










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.17 no.1 Ciudad de México Abr. 2007
Artículos
Distribución latitudinal de las fases reproductivas de Lithothrix aspergillum (Corallinales, Rhodophyta) en la costa del Pacífico de México
Latitudinal distribution of the Lithothrix aspergillum (Corallinales, Rhodophyta) of the Mexican Pacific coast
Raúl Aguilar-Rosas1 Luis Ernesto Aguilar-Rosas2 Catalina Mendoza-González3 y Luz Elena Mateo-Cid3
1 Facultad de Ciencias Marinas, Universidad Autónoma de Baja California, km 107 Carretera Tijuana Ensenada, Ensenada, Baja California. C. P. 22830. E-mail: raguilar@uabc.mx.
2 Instituto de Investigaciones Oceanológicas, Universidad Autónoma de Baja California, km 107 Carretera Tijuana Ensenada, Ensenada, Baja California. C. P. 22830.
3 Departamento de Botánica, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional, Prolongación Carpio y Plan de Ayala, 11340 México, D. F.
Recibido: 12 de febrero de 2006
Aceptado: 08 de diciembre de 2006
Resumen
Se llevaron a cabo recolectas de Lithothrix aspergillum J. E. Gray (Corallinales, Rhodophyta) en diversas localidades en la costa occidental de Baja California, durante 1996 a 2002, y una revisión exhaustiva de ejemplares depositados en herbarios nacionales y extranjeros, con el fin de llegar a determinar la presencia y distribución latitudinal de las fases reproductivas de L. aspergillum en la costa del Pacífico de México. El análisis de la información reveló que los talos tetrasporangiales se encuentran presentes en todo el intervalo de distribución conocido para L. aspergillum en la costa occidental de la Península de Baja California; los talos bisporangiales están restringidos a la parte noroccidental de la Península; mientras que los escasos talos gametangiales (femeninos y masculinos) son comunes en la parte sur de la Península. En general se observó que los talos bisporangiales fueron los más arborescentes y de mayor talla (9.4 ± 1.4 cm promedio); los talos tetrasporangiales los más compactos, ramificados y de menor talla que los bisporangiales (4.4 ± 1.2 cm), mientras que los talos gametangiales se caracterizaron por ser los más pequeños (3.5 ± 0.8 cm). Se describen los caracteres morfológicos y reproductivos de cada una de las fases reproductivas.
Palabras clave: Corallinales, distribución latitudinal, Lithothrix aspergillum, morfología, Pacifico de México, reproducción.
Abstract
Population of Lithothrix aspergillum J. E. Gray (Corallinales, Rhodophyta) was collected at several sites on the west coast of Baja California during 1996 to 2002 and a through revision was made of specimens deposited in herbaria in Mexico and California in order to determine the occurrence and latitudinal distribution of the reproductive phases of L. aspergillum on the Mexican Pacific coast. Analysis of the information reveled that tetrasporangial thalli are present along the all range of distribution know for L. aspergillum in the west coast of the Baja California peninsula; the bisporangial thalli are confined in the northwest part of the peninsula; meanwhile the scarce gametangial thalli (male and female) are commons in the south part of the peninsula. In general, bisporangial thalli are arborescent and large, with a mean size of 9.4 ± 1.4 cm; tetrasporangial thalli are more compact and branched, and lower in size than tetrasporangial thalli with a mean size of 4.4 ± 1.2 cm; meanwhile, gametangial thalli are characterized by the small size, with a mean size of 3.5 ± 0.8 cm. The morphology and reproductive characters of each reproductive phases are described.
Key words: Corallinales, latitudinal distribution, Lithothrix aspergillum, morphology, Pacific of Mexico, reproduction.
