










Serviços Personalizados
Journal
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.15 no.3 Ciudad de México 2005
Article
Age, growth and mortality of Caulolatilus princeps (Pisces: Malacanthidae) from the southern Gulf of California
Edad, crecimiento y mortalidad de Caulolatilus princeps (Pisces: Malacanthidae) en el sur del Golfo de México
Juan F. Elorduy-Garay1, Sergio S. Ruiz-Córdova2 and Juan G. Díaz-Uribe3
1 Depto. de Pesquerías y Biología Marina, IPN-CICIMAR; Apdo. Post. 592, 23000, La Paz, B.C.S., México; Tel. (612) 122 5344; Fax (612) 122 5322. e-mail: jelorduy@ipn.mx
2 Dept. of Fisheries & Allied Aquacultures, Auburn University, Auburn, AL 36849-5419, U.S.A.
3 INP, Centro Regional de Investigaciones Pesqueras de La Paz, , La Paz, B.C.S., 23080, México.
Recibido: 14 de septiembre de 2004
Aceptado: 28 de abril de 2005
Abstract
The morphometric relationships, age, and growth of the Ocean Whitefish (Caulolatilus princeps, Jenyns 1842) were studied from a sample of 1783 specimens (1196 individuals for ageing), taken around the Bay of La Paz, in the southern Gulf of California from February 1986 to May 1987. The standard length ranged from 152 to 505 mm, and the eviscerated weight from 150 to 2660 g. The standard length-eviscerated weight relationships were estimated as: W = 0.00029 x L2.922, W = 0.00024 x L2.960, W = 0.00021 x L2.979 for males, females and the sexes combined, respectively. Twenty one age classes, ranging from 1 to 21 years were defined by whole burned otolith readings. Mean standard length, and standard length by otoliths edge-at-age data for the first 16 age groups were used to estimate the growth parameters of the von Bertalanffy equation: L∞ = 476.7 mm, K = 0.093, to = -3.904, for males; L∞ = 474.8 mm, K = 0.101, to = -3.339, for females; L∞ = 460.9 mm, K = 0.113, to = -2.765, for the sexes combined. The asymptotic weights were: 1847.6, 2006.8 and 1972.1 g, for males, females, and the sexes combined, respectively. Total (Z year-1) and natural (M year-1) mortalities were estimated as, respectively: 0.45 and 0.15 for males, 0.42 and 0.15 for females, and 0.41 and 0.17 year-1 for the sexes combined. The exploitation ratios (E) were: 0.67 for males, 0.63 for females, and 0.60 for the sexes combined, indicating that the fishing pressure on this population is higher than the recommended ratio of 0.5.
Key words: Malacanthids, morphometric relationships, otoliths, Gulf of California.
Resumen
Investigamos las relaciones morfométricas, edad y crecimiento de la pierna, Caulolatilus princeps (Jenyns, 1875), en una muestra de 1783 peces (1196 para lecturas de edad), del sur del Golfo de California (cercanías de la Bahía de La Paz) desde febrero 1986 hasta mayo 1987. La longitud estándar varió de 152 a 505 mm, y el peso eviscerado de 150 a 2660 g. Las relaciones peso-longitud se estimaron en: W = 0.00029 x L2.922, W = 0.00024 x L2.960, W = 0.00021 x L2.979 para machos, hembras y sexos combinados, respectivamente. Leyendo otolitos enteros quemados se definieron veintiuna clases de edad, desde 1 año. Con los datos de longitud estándar, promedios y por tipo de borde, de cada edad se estimaron los parámetros de la ecuación de crecimiento de von Bertalanffy: L∞ = 476.7 mm, K = 0.093, to = -3.904, para machos; L∞ = 474.8 mm, K = 0.101, to = -3.339, para hembras; L∞ = 460.9 mm, K = 0.113, to = -2.765, para sexos combinados. Los pesos asintóticos fueron: 1847.6, 2006.8 y 1972.1 g, para machos, hembras, y sexos combinados, respectivamente. Las mortalidades total (Z año-1) y natural (M año-1) se estimaron en, respectivamente: 0.45 y 0.15 para machos, 0.42 y 0.15 para hembras, y 0.41 y 0.17 año-1 para sexos combinados. Las tasas de explotación (E) fueron: 0.67 para machos, 0.63 para hembras, y 0.60 para sexos combinados, lo que indica que la presión de pesca sobre esta población is superior a la tasa de 0.5 recomendada.
