulares del sistema sensorial de la línea lateral. Se observa la conexión de dos secciones del músculo intermandibularis con la base del tentáculo mental (Fig. 1 c).
Esqueleto visceral. El arco mandibular incluye a los huesos premaxilar, maxilar, dentario, articular y angular (Figs. 2 b, 3a). El paladar está formado por el palatino, mesopterigoides y ectopterigoides (Fig. 3 d). El palatino muestra un cóndilo adicional que mediante ligamentos se articula con otro del paraesfenoides del neurocráneo. La suspensión mandibular está formada por el hiomandibular, metapterigoides, simpléctico y cuadrado (Fig. 3 d).
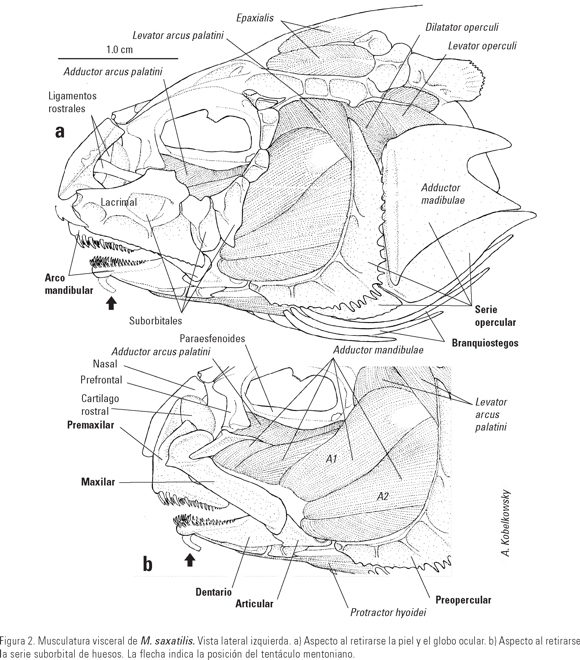

El aparato hioideo está formado por el basihial, hipohial superior, hipohial inferior, ceratohial, epihial, interhial, urohial y siete branquiostegos (Fig. 3 e). La unión entre hipohial inferior y ceratohial, y entre este y el epihial es por sutura. La longitud de los branquiostegos aumenta del primero al último, siendo el primero notablemente delgado y los branquiostegos 2 y 3 son angulosos.
El aparato branquial (Fig. 4 c) está constituido por tres basibranquiales, tres hipobranquiales, cuatro ceratobranquiales, el faríngeo inferior (ceratobranquial 5), cuatro epibranquiales, y cuatro faringobranquiales. Los surcos para las arterias branquiales se encuentran en todos los hipo, cerato y epibranquiales de los cuatro primeros arcos.

En vista dorsal el basibranquial 2 tiene forma de matraz, mientras que el 3 es lanceolado. La longitud de los hipobranquiales disminuye del primero al tercero. Los ceratobranquiales 1 a 4 son similares entre sí, mientras que el ceratobranquial 5 (Fig. 4 d) o hueso faríngeo inferior es triangular, con un proceso posterior largo, y lleva los dientes faríngeos inferiores. Cada uno de los epibranquiales forma un proceso dorsal para la inserción de los músculos levatores externi y levator posterior (Fig. 4 b). El faringobranquial 1 es delgado, se fija a la región ótica del neurocráneo, suspendiendo al aparato branquial. Los faringobranquiales 2 a 4 llevan los dientes faríngeos superiores. El faringobranquial 2 tiene la porción anterior plana y la posterior lleva dientes. El faringobranquial 3 es trapezoidal y su superficie ventral está tapizada por dientes. El faringobranquial 4 es ovalado y lleva un conjunto de dientes.
Dentición. M. saxatilis lleva dientes en los arcos mandibular y branquiales, y carece de dientes en el paladar (vomer, palatino, ectopterigoides) y la lengua (basihial).