Introducción
Lithothrix aspergillum J. E. Gray es una alga coralina articulada que se encuentra ampliamente distribuida en la costa del Pacífico de norte América desde Alaska hasta Isla Magdalena, Baja California Sur, México (Scagel et al., 1989). En todo el intervalo de distribución geográfica de L. aspergillum, se ha determinado que sólo en el sur de California, E.U.A. (Orange County) las poblaciones se mantienen a través de talos bisporangiales, tetrasporangiales y gametangiales (Ganesan y Desikachary, 1970; Abbott y Hollenberg, 1976; Tyrrell y Johansen, 1995; Pearson y Murray, 1997), y en contraste las poblaciones localizadas al norte (Columbia Británica) y sur (Baja California Sur, México) de esta área, se reproducen solo por medio de talos bisporangiales (Tyrrell y Johansen, 1995; Pearson y Murray, 1997). Tyrrell y Johansen (1995), basaron su estudio en el análisis de los ejemplares recolectados por E. Y. Dawson en la costa occidental de la Península de Baja California, y determinaron que solo talos bisporangiales o estériles se presentan en la costa Pacífico de México, argumentando que este comportamiento reproductivo se debe a la presencia de aguas frías producidas por el fenómeno de surgencias, cuya temperatura superficial del agua de mar se encuentra por debajo de los 14 °C (Dawson, 1945,1951, 1954). La posibilidad de que se desarrollen talos tetrasporangiales y gametangiales solo al sur de California, lo atribuyen a que en cierta época del año, la temperatura promedio del agua de mar se presenta por arriba de los 15-16 °C (Tyrrell y Johansen, 1995).
Estudios previos realizados en la costa Pacífico de México, han permitido constatar la presencia de talos bisporangiales y tetrasporangiales, y una marcada ausencia de talos gametangiales (Dawson, 1953; Mendoza-González y Mateo-Cid, 1985; Aguilar-Rosas et al., 1990; Mendoza-González y Mateo-Cid, 1992; Mateo-Cid y Mendoza-González, 1994); sin embargo, partiendo del hecho de que las surgencias muestran una distribución discontinua a lo largo de la costa occidental de la Península de Baja California, se espera encontrar todo tipo de fases reproductivas de L. aspergillum tanto en ambientes fríos como en lugares cálidos.
Con base en lo antes expuesto, este estudio tiene como objetivos: a) describir los caracteres morfológicos y reproductivos de L. aspergillum en la costa Pacífico de México y b) determinar, a través de una revisión exhaustiva de especimenes recolectados durante 1996-2002 y materiales depositados en herbarios, la presencia y distribución latitudinal de las fases reproductivas en la costa Pacífico de México.
Materiales y métodos
Las observaciones están basadas en (1) especimenes conservados en formaldehído al 4 % en agua de mar, recolectados entre octubre de 1996 y noviembre de 2002 en diversas localidades de la costa occidental de Baja California (Fig. 1); y (2) especimenes secos depositados en los Herbarios de la Facultad de Ciencias Marinas (CMMEX), Herbario de la Escuela Nacional de Ciencias Biológicas (ENCB), Herbario del Museo de Historia Natural de Los Ángeles (LAM) y el Herbario de la Universidad de California (UC), en Berkeley. Para llevar a cabo la descripción de las plantas, se determinó la talla y se efectuaron cortes transversales de las estructuras vegetativas y reproductivas. Los especimenes fueron descalcificados con 0.6M HNO3 y deshidratados con alcohol etílico. Segmentos pequeños fueron embebidos en parafina, cortados en secciones de 9-10 m de grosor con ayuda de un microtomo de mano, fijos sobre portaobjetos con pegamento Ruyter's (Martoja y Martoja-Pierson, 1970) y teñidos con anilina azul y eosina-hematoxilina. Las fotografías se tomaron con una cámara digital (Sony DSC-S85) en un microscopio Zeiss (Axioscop 40). Las preparaciones permanentes y el material recolectado quedaron depositado en el Herbario CMMEX. La identificación de las fases reproductivas se llevó a cabo utilizando los trabajos de Abbott y Hollenberg (1976), y Ganesan y Desikachary (1970). Los acrónimos de los herbarios siguen a Holmgren et al. (1990).

Resultados
Lithothrix aspergillum J. E. Gray (1867): 33
Localidad tipo: Isla Vancouver, Columbia Británica, Canadá.