Palabras clave: Malacántidos, relaciones morfométricas, otolitos, Golfo de California.
Introduction
The Ocean Whitefish, Caulolatilus princeps, Jenyns, 1842, is a malacanthid (classified as Branchiostegidae by Dooley, 1978) found from Vancouver Island (Canada) to the Gulf of California (México), and from Ecuador, and the Galapagos Islands to Peru (Dooley 1978). They are most commonly found at depths ranging from 80 to 150 m, and over sandy or muddy substrate. This species and the co-occurring Caulolatilus affinis form part of the catch in artisanal fisheries of Baja California Sur, and are reported with the common name of "pierna" in the local landings reports (SAGARPA, 2002). They are captured all the year round by hook and line, although they also appear in by-catches of shrimp trawlers (Elorduy-Garay, 1993). The 2000 pierna catch from Baja California Sur (1,073 t) was about 93% of the total Mexican catch (SAGARPA, 2002), with near 60% of that from the Bay of La Paz area (Ramírez-Rodríguez & Hernández-Herrera, 1999-2003). There is no certainty of different stocks along the distribution range of the species, but based on the grounds were it is fished we can assume to be at least two stocks in the Baja California region, one on the west coast of the Baja California Peninsula, as far north as San Ignacio, and the other in the Gulf of California.
The biology of C. princeps has not been very much studied. Moser et al. (1986) described the larvae from the California (USA) and Baja California (Mexico) region. Elorduy-Garay & Ramírez-Luna (1994) found this species to be a partial spawner with a pro-truded reproductive season (from October to March) in the Gulf of California. Elorduy-Garay & Caraveo-Patiño (1994) found this species to be an oportunistic predator of crustaceans and fishes in the same region. Caulolatilus affinis, a co-occurring species with Caulolatilus princes, has been recently described in terms of age, growth, and mortality (Elorduy-Garay & Díaz-Uribe 1994, Elorduy-Garay & Ruiz-Córdova 1998), which lives up to 16 years. Elorduy-Garay & Peláez-Mendoza (1996) described the feeding behaviour of C. affinis as similar to C. princeps. The reproduction of C. affinis has been found very similar to that of C. princeps (Ceballos-Vázquez & Elorduy-Garay 1998). For other malacanthid species (particularly from the Atlantic Coast of the U.S.A.) there have been a number of reports on their life histories and fisheries, including detailed descriptions of age and growth (Ross & Huntsman 1982, Ross & Merriner 1983, Turner et al. 1983, Harris & Grossman 1985, Erickson & Grossman 1986).
Since age and growth studies of commercially exploited species began, many authors have stressed the importance of proving that the method used for age determination should be valid for each species considered, even when two species are ecologically or taxonomically related (Beamish & McFarlane 1983). Elorduy-Garay & Díaz-Uribe (1994) showed that otoliths from C. affinis are adequate structures for age determination purposes. Therefore, the objective of this work is to validate the age reading of otoliths from C. princeps, and estimate the individual growth of the species, and to estimate mortalities and exploitation rates.
Materials and methods
Samples of C. princeps were collected monthly, from February 1986 to May 1987, mainly from the commercial catches present in some fish markets of La Paz and whose origin was verified to be from the Bay of La Paz, Cerralvo or San José islands (Fig. 1). A small number of samples were collected from research fishing using hook and line similar to those taken from the commercial fishery.