Los dientes del arco mandibular (Fig. a, b, c) son cónicos y curvados hacia atrás y hacia adentro. Los correspondientes a los premaxilares están arreglados en una hilera externa de elementos grandes y varias hileras de dientes más pequeños. Los dientes del dentario forman un conjunto de elementos de mediano tamaño, delgados y curvados hacia atrás.
En los arcos branquiales los dientes son numerosos y pequeños. Los dientes del primer arco branquial se encuentran en un promedio de cuatro branquiespinas y en 8 placas dentarias. Los dientes de los arcos branquiales 2 a 4 son pequeños y se encuentran solamente en placas dentarias. Los dientes faríngeos superiores se localizan en los huesos faringobranquiales 2 a 4 (Fig. 4 e). Los dientes faríngeos inferiores del borde interno son grandes, mientras que los restantes decrecen en tamaño hacia el borde externo del ceratobranquial 5 o hueso faríngeo inferior (Fig. 4 d).
Musculatura visceral. De la musculatura mandibular sobresalen los músculos intermandibularis, adductor mandibulae, adductor arcus palatini y levator arcus palatini. El intermandibularis (Fig. 1 c) se divide en dos secciones, de las que la ventral se modifica en dos músculos de posición oblicua, que mueven el tentáculo mentoniano. El adductor mandibulae (Fig. 2) está dividido en los paquetes A1, A2, A3 y Aw. El A3 se subdivide en tres secciones, de las cuales las dos más anteriores se insertan en la superficie posterior de la cabeza del maxilar. La sección más anterior se origina en la concavidad del palatino. Entre los paquetes A2 y A3 se forma una fascia que se extiende desde el metapterigoides hasta el proceso subocular del hueso suborbital 3. El adductor arcus palatini (Fig. 2) es extenso, se origina en el parasfenoides y se inserta sobre parte de la superficie dorsal del mesopterigoides y en la superficie ventral de la lámina anterior del metapterigoides. El levator arcus palatini (Fig. 2) se origina en el esfenótico y se inserta en la porción superior del hiomandibular.
De la musculatura hioidea sobresalen los músculos protractor hyoidei y sternohyoideus. El protractor hyoidei (Fig. 1 c) se origina en la superficie externa del aparato hioideo y se inserta mediante un ligamento a la parte interna de la sínfisis mandibular. El sternohyoideus (Fig. 4 b) se origina en el cleitrum y se inserta en ambas caras del urohial.
De los músculos dorsales de los arcos branquiales sobresalen los cuatro músculos levatores externi, el levator posterior y el retractor dorsalis. Los levatores externi (Fig. 4 b) se originan del pterótico y aumentan su longitud del primero al último. El levator posterior (Fig. 4 b) se origina de la cápsula ótica y se inserta en el epibranquial 4. El músculo retractor dorsalis está formado por varios paquetes que se originan de las partes laterales y ventrales de las dos primeras vértebras y se insertan sobre los faringobranquiales.
De los músculos ventrales a los arcos branquiales sobresalen los músculos pharyngoclavicularis externus e internus, transversi ventrales anterior y posterior, y el rectus communis (Fig. 4 b). El músculo pharyngoclavicularis internus está formado por varios paquetes que, se originan de la porción media de la superficie anterior del cleitrum y se insertan en el hueso faríngeo inferior. El músculo pharyngoclavicularis externus se origina cerca del borde de la porción inferior del cleitrum y se inserta en el faríngeo inferior. El músculo transversus ventralis anterior pasa del ceratobranquial 4 de un lado al del otro. El transversui ventralis posterior pasa del ceratobranquial 5 de un lado al del otro. El músculo rectus communis se origina en el ceratobranquial 5 y se inserta en el borde superior del urohial (Fig. 4 b).
Tracto digestivo. La porción anteroventral de la cavidad visceral está ocupada por el tracto digestivo y el hígado, mientras que el riñón, las gónadas, el bazo y la reducida vejiga gaseosa, ocupan el resto (Fig. 5 a).