Estructura vegetativa: Talos erectos, cuyos ejes nacen de una porción basal costrosa extendida sobre el sustrato, de color rosa-brillante a rojo verdoso, de (3.0) 4.0-8.5 (12.0) cm de alto (Fig. 2-5). Intergenículas basales cilíndricas de (0.4) 0.5-0.85 (1.0) mm de diámetro y (0.3) 0.4-1.0 mm de longitud; las porciones medias con intergenículas cilíndricas a comprimidas de (0.3) 0.4-0.7 mm de ancho y las superiores comprimidas de (200) 240-270 (400) μm de ancho; ramificación primaria dicotómica, la secundaria verticilar, radial o irregular; ápices obtusos. En sección longitudinal médula de la intergenícula unizonal, las células son de 480-580 μm de largo y (6) 8-10 (12.5) μm de diámetro; corteza formada por 2 capas de células, las más externas de (3.5) 4.0-6.0 μm longitud y (8) 9-10 (12) μm ancho, la interna de (8.0) 9.0-10 μm ancho y (15) 16-19 (21) μm longitud, epitalo de una sola capa de células de (9) 10-13 (14) μm ancho y (6) 7-8.5 (9.5) μm alto (Fig. 6-8). Genícula unizonal de (0.4) 0.5-0.8 (1.0) mm ancho (Fig. 7).
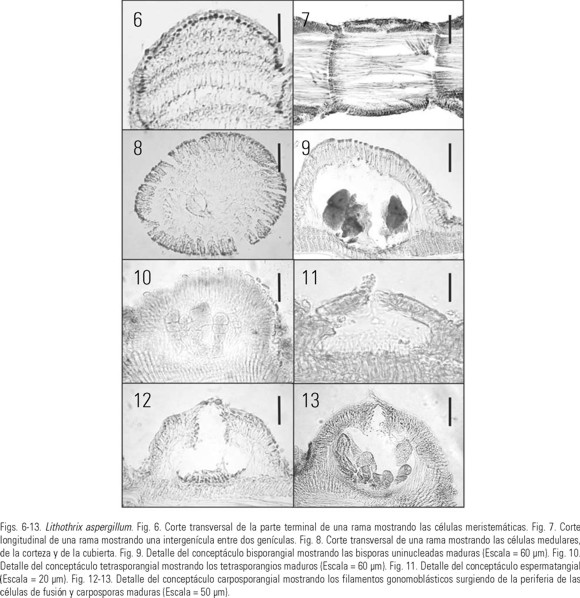
Estructuras reproductivas: Talos bisporangiales de 9.4 ± 1.4 cm de alto (n 95) (Fig. 2), con conceptáculos extremadamente prominentes sobre los ejes axiales; cámaras bisporangiales de (400) 430-480 (510) μm diámetro y (180) 190-240 (250) μm de longitud; bisporangios de (30) 36-47 (51) μm diámetro y (72) 78-95 (102) μm longitud (Fig. 9). Talos tetrasporangiales de 4.4 ± 1.2 cm de alto (n 93) (Fig. 3), con conceptáculos prominentes sobre los ejes axiales y ramas laterales; cámaras tetrasporangiales de (280) 300-320 (350) μm diámetro y (200) 220-240 (260) μm de alto; tetrasporangios zonados de (36) 45-54 (58) μm diámetro y (96) 120-168 (175) μm longitud (Fig. 10). Talos espermatangiales de 2.2 ± 0.4 cm (n 10) (Fig. 5), con conceptáculos ligeramente elevados o inconspicuos, de forma piramidal sobre las ramas laterales cortas; cámaras espermatangiales miden de 80-90 (100) μm diámetro y 40-44 (50) μm de alto (Fig 11). Talos carposporangiales de 3.0 ± 0.8 cm (n 12) (Fig. 4), con conceptáculos globulares y extremadamente prominentes sobre las ramas laterales; camaras carposporangiales de 180-280 (300) μm de diámetro (Fig. 6) y 110-160 (180) μm de alto (Fig. 12-13).
Habitat: Sobre rocas, epífito o epizoico, en pozas intermareales. En la zona submareal entre 2-10 m de profundidad.