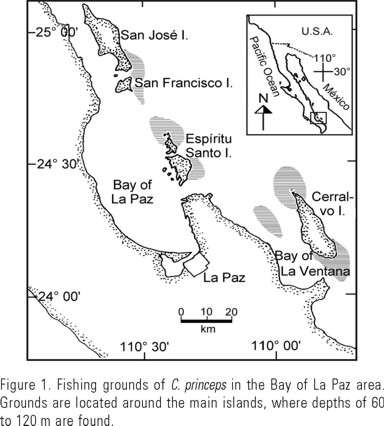
Fish were measured in total (TL) and standard (SL) lengths (to the nearest mm), weighed with an electronic balance (5 g precision), and sexed where possible. As most fish sampled were already eviscerated (without entrails and gills), a functional lineal regression was performed, by sex and the sexes combined, using those in which both weights (total and eviscerated) were available (Ricker 1973). This was done for comparative purposes, because measuments taken by different authors vary. Standard length was grouped into 10 mm size classes, which is the most common class for the species in this family. Both sagittae otoliths were collected and stored dry in plastic envelopes until their processing at the laboratory.
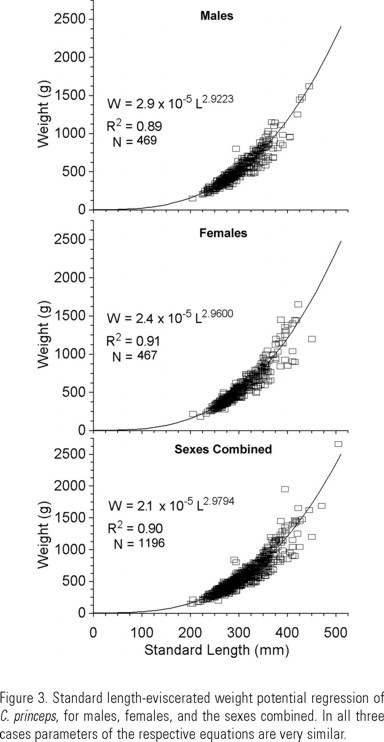
The standard length-eviscerated weight relationship was adjusted to a potential regression line (W = a Lb), where W is the weight (g), and L is the standard length (mm), and a and b are constants. A Student t test was performed to show if b deviated significantly from 3 (Pauly 1984).
A total sample of 1196 otoliths was thoroughly cleaned and burned in an oven at 325 °C for five minutes (Elorduy-Garay & Díaz-Uribe 1994). Otoliths were immersed in glycerol after burning, and read with reflecting light against a black background under a dissecting microscope calibrated to 10x. A random subsample of 185 otoliths was taken for a partial validation, that comprised counting of growth rings, edge type determination, and long radius measurements. For ageing of the total sample only the edge type and number of growth rings were considered. Otoliths were read by three readers independently, and without knowledge of length, weight, and sex of the fish. An analysis of variance with a Kruskal-Wallis test (Sokal & Rohlf 1979) was applied to the long radius measurements to test the homoscedasticity between readers.
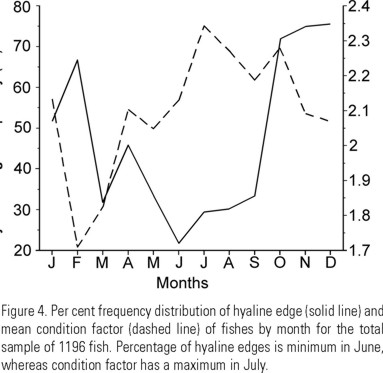
Validation was carried out under the criteria of proportionality and seasonality (Brothers 1979). Proportionality was analyzed by linear regression of the standard length of fish on otoliths long radius. Due to the high dispersion of points around the regression line, crude data were grouped into 0.2 mm long radii classes. Also, the three regression lines obtained from grouped data were compared by a multiple comparison test of the slopes and elevations (Zar 1974). Seasonality of ring deposition was analyzed by the monthly distribution of hyaline edges and compared to the monthly average of the condition factor (CF): CF = W * 100 / Lb, where b is the exponent of the length-weight relationship (Pauly 1984), under the consideration that when the condition factor is high the hyaline edges should be low and viceversa.
Mean length-at-age were calculated from the whole sample with and without otolith's edge consideration, for males, females, and the sexes combined. We used Ford-Walford's method (Walford 1946) for a first estimation of L∞, while K and to were calculated linearizing the VBGF; these first estimates of the von Bertalanffy parameters were used as "seeds" to estimate de final growth parameters of the VBGF (Ricker 1975) by a non linear least squares regression using Marquardt's algorithm (Conway et al. 1970). VBGF was fitted to data both with and without edge consideration. VBGF in terms of weight used the same K and to parameters, W∞ was calculated from the length-weight potential regression using L∞, and b is the exponent of the potential regression.