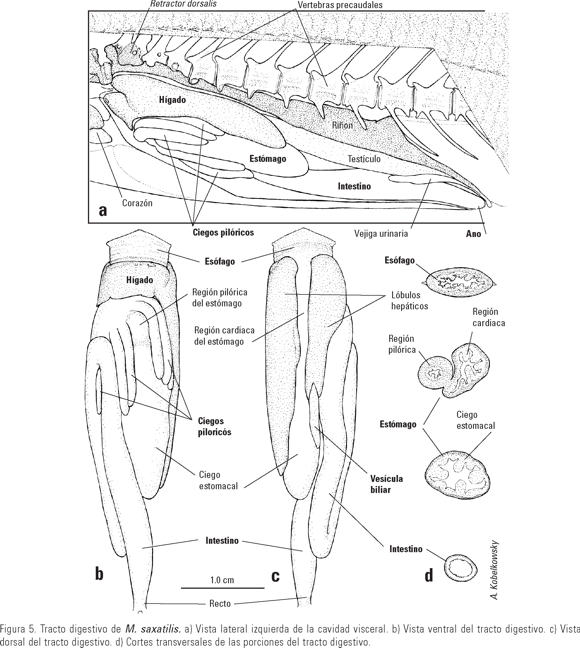
El hígado (Fig. 5) es alargado y con dos lóbulos que cubren los costados del estómago, siendo el lóbulo izquierdo el más extenso. La porción media del hígado es ventral al esófago y muestra en cada lado una marcada escotadura (Fig. 5 a) por la que pasa el ligamento lateral del esófago. La vesícula biliar es alargada y se ubica en el extremo posterior del lóbulo hepático derecho, sobre el ciego estomacal (Fig. 5 b). Dorsalmente al estómago, y entre ambos lóbulos hepáticos se localiza la vejiga gaseosa, la cual tiene el diámetro extremadamente reducido.
El esófago (Fig. 5) es de sección aplanada y muestra una primera porción amplia y otra posterior más angosta. Sus bordes anteriores son agudos, correspondiendo en forma con los bordes posteriores del aparato branquial. De sus bordes laterales parten dos ligamentos laminares que lo unen con la pared anterior de la cavidad visceral. Su superficie interna consiste de surcos longitudinales someros, con abundantes digitaciones finas.
El estómago (Fig. 5) es largo y ancho y está formado por una porción cardiaca de mediano tamaño, una porción pilórica notablemente reducida; desde estas partes se extiende hacia atrás un amplio ciego estomacal. Los surcos de la superficie interna del estómago son continuación de los del esófago, sin embargo, no llevan las digitaciones finas. La porción pilórica lleva un reducido número de surcos y un amplio esfínter.
El intestino (Fig. 5) es relativamente corto y forma dos asas longitudinales. Su longitud varió de 126 a 153 mm, con un promedio de 135 mm. Su longitud corresponde a cerca del 70% de la longitud patrón del pez. Su superficie interna está formada por pequeñas papilas y no muestra surcos. De su área cercana al píloro se desprenden siete ciegos pilóricos largos, de los cuales el más posterior se ubica bajo el asa intestinal anterior. Un ejemplar registró ocho ciegos. La longitud promedio de los ciegos pilóricos varió de 16 a 23 mm y su superficie interna es similar a la del intestino.
Discusión
Aunque el aspecto fusiforme de Menticirrhus saxatilis es similar al de las especies nectónicas de teleósteos, la posición ventral de la boca y la presencia de estructuras sensoriales, tales como el tentáculo mentoniano y los orificios rostrales, permiten reconocer a esta especie como bentófaga.
La organización del esqueleto visceral de M. saxatilis corresponde al patrón general descrito por Gregory (1933) para los Perciformes. Sin embargo, se reconocen características exclusivas de la familia Sciaenidae, como el notable ensanchamiento de los canales del sistema sensorial de la línea lateral en la mandíbula inferior y en el preopercular, aspectos que son descritos por Topp y Cole (1968) en especies de Sciaenops. En el presente trabajo se reconoce una comunicación directa de los orificios mentonianos con la rama mandibular de dicho sistema sensorial.