Material examinado. Baja California: Islas Coronado, 20.VI.1947, EYD 4255 (LAM 19407); 26.XI.1949, PCS 4878 (UC 974664); Playa Rosarito, 5.III.1947, COOPER 544 (LAM 14517); Punta Popotla (Estudios Fox), 19.XI.1998, RAR 3199 (CMMEX 4421; 3 km al norte de Punta Descanso, 8.IV.1945, EYD 78-45 (UC 694016); 8 km al sur De Punta Descanso, 14.I.1946, EYD 209 (LAM 4252); Club Marena, 26.IX.1996, RAR 3192 (CMMEX 4413); 4.IV.1997, RAR 3193 (CMMEX 4414); Rancho Don Pancho, 9.IV.1997, RAR 3194 (CMMEX 4418); Raul's, 9.II.1995, BC-95-150/01 (ENCB 12181); 3.XII.2002, RAR 3204 (CMMEX 4426); Medio Camino, 7.V.1980, IPR 88 (CMMEX 2363); Punta Mezquitito, 7.I.2000, RAR 3203 (CMMEX 4425); Bajamar, 1.III.1999, RAR 3201 (CMMEX 4423); Barco Hundido, 26.VI.1982, LA 480 (CMMEX 2447); Punta San Miguel (Boca del río), 5.XI.1998, RAR 3198 (CMMEX 4420); Villa de las Rosas, 13.XI.1981, MAR s/n (CMMEX 4408); 21.IV.1999, RAR 3200 (CMMEX 4422); Punta Morro, 9.IV.1978, MAR 4 (CMMEX 4405); 9.IV.1995, BC-96-177/01 (ENCB 12268); .2.VIII.1996, BC-96-248/01 (ENCB 13029); Hotel Marina Coral (= Playitas), 9.I.1996, LEAR s/n (CMMEX 4411); 19.I.1996, RAR 3191 (CMMEX 4410); 3.VIII.1996, BC-96-251/01 (ENCB 12267); El Mosquito, 1.VIII.1987, s/n (ENCB 13027); Islas Todos Santos, 26.II.1949, PCS 4788 (UC 974776); Rancho Packard, 19.V.1983, s/n (ENCB 4639); 19.IX.1998, RAR 3197 (CMMEX 4419); Bahía de Todos Santos, 5.IX.1948, EYD 5189 (UC 949733); 6.IX.1948, EYD 5272 (LAM 36627); La Bufadora, 17.IX.1987, s/n (ENCB 7980); Punta San José, 4.XII.1979, MAR 46 (CMMEX 4341); Punta San Isidro, 21.VI.1994, BC94-24/02 (ENCB 12182); Cabo Colonet, 13.I.1946, EYD 93 (LAM 4248); Isla San Martín, 9.XI.1950, PCS 6601 (UC 974695); Península de San Quintín, 3.I.1950, EYD 8758 (UC 925586); Punta Baja, 9.IV.1946, EYD 1284 (LAM 4282); Punta Santa Rosalillita, 9.X.1946, EYD 2766 (LAM 19417); 10.X.1946, EYD 2853 (UC 756272); Isla San Benito, 19.VIII.1946, EYD 1528-46 (LAM 19411); 5.XI.1950, PCS 6463 (UC 974545); Baja California Sur: Isla de Cedros, 19.IV.1951, EYD 9857 (LAM 57243); 30.X.1951, EYD 10581 (UC 980190); Bahía Vizcaíno (Desembarcadero de Miller's), 12.IV.1946, EYD 1388 (LAM 4292); 11.X.1946, EYD 2944 (UC 756273); Punta Eugenia, 1.XI.1951, EYD 10407 (UC 940203); Bahía Tortugas, 7.III.1949, EYD 6583 (LAM 54790); 18.XI.1986, XI-4 (ENCB 7959); Bahía Asunción, 28.IV.1950, EYD 9177 (UC 925585); 26.VII.1989, s/n (ENCB 7955); Isla Magdalena (Punta Entrada), 2.V.1950, EYD 9298 (UC 974662); Bahía Magdalena, 23.IV.1987, s/n (ENCB 7954).