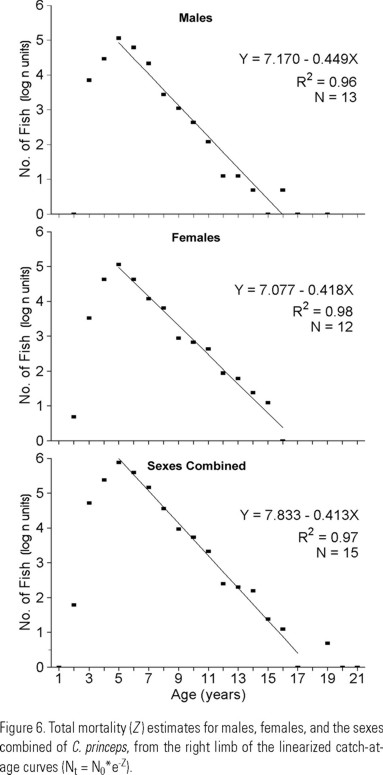
Mortality was estimated for the total sampling period. The total instantaneous mortality rates (Z ) were calculated from the catch-at-age curves (Nt = N0 * e-Z). Natural mortality (M) was estimated using L∞, K, and mean water temperature for the area (18°C for depths from 30 to 100 m; Cole & McLain 1989) based on the empirical equation of Pauly (1983). Fishing mortality (F ) was calculated as F = Z-M. The exploitation rate was calculated as E = F / Z.
Results
Morphometric data were obtained for a total of 1783 specimens (759 males, 751 females, and 270 not sexed). Otoliths were sampled from 1196 specimens (469 males, 467 females, and 260 not sexed). This sample size was used for length-weight calculations. The subsample for validation consisted of 185 individuals (75 males, 72 females, and 38 not sexed).
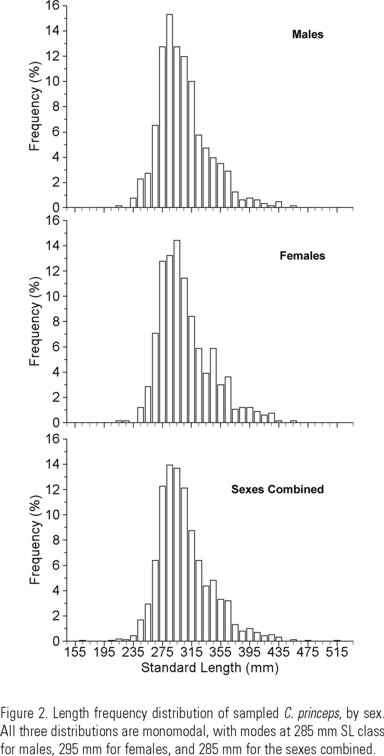
Standard length for the total sample ranged from 152 to 505 mm. There was a higher proportion of females in the larger length classes, although both sexes were represented to the same class interval (445 mm) (Fig. 2). The eviscerated weight ranged from 150 to 2660 g for the sexes combined, 150 to 1620 g for males, and 180 to 1650 g for females. The lineal regressions of standard length on total length, and eviscerated weight on total weight were highly significant in all cases (R 2 > 0.93) (Table 1). The standard length-eviscerated weight relationships showed R 2 ≥ 0.90 in all cases (Fig. 3). The slopes are lower, but not significantly different from 3. The same is true for the total length-total weight relationships. Therefore we conclude that C. princeps has an isometric growth pattern. The estimated exponents of the standard length-eviscerated weight relationships were used for the calculation of the condition factor and of W∞ by sex. As there were no significant differences between condition factors of females and males, only the sexes combined value was used.
Validation: A total of 23 otoliths (12.6%) from the sub-sample was discarded, mainly because they were broken, lost or deformed, or because of disagreement in the three readings. Most of the disagreements between readers of the otoliths of C. princeps occurred on those of larger size and those that apparently had double rings. Measurements from the readers were homogeneous (Hrm = 0.0376, P > 0.05), so they were averaged for each otolith.