En términos generales la morfología bucofaríngea de M. saxatilis es similar as la de Micropogonias furnieri, descrita por Ruiz et al. (2001), sin embargo, en el presente estudio se reconoce como un rasgo especial la articulación adicional entre el palatino y el paraesfenoides. Los cóndilos del paraesfenoides no son descritos por Jardim (1988) en su estudio del neurocráneo de especies de Menticirrhus de Brasil.
En términos generales la organización de la musculatura visceral de M. saxatilis corresponde al patrón general de los teleósteos, que describe Winterbottom (1974), sin embargo, sobresalen algunas variaciones.
En el presente trabajo se describe por primera vez en la literatura la subdivisión del intermandibularis en dos secciones, especializándose una en el movimiento del tentáculo mentoniano. Esta situación morfológica es diferente a la de los tentáculos mandibulares del bagre Ameiurus nebulosus (LeSueur, 1819), que Winterbottom (1974) ilustra en contacto con el músculo protractor hyoidei.
Considerando que el adductor mandibulae presenta en los teleósteos una gran diversidad morfológica, en el presente estudio se remarca la división de la sección A3 en varios paquetes musculares con inserción en el maxilar. Asimismo, resalta la separación entre las secciones A1 y A3, mediante una fascia que se extiende entre el suborbital 3 y el metapterigoides.
Mientras que en otros teleósteos el retractor dorsal se origina de la parte ventral de las primeras vértebras, sobresale la situación en M. saxatilis en la que se origina también de las partes laterales de las dos primeras vértebras.
La organización de los dientes de M. saxatilis es similar a la de los teleósteos carnívoros, en los cuales los dientes son numerosos, puntiagudos y curvados hacia atrás. Esto corresponde con la dieta registrada de esta especie por Bowman et al. (2000) en la parte norte del Atlántico y de M. americanus (Linnaeus, 1758) por Chaves y Umbria (2003) en el Atlántico Sur, y que consiste de crustáceos peracáridos y decápodos, sifones de bivalvos, poliquetos y ocasionalmente peces.
La dentición mandibular y branquial no muestra alguna especialización en la durofagia o ingestión de animales con concha dura, como describe Grubich (2005) en el esciénido Pogonias cromis (Linnaeus, 1766).
La organografía de la cavidad visceral de M. saxatilis corresponde al patrón general de los Sciaenidae, como el descrito por Kobelkowsky (1999) en los esciénidos de los géneros Bairdiella y Stellifer. Sin embargo, la especie en estudio muestra como una adaptación a la vida bentónica, la notable reducción de tamaño de la vejiga gaseosa, mientras que en numerosas especies de Sciaenidae, este órgano es grande y se supone relacionado con la emisión de sonidos y posiblemente la recepción del mismo, como indica Tavolga (1971).
La morfología del tracto digestivo de M. saxatilis corresponde a la que (Nikolsky, 1963) describe para los teleósteos carnívoros, consistente en tener el estómago relativamente grande y la longitud del tracto digestivo menor a la longitud patrón del pez. Asimismo, es similar a la que describen Castillo et al. (1988) en Bairdiella ronchus (Cuvier, 1830), la cual es una especies de Sciaenidae reconocida como carnívora de tipo bentófaga. Con frecuencia los carnívoros muestran un ciego estomacal muy desarrollado, para contener los animales ingeridos y evitar su tránsito directo al intestino, mientras que los teleósteos herbívoros muestran el intestino largo, debido a que algunos componentes de su dieta son de lenta digestión, requiriendo más tiempo.