Distribución latitudinal de las fases reproductivas: El análisis de la información de herbarios y recolectas realizadas, confirmó la presencia de L. aspergillum en 36 localidades a lo largo de la costa occidental de la península de Baja California (Fig. 1); además, reveló una marcada variación latitudinal de los talos tetrasporangiales, bisporangiales y gametangiales (carposporangiales y espermatangiales) (Fig. 1). Los talos tetrasporangiales muestran una amplia distribución geográfica, al estar presente en todo el intervalo de distribución conocido para L. aspergillum en la costa occidental de la Península de Baja California. Las plantas bisporangiales están restringidas hacia la parte noroccidental de la Península, desde las Islas Coronado hasta Punta Santa Rosalía y Bahía Tortugas, siendo estos los ambientes costeros que muestran la mayor influencia del fenómeno de surgencias. Por otro lado, las escasas plantas gametangiales se presentan, principalmente en la parte sur de la Península, en ambientes protegidos que muestran amplios intervalos de temperatura durante el año, como son Bahía Vizcaíno, Bahía Tortugas y Bahía Magdalena.
Discusión
Los talos de Lithothrix aspergilum son perennes y crecen sobre sustrato rocoso, epífitos o epizoicos (Emerson y Zedler, 1978; Aguilar-Rosas y Machado Galindo, 1990). La talla de los talos analizados para la costa del Pacífico de México es de 2-12 cm, ligeramente más grandes a los registrados previamente por Dawson (1953) para la misma región, y correspondiendo a los intervalos conocidos para California, E.U.A. (Ganesan y Desikachary, 1970; Abbott y Hollenberg, 1976). Dawson (1953), al describir los talos de L. aspergillum para el Pacífico mexicano, se basó sólo en talos tetrasporangiales, las plantas gametangiales no eran conocidas para el área. Además, indicó haber observado en un espécimen de Punta Banda que el ¨tetrasporangio¨ mostró solo una división produciendo bisporas. Por otro lado, Tyrrell y Johansen (1995) y Pearson y Murray (1997), establecen que a lo largo de la costa Pacifico de México, las poblaciones de L. aspergillum se mantienen solo a través de talos bisporangiales o vegetativos, estando las plantas tetrasporangiales y gametangiales ausentes; debido a la presencia de aguas frías producidas por el fenómeno de surgencias a lo largo de la costa, cuya temperatura superficial del agua de mar se encuentra por debajo de los 14 °C (Dawson, 1951, 1954); además, indican que en la costa sur de California se presentan temperaturas promedio del agua de mar por arriba de los 15-16 °C, mismas que favorecen la presencia de todas las fases reproductivas (asexuales y sexuales). Sin embargo, a partir de nuestros resultados todas las fases reproductivas (bisporangiales, tetrasporangiales, gametofíticas) de Lithothrix aspergillum se encuentran distribuidas a lo largo de la costa occidental de la Península de Baja California. En general, los talos tetrasporangiales muestran una amplia distribución geográfica; mientras que los talos bisporangiales están restringidos a la parte noroccidental de la Península y los escasos talos gametangiales (femeninos y masculinos) son comunes en la parte sur de la Península. Posiblemente, las áreas costeras influenciadas por aguas frías de surgencias, con temperaturas menores de 14 °C (Dawson, 1951; 1954).y la presencia de bahías protegidas con variaciones de temperatura anuales entre 16-30 °C (Gómez-Gutiérrez et al., 2001), son las que determinan el patrón latitudinal de las fases reproductivas en el Pacífico de México.
Dependiendo de la fase reproductiva, las plantas muestran una marcada variación morfológica. En general, los talos bisporangiales son más grandes (> 10 cm), menos ramificados y arborescentes, con un mayor número de conceptáculos en los ejes; en comparación a los talos tetrasporangiales que son pequeños (< 5.5 cm), compactos, más ramificados y un menor número de conceptáculos en los ejes. Los talos gametangiales (carposporangiales y espermatangiales) son más pequeños, con numerosas ramas laterales. Las plantas tetrasporangiales, bisporangiales y carposporangiales, no se pueden distinguir superficialmente, ya que los conceptáculos son generalmente globulares y extremadamente prominentes, en comparación con las plantas espermatangiales que se reconocen por sus conceptáculos pequeños y en ocasiones inconspicuos (Ganesan y Desikachary, 1970; Segawa, 1947; Tyrrell y Johansen, 1995).
Al comparar los caracteres morfométricos de los conceptáculos para los talos tetrasporangiales, bisporangiales, carposporangiales y espermatangiales descritos por Ganesan y Desikachary (1970) para California, E.U.A., con los determinados en el presente estudio, se encontraron algunas diferencias. Nuestros datos indican que las cámaras tetrasporangiales y bisporangiales son más grandes; por otro lado, las cámaras carposporangiales son más anchas y de menor altura. Las dimensiones de los tetrasporangios y bisporangios corresponden a los intervalos determinados por Ganesan y Desikachary (1970) para California y a los registrados para la costa del Pacífico de México por Dawson (1953).