Standard length of fish and otolith radius had a linear relationship in all cases, and the parameters from crude and grouped data sets were in every case not significantly different (Student t-tests, P > 0.05) (Table 2). The multiple comparison test showed that the three regression lines (for males, females, and the sexes combined) were very similar (F0 .05(1)(2, 33) = 3.29 > F( slope) = 1.5967, and F0 .05(1)(2, 35) = 3.27 > F( elevation) = 0.2120). Therefore, a "common" regression line (Zar, 1974) could be used, with a slope bc = 7.3387, and an intercept ac = 6.4934.
The hyaline edge of the otoliths dominated from September-October to February-March, which was coincident with the descent of the condition factor of the fish (July-March) (Fig. 4).
Age and Growth: Males were represented by age classes 2 to 19 (except 18), with a predominance of age class 5 (27.6%). Females were present in age classes 2 to 16, with the same age class 5 (27%) as the dominant one. The sexes combined (including those whose sex could not be determined) had age classes from 1 to 21 (except 18 and 20). Age class 5 dominated in the material (27%).
For the estimation of the VBGF parameters the following data, up to 16 years of age, were considered: the observed length-at-age without consideration of the otoliths edge, and the observed lengths considering the otoliths edge. VBGF parameters for males and females are similar, L∞ and to are slightly lower in females, and k is a little higher (Fig. 5).
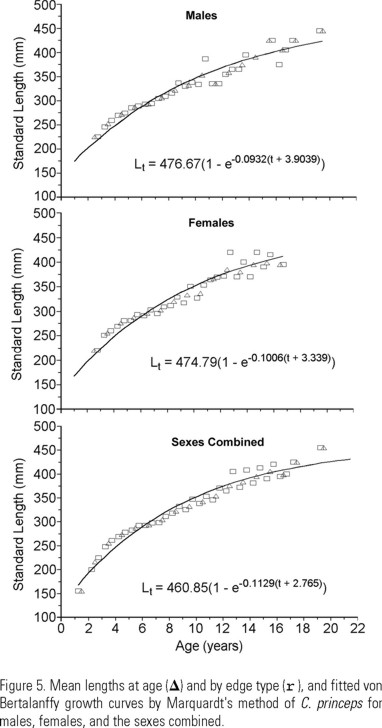
In terms of length the resulting equations were:

During the first 10 years there is alternation between mean lengths of males and females of the same age; from the 11th to the 14th year females have larger lengths than males; from the 15th year onwards males have larger mean lengths than females of the same age.
In terms of weight the resulting equations were:
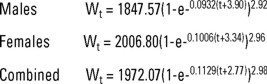
Mortality: Ages considered for the calculation of mortalities were 5-17 for males, 5-16 for females, and 5-20 for the sexes combined. Total mortality (Z ) was 0.45, 0.42, and 0.41 (Fig. 6); natural mortality (M ) was 0.15, 0.15, and 0.17; fishing mortality (F ) was 0.30, 0.26, and 0.25; and the exploitation rate (E ) was 0.67, 0.63, and 0.60, in all cases for males, females and the sexes combined, respectively.
Discussion
Collecting from the commercial fishery usually renders a low representation of young and small fish in the samples, especially if fishing is made with hook and line gear which are highly size selective. We consider our samples as representative because the length structure of fish taken by us at research fishing did not differ from those taken at the market place. Sex ratio was 1:1 for the total sample, and the subsamples. Length frequency distributions showed no differences in the modes of males and females. In contrast, various authors have reported important differences in sex ratios among malacanthids, always with females predominating at small sizes and young ages (Ross & Merriner 1983, Erickson & Grossman 1986, Elorduy-Garay & Ruiz-Córdova 1998); most of these authors have indicated that protogynous hermaphroditism may be one of the possible explanations for this phenomenon.
Our data indicate that C. princeps has an almost isometric growth pattern, understood as an slope near the value of 3, even for gutted fish. The slopes of the fitted curves are very similar for both sexes. The ordinates at the origin differ slightly. They are positive in all cases. One of the postulates underlying the VBGF is the assumption of isometric growth (i.e., that b=3), so in our case the obtained b values can be used reliably to estimate the condition factor and the weight-related growth parameters.