De acuerdo con lo anteriormente anotado, se concluye que M. saxatilis es una especie morfológicamente adaptada a una condición bentófaga, considerando la posición ventral de la boca y la presencia de estructuras que probablemente identifican el alimento del fondo, como los orificios rostrales y mandibulares y el tentáculo mentoniano. El carácter carnívoro de esta especie se reconoce por el aspecto puntiagudo de los dientes y por la menor longitud del tracto digestivo con relación a la longitud del cuerpo. Asimismo, se concluye que la organización del esqueleto y la musculatura viscerales corresponde al patrón morfológico del orden Perciformes, sin embargo, el presente trabajo es el primero en reconocer la articulación especial entre palatino y paraesfenoides.
Referencias
BOWMAN R. E., C.E. STILLWELL, W.L.MICHAELS, & M.D. GROSSLEIN. 2000. Food of Northwest Atlantic fishes and two common species of squid. NOAA Technical memorandum NMFS-NE155. 137 p. [ Links ]
CASTILLO, R.M.A., M.A. PÉREZ H. & A. KOBELKOWSKY D. 1988. Estudio de la biología de Bairdiella ronchus (Pisces: Sciaenidae) de la Laguna de Tampamachoco, Veracruz, México. Universidad y Ciencia 5(10):69-84. [ Links ]
CHAVES P.T.C. & S.C.UMBRIA. 2003. Changes in the diet composition of transitory fishes in coastal systems, estuary and continental shelf. Brazilian Archives of Biology and Technology 46(1):41-46. [ Links ]
GERKING, S.D. 1994. Feeding Ecology of fish. Academic Press, San Diego. 416 p. [ Links ]
GREGORY, W.K. 1933. Fish skulls. A study of the evolution of natural mechanisms. Eric Lundberg, Laurel, Florida. 481 p. [ Links ]
GRUBICH, J.R. 2005. Disparity between feeding performance and predicted muscle strength in the pharyngeal musculature of black drum, Pogonias cromis (Sciaenidae). Environmental Biology of Fishes 74:261-272. [ Links ]
JARDIM, L.F.A. 1988. Osteologia do sincránio das especies de Menticirrhus Gill, 1861 da costa sul do Brasil (Perciformes, Sciaenidae). Atlântica 10(1):41-74 [ Links ]
KOBELKOWSKY, D.A. 1999. El sistema urogenital de los géneros Bairdiella y Stellifer (Pisces: Sciaenidae). Revista de la Sociedad Mexicana de Historia Natural 49:153-162. [ Links ]
KONG, U.I. & I.P. IRACHET. 1995. Osteología comparada del cráneo de ocho especies de peces Sciaenidae de Chile. Estudios Oceanológicos 14:5-16. [ Links ]
MCMICHAEL JR. R.H. & S. ROSS. 1987. The relative abundance and feeding habits of juvenile kingfish (Sciaenidae:Menticirrhus), in a Gulf of Mexico surf zone. Northeast Gulf Science 9(2):109-123. [ Links ]
NIKOLSKY, G.V. 1963. The ecology of fishes. Academic Press, London. 352 p. [ Links ]
RESÉNDEZ- MEDINA, A. & A. KOBELKOWSKY D. 1991. Ictiofauna de los sistemas lagunares costeros del Golfo de México, México. Universidad y Ciencia 8(15):91-110. [ Links ]
RUIZ, L.J., A. PRIETO A. & M. LEMUS. 2001. Morfología bucofaríngea y hábitos alimentarios de Micropogonias furnieri (Pisces: Sciaenidae) en la costa norte del Estado de Sucre, Venezuela. Revista de Biología Tropical 49(3-4):903-913. [ Links ]
TAVOLGA, W.N. 1971. Sound production and detection. In: Hoar, W.S. & D.J. Randall (Eds.) Fish Physiology. Academic Press. 5:135-205. [ Links ]
TOPP, R.W. & C.F. COLE. 1968. An osteological study of the sciaenid genus, Sciaenops Gill (Teleostei, Sciaenidae). Bulletin of marine Science 902-945. [ Links ]
WINTERBOTTOM, R. 1974. A descriptive synonymy of the striated muscles of the Teleostei. Proceeding of the Academy of Natural Sciences of Philadelphia 125 (12):225-317. [ Links ]