Lithothrix aspergillum posee un ciclo de vida trifásico común en algas rojas, con talos bisporangiales, tetrasporangiales y gametangiales (masculinos y femeninos) que tienen características morfológicas y reproductivas bien definidas (Ganesan y Desikachary, 1970; Murray y Dixon, 1992). Estudios previos han demostrado que L. aspergillum puede regenerar sus ejes rápidamente a partir de remanentes basales perennes o crecer a partir de fragmentos desprendidos (Gittins, 1975; Tyrrell y Johansen, 1995), de tal manera que L. aspergillum mantiene sus poblaciones por medio de propagación vegetativa o la producción asexual de bisporas, donde talos gametangiales no están presentes (Pearson y Murray, 1997).
Pearson y Murray (1997) han sugerido que las poblaciones en las cuales dominan los talos bisporangiales o que están ausentes de conceptáculos, se mantienen por medio del reclutamiento de esporas recibidas de áreas contiguas, por medio de una propagación vegetativa a partir de las costras básales perennes o representan un remanente aislado, establecido por un evento fundador. Este mecanismo de mantenimiento, se ha sugerido para las poblaciones de L. aspergillum localizadas en el limite norte de distribución geográfica en la costa Pacifico de Norteamérica por DeWreede y Vandermeulen (1988), ya que después de realizar un estudio intensivo de 3 años en el estrecho de Georgia, no encontraron conceptáculos en los talos de L. aspergillum.
Tyrrell y Johansen (1995), han argumentado que si las bisporas germinan para desarrollar talos bisporangiales, entonces L. aspergillum presenta dos tipos de ciclos de vida, uno trifásico y otro bisporangial, similar al registrado para Titanoderma corallinae (Chamberlain, 1987). Nuestros resultados indican, que algunas de las poblaciones de L. aspergillum presentes en la costa Pacífico de México, se desarrollan a través de un ciclo de vida trifásico, de la misma manera que se presenta en las poblaciones ubicadas al sur de California (Tyrrell y Johansen, 1995; Pearson y Murray, 1997).
Agradecimientos
Se agradece a la Facultad de Ciencias Marinas, al Instituto de Investigaciones Oceanológicas de la Universidad Autónoma de Baja California y al Instituto Politécnico Nacional, el apoyo brindado para la realización del presente trabajo. La tercera autora agradece la beca otorgada por la Comisión, Operación y Fomento de Actividades Académicas del IPN. A los curadores de los Herbarios CMMEX, ENCB, LAM y UC, quienes otorgaron las facilidades requeridas para la consulta de sus colecciones. Los autores agradecen los comentarios y sugerencias de C. Wallace do Nascimento Moura y un revisor anónimo, para mejorar este trabajo.
Referencias
ABBOTT, I. A. & G. J. HOLLENBERG. 1976. Marine algae of California. Stanford University Press, Stanford, California. 827 p. [ Links ]
AGUILAR-ROSAS, R. & A. MACHADO-GALINDO. 1990. Ecological aspects of Sargassum muticum (Fucales, Phaeophyta) in Baja California, Mexico: reproductive phenology and epiphytes. Hydrobiologia 204/205:185-190. [ Links ]
AGUILAR-ROSAS, R., I. PACHECO-RUIZ & L. E. AGUILAR-ROSAS. 1990. Algas marinas de las Islas Todos Santos, Baja California, México. Ciencias Marinas 16:117-129. [ Links ]
CHAMBERLAIN, Y. 1987. Conceptacle production and life history in four species of Pneophyllum (Rhodophyta, Corallinaceae) from the British Isles. British Phycological Journal 22:43-48. [ Links ]
DAWSON, E.Y. 1945. Marine algae associated with upwelling along the northwestern coast of Baja California, Mexico. Bulletin Society California Academy Science 44:57-71. [ Links ]
DAWSON, E.Y. 1951. A further study of upwelling and associated vegetation along Pacific Baja California, Mexico. Journal of Marine Research 10:39-58. [ Links ]
DAWSON, E.Y. 1953. Marine red algae of Pacific México. I. Bangiales to Corallinaceae subf. Corallinoideae. Allan Hancock Pacific Expeditions 17:1-239. [ Links ]
DAWSON, E. Y. 1954. On correlation of marine vegetation with upwelling along the Pacific coast of Baja California, Mexico. In: K. Nisizawa (ed.) Proceeding of the Seventh International Botanical Congress 827 p. [ Links ].