The percentage of unreadable otoliths (12.6%) was similar to other studies with malacanthids: 10% for Lopholatilus chamaeleonticeps (Turner et al. 1983), 8% for C. affinis (Elorduy-Garay & Ruiz-Córdova 1998); and much lower than for C. microps (29%) (Ross & Huntsman 1982).
Ages were well represented in a number of age classes. Ages 1, 2, and 3 are not recruited to the stock, and age 4 is only partially recruited. Data show that both males and females of C. princeps reach at least 14 years of age, and those beyond age 14 are scarce. Due to the poor representation of fish older than 16 years (1 individual 17 yrs old, 2 individuals 19 yrs old, and 1 individual 21 yrs old) it was necessary to exclude them from the calculations of growth curve. Including those values led to overestimates of L∞ and to, and underestimates of K.
Growth in C. princeps is adequately represented by the VBGF as shown by the variance explained by the model for each case, with good agreement between the observed data and the fitted model. Observed standard lengths at ages 5 yr, 6 yr and 7 yr are relatively close to each other, and appear as if there were almost no growth in those years. This ages correspond to birth dates from 1979 to 1981, in those years there was a marked descent of mean sea surface temperature in the southern Gulf (D. Lluch Belda, pers.comm.). Nevertheless, no attempt has been made to statistically relate this descent in growth to any oceanographic data. Fish older than 10 years had a differential sex contribution: 46.9% were females, and only 30.6% males (another 22.5% could not be sexed).
The parameter K of the VBGF shows that Caulolatilus princeps is a slow-growing fish of medium to high longevity (one individual aged 21 years). Among the malacanthids studied so far, L. chamaeleonticeps is the species that attains the greatest longevity (35 yr, 1150 mm FL) (Turner et al. 1983); and, Branchiostegus japonicus japonicus reaches a maximum of 8 yr and 322 mm TL (Hayashi 1976).
Growth differences between sexes have been observed for most malacanthids (Hayashi 1976, Ross & Huntsman 1982, Turner et al. 1983, Elorduy-Garay & Ruiz-Córdova 1998). Males always grow faster (higher K) and larger (higher L∞) than females. Caulolatilus princeps could be taken as the exception to the rule, as both sexes have quite similar growth parameters. In fact, K value for females is about 8% higher than for males, while L∞ is almost the same (2 mm difference) for the two sexes.
Growth rates obtained for Caulolatilus princeps are in accordance with those of other malacanthids (Hayashi 1976, Ross & Huntsman 1982, Turner et al. 1983, Elorduy-Garay & Ruiz-Córdova 1998) and also with other families inhabiting rocky-bottom deep waters as the species under study, whose K values range from 0.10 to 0.22 and maximum ages range from 9 to 16 yr (Matheson et al. 1986).
Estimated t0 values for C. princeps (-3.9039 for males and -3.3390 for females) are large due to the low representation of 0+, 1+, 2+ and 3+ aged fish in the samples.
Instantaneous rates of total mortality (Z = 0.41 for the sexes combined) are higher than those estimated in C. microps (0.22) from North Carolina and South Carolina waters (Ross & Huntsman 1982) and L. chamaeleonticeps (0.10 to 0.25) from waters off Georgia (Harris & Grossman 1985), and similar to L. chamaeleonticeps (0.46) from the Middle Atlantic-southern New England region (Turner et al. 1983).
Total mortality rates estimated for C. princeps (0.41 - 0.45) are slightly lower than those estimated for C. affinis (0.43 - 0.51), the co-occurring species into the study area (Elorduy-Garay & Ruiz-Córdova 1998). However exploitation rates for both species are very similar: from 0.61 to 0.67 for C. princeps, and from 0.56 to 0.69 for C. affinis. This similarity could be explained since both species support the artisanal fishery of "pierna".
When basic data for stock assessment is incomplete, the Gulland's rule (Gulland 1971) has been claimed as a useful method to draw fisheries management advice. Recently it has been recognized that fishing mortality at the maximum sustainable yield (FMSY) can vary from 0.33 times the natural mortality M in short-lived species to 4*M in long-lived top-predator species (Caddy & Csirke 1983; Caddy & Mahon 1996). This translates into exploitation rates ranging from 0.25 to 0.80. According to maximum age (21 years) found for C. princeps and considering this species as an active predator (Elorduy-Garay & Caraveo-Patiño 1994), interval of estimated exploitation rates is not an unexpected result. Although more precise data on population dynamics and recruitment process are needed for the stock assessment there are evidences that the fishery could be reaching a risky level. Therefore, a management plan is needed in which a monitoring program has to be included.
Acknowledgments
This study was partially funded by the Secretaría de Educación Pública, contract DIGICSA: C88-01-0282. The work of Axayácatl Rocha in sampling and ageing is gratefully acknowledged. Our special thanks to the Cuevas family for their assistance in fishing, and to Mr. Reyes Barrón owner of "El Paton" fish market for the permission to take samples at his facilities. Our special thanks to Elis Glazier for the english edition of the manuscript. JFEG received fellowships from CONACyT, and COFAA and EDI from the Instituto Politécnico Nacional.
References
BEAMISH, R.J. & G.A. MCFARLANE. 1983. The forgotten requirement for age validation in fisheries biology. Transactions of the American Fisheries Society 112: 735-743. [ Links ]
BROTHERS, E.B. 1979. Age and growth studies on tropical fishes. In: Saila, S.B. & P.H. Roedel (Eds.). Stock assessment for tropical small scale fisheries. The University of Rhode Island. Kingston, RI, pp. 119-136. [ Links ]
CADDY, J.F. & J. CSIRKE. 1983. Approximations to sustainable yield for exploited and unexploited stocks. Oceanographie Tropical 18: 3-15. [ Links ]
CADDY, J.F. & R. MAHON. 1996. Puntos de referencia para la ordenación pesquera. FAO, Documento Técnico de Pesca 347. [ Links ]
CEBALLOS-VÁZQUEZ, B.P. & J.F. ELORDUY-GARAY. 1998. Gonadal development and spawning of Caulolatilus affinis, Gill 1865 (Pisces: Branchiostegidae) in the Gulf of California. Bulletin of Marine Science 63: 469-479. [ Links ]
COLE, A.D. & D.R. MCLAIN. 1989. Interannual variability of temperature in the North Pacific Eastern Boundary region, 1971-1987. NOAA Technical Memorandum NMFS, USA, 17 pp. [ Links ]
CONWAY, G.R., N.R. GLASS & J.C. WILCOX. 1970. Fitting nonlinear models to biological data by Marquardt's algorithm. Ecology 51: 503-507. [ Links ]
DOOLEY, J.K. 1978. Systematics and biology of the tilefishes (Perciformes: Branchiostegidae and Malacanthidae), with descriptions of two new species. National Oceanic and Atmospheric Administration Technical Report, National Marine Fisheries Service Circular 411: 1-79. [ Links ]
ELORDUY-GARAY, J.F. & J. CARAVEO-PATIÑO. 1994. Hábitos alimentarios de la pierna, Caulolatilus princeps Jenyns 1842 (Pisces: Branchiostegidae) en la Bahía de La Paz, B.C.S., México. Ciencias Marinas 20: 199-218. [ Links ]
ELORDUY-GARAY, J.F. & J.G. DÍAZ-URIBE. 1994. Age validation of Caulolatilus affinis, Gill 1865, (Pisces: Branchiostegidae) from the Gulf of California using otoliths. Scientia Marina 58: 277-287. [ Links ]
ELORDUY-GARAY, J.F. & A.K. PELÁEZ-MENDOZA. 1996. Hábitos alimentarios de Caulolatilus affinis, Gill 1865 (Perciformes: Branchiostegidae) en la Bahía de La Paz, B.C.S., México. Revista de Biología Tropical 44: 241-249. [ Links ]