DEWREEDE R. E. & H. VANDERMEULEN. 1988. Lithothrix aspergillum (Rhodophyta): regrowth and interaction with Sargassum muticum (Phaeophyta) and Neorhodomela larix (Rhodophyta). Phycologia 27:469-476. [ Links ]
EMERSON, S. E. & J B. ZEDLER. 1978. Recolonization of intertidal algae: An experimental study. Marine Biology 44:315-324. [ Links ]
GANESAN, E. K. & T. V. DESIKACHARY. 1970. Studies on the morphology and reproduction of the Articulated Corallines-V. Lithothrix Gray. Phycos 9:41-51. [ Links ]
GITTINS, B. T. 1975. The Biology of Lithothrix aspergillum J. E. Gray (Corallinaceae, Rhodophyta). PhD dissertation, University of California, Irvine, 188 p. [ Links ]
GOMEZ-GUTIERREZ, J, PALOMARES-GARCIA, R, HERNANDEZ-TUJILLO, S. 2001. Community structure of zooplankton in the main entrance of Bahía Magdalena, México during 1996. Revista de Biología Tropical 49:454-558. [ Links ]
GRAY, J. E. 1867. Lithothrix, a new genus of Corallinae. Journal of Botany 5:33. [ Links ]
HOLMGREN, P. K., N. H. HOLMGREN & L. C. BARNETT. 1990. Index Herbariarum. I. The Herbaria of the World. New York Botanical Garden, Bronx, N.Y. (Regnum Vegetabile vol. 120). 693 p. [ Links ]
MARTOJA, R. & M. MARTOJA-PIERSON. 1970. Técnicas de histología animal. Barcelona. Toray-Masson. 65 pp. [ Links ]
MATEO-CID, L. E. & A. MENDOZA-GONZALEZ. 1994. Estudio florístico de las algas bentónicas de Bahía Asunción, Baja California Sur, México. Ciencias Marinas 20:41-64. [ Links ]
MENDOZA-GONZALEZ, A. & L. E. MATEO-CID. 1985. Contribución al estudio florístico ficológico de la costa occidental de Baja California, México. Phytologia 59:17-33. [ Links ]
MENDOZA-GONZALEZ, A. & L. E. MATEO-CID. 1992. La familia Corallinaceae (Rhodophyta, Corallinales) en la Península de Baja California. Memorias del IX Simposio Internacional de Biología Marina, La Paz, B.C.S., México. 49-53 pp. [ Links ]
MURRAY, S. N & P. S. DIXON. 1992. The Rhodophyta: some aspects of their biology. III. Oceanogr. Marine Biology Annual Review 30:1-148. [ Links ]
PEARSON, E. A. & S. N. MURRAY. 1997. Patterns of reproduction, genetic diversity, and genetic differentiation in California population of the geniculate coralline alga Lithothrix aspergillum (Rhodophyta). Journal of Phycology 33:753-763. [ Links ]
SCAGEL, R. F., P. W. GABRIELSON, D. G. GARBARY, L. GOLDEN, M. W. HAWKES, S. C. LINDSTROM, J. C. OLIVEIRA & T. B. WIDDOWSON. 1989. Synopsis of the Benthic Marine Algae of British Columbia, Southeast Alaska, Washington and Oregon. Phycological Contribution No. 3, Univ. of British Columbia. 532 p. [ Links ]
SEGAWA, S. 1947. Systematic anatomy of the articulated corallines (XI). Lithothrix aspergillum Gray. Seibutu 2:87-90. [ Links ]
TYRELL, B. & H. W. JOHANSEN. 1995. Reproductive and regenerative strategies of Lithothrix aspergillum (Corallinales, Rhodophyta) in southern California. Phycologia 34:39-44. [ Links ]