ELORDUY-GARAY, J.F. & S. RAMÍREZ-LUNA. 1994. Gonadal development and spawning of female Ocean Whitefish, Caulolatilus princeps (Pisces: Branchiostegidae) in the Bay of La Paz, B.C.S., Mexico. Journal of Fish Biology 44: 553-566. [ Links ]
ELORDUY-GARAY, J.F. & S.S. RUIZ-CÓRDOVA. 1998. Age, growth, and mortality of C. affinis, Gill, 1865 (Osteichthyes: Branchiostegidae), from the Southern Gulf of California. Pacific Science 52: 259-272. [ Links ]
ERICKSON, D.L. & G.D. GROSSMAN. 1986. Reproductive demography of tilefish from the South Atlantic Bight with a test for the presence of protogynous hermaphroditism. Transactions of the American Fisheries Society 115: 279-285. [ Links ]
GULLAND, J.A. (ED.). 1971. The fish resources of the ocean. Fishing News (Books). Farnham, Surrey, G. Britain. 255 p. [ Links ]
HARRIS, M.J. & G.D. GROSSMAN. 1985. Growth, mortality and age composition of a lightly exploited tilefish substock off Georgia. Transactions of the American Fisheries Society 114: 837-846. [ Links ]
HAYASHI, Y. 1976. Studies on the growth of the red tilefish in the East China Sea -II. Estimation of age and growth from otolith reading. Bulletin of the Japan Society of Scientific Fisheries 42: 1243-1249. [ Links ]
MATHESON, R.H. III, G.R. HUNTSMAN & C.S. MANOOCH III. 1986. Age, growth, mortality, food and reproduction of the scamp, Mycteroperca phenax, collected off North Carolina and South Carolina. Bulletin of Marine Science 38: 300-312. [ Links ]
MOSER, H.G., B.Y. SUMIDA, D.A. AMBROSE, E.M. SANDKNOP & E.G.STEVENS. 1986. Development and distribution of larval and pelagic juveniles of Ocean Whitefish Caulolatilus princeps in the CalCOFI survey region. California Cooperative Fisheries Investigations Report 27: 162-169. [ Links ]
PAULY, D. 1983. Some simple methods for the assessment of tropical fish stocks. FAO Fisheries Technical Paper 234: 1-52. [ Links ]
PAULY, D. 1984. Fish population dynamics in tropical waters: a manual for use with programmable calculators. ICLARM Studies and Reviews 8: 325 pp. [ Links ]
RAMÍREZ-RODRÍGUEZ, M. & A. HERNÁNDEZ-HERRERA. 1999-2003. Sistema automatizado para el manejo de avisos de arribo (SIMAVI 1.0 ). IPN-CICIMAR, Conservation International, A.C., La Paz, B.C.S., 27 pp. [ Links ]
RICKER, W.E. 1973. Linear regressions in fishery research. Journal of the Fisheries Research Board of Canada 30: 409-434. [ Links ]
RICKER, W.E., 1975. Computation and interpretation of biological statistics of fish populations. Bulletin of the Fisheries Research Board of Canada 191: 1-382. [ Links ]
ROSS, J.L. & G.R. HUNTSMAN. 1982. Age, growth, and mortality of Blueline Tilefish from North Carolina and South Carolina. Transactions of the American Fisheries Society 111: 585-592. [ Links ]
ROSS, J.L. & J.V. MERRINER. 1983. Reproductive biology of the Blueline Tilefish Caulolatilus microps off North Carolina and South Carolina. NMFS Fisheries Bulletin 81: 553-568. [ Links ]
SAGARPA 2002. Anuario estadístico de pesca 2000. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación, México, D.F., 268 p. [ Links ]
Sokal, R.R. & F.J. Rohlf. 1979. Biometría: principios y métodos estadísticos en la investigación biológica. H. Blume Ediciones, Madrid. [ Links ]
TURNER, S.C., C.B. GRIMES & K.W. ABLE. 1983. Growth, mortality and age-size structure of the fisheries for Tilefish, Lopholatilus chamaelonticeps, in the Middle Atlantic-Southern New England Region. NMFS Fisheries Bulletin 81, 751-763. [ Links ]
WALFORD, L.A. 1946. A new graphic method of describing the growth of animals. Biological Bulletin 90: 141-147. [ Links ]
Zar, J.H. 1974. Biostatistical analysis. Prentice-Hall, Englewood Cliffs. [ Links ]

