










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.15 no.3 Ciudad de México 2005
Artículo
Distribución, cobertura, morfometría y concentración de pigmentos de Ulva fasciata Delile en la costa N de La Habana, Cuba
Distribution, cover, morfometry and pigments concentration of Ulva faciata Delile from the northern coast of Havana, Cuba
Mercedes Cano Mallo1, Jhoana Díaz Larrea1, Olga Valdés-Iglesias2, Miguel Gómez Bastista3 y Thierry Chopin4
1 Dpto. de Biología Marina, Instituto de Oceanología, CITMA 1ra Ave. # 18406, CP 12100, C. de La Habana, Cuba. e-mail: vfajer@ff.oc.uh.cu; ecomar@oceano.inf.cu
2 Dpto. de Bioactivos Marinos y Productos Naturales, Instituto de Oceanología, Loma y 37, Nuevo Vedado, C. de La Habana, CP. 10600. e-mail: cebimar@infomed.sld.cu
3 Laboratorio de Vigilancia Radiológica Ambiental del Centro, Carr. Castillo de Jagua, Km 1/2 CP 59350, Cienfuegos, Cuba. email: lvral@perla.inf.cu
4 Center for Coastal Studies and Aquaculture, Dep. of Biology, University of New Brunswick, PO Box 5050, St. John, N.B., E21 4I. 5. Canada. e-mail: tchopin@unbsj.ca
Recibido: 17 de julio de 2004
Aceptado: 5 de febrero de 2005
Resumen
En el presente trabajo se evaluó la distribución, el porcentaje de cobertura, la variación morfométrica y la concentración de clorofila a, b y carotenoides de Ulva fasciata Delile en el medio natural y la variación temporal y espacial de estas variables. Se realizó al norte N de La Habana, en la zona comprendida entre los ríos Quibú y Jaimanitas entre los meses de marzo de 1996 y julio de 1999. Se escogieron para realizar los muestreos tres zonas con características distintas en cuanto a la protección al oleaje y al aporte de nutrientes procedentes de ambos ríos. Se observó una variación morfométrica relacionada con la longitud y el ancho. Se presenta el análisis morfológico realizado, teniendo en cuenta las variaciones de estos caracteres. La especie no se distribuye de igual forma en toda el área: el mayor porcentaje de cobertura y las mayores tallas se observaron en los meses de enero a junio y el menor porcentaje de agosto a octubre. El sustrato rocoso favorece la cobertura algal. La morfometría de los ejemplares de Ulva fasciata difieren en los distintos ambientes naturales y las frondas mas largas y estrechas y de color verde intenso crecen cerca del río Quibú. En la zona aledaña al río Jaimanitas, tienen las frondas largas y anchas y son de color verde y las que crecen en la zona central presentan características intermedias. Se comprobó la riqueza de pigmentos fotosintéticos de Ulva fasciata en la zona. El valor medio la clorofila b varío entre 93.20 mg/g (b.s) - 1.10 mg/g (b.s), seguidos de la clorofila a con valores que oscilaron entre 7.83 mg/g (b.s) y 0.03 mg/g (b.s) y los carotenoides (1.2 mg/g (b.s)-0.03 mg/g (b.s)). Las diferencias de concentraciones de pigmentos en los distintos ambientes y meses, justifica las diferencias de tonalidades del color verde, que se pudieron observar en el vegetal con anterioridad.
Palabras clave: Distribución, cobertura, pigmentos, morfometría, Ulva fascista, Ulvaceae.
Abstract
In this paper was evaluated the distribution, cover percentage, morphometric variation and chlorophyll and carotenoides concentration of Ulva fasciata Delile in natural condition and was evaluated the seasonal and space variation. The work was carried in North N of Havana between Quibú river and Jaimanitas river between March/96 and July/99. The samples were chosen in three zones with different characteristics as the protection to the surf and the contribution of nutrients coming from both rivers. A morphometric variation related to the longitude and the width was observed. The morphological analysis done is presented, keeping in mind the variations of these characters. The results show that the distribution of the specie is not uniform. The station near of the Quibú river was the most productive. The good time of the development of U. fasciata happened between January and June and the time of less development was between August and October. There was significative differences in the cover percentage presented in the different substrates. The rock substrate presented the highest cover percentage values. The morphology and the size of the samples of Ulva fasciata differ in the different natural conditions. The samples that grew near of Quibú river had the longest and narrow fronds and had different morphotype, in the months of maximum growth. In the area near to the river Jaimanitas presents as characteristic distinctive the long and wide fronds with the flat borders. The algae that grew in the area more far from the outlet of the rivers, they present characteristic intermediate morphotipic. The chlorophyll a and b and carotenoides changed with the sample month and the study stations.
Key words: Distribution, cover, pigments, morphometric, Ulva fasciata, Ulvaceae.
Introducción
Algunos géneros de las algas verdes como Enteromorpha Link, Ulva Linnaeus, Bryopsis Lamouroux, Codium Stackhouse, Caulerpa Lamouroux y Monostroma Thuret son de gran importancia económica por su uso como alimento humano, en la extracción de bioactivos y en la preparación de cosméticos (Diaz-Piferrer & López,1959; De Lara-Isassi & Alvarez-Hernández,1994). En la actualidad, la explotación de estos géneros es muy limitada, Japón es uno de los países con tradición en el aprovechamiento de Ulva, Enteromorpha y Monostroma principalmente para el consumo humano y Francia, más recientemente, ha autorizado también el consumo de estas algas por su valor nutricional (Mabeau & Fleurence, 1993).
En Cuba no existe tradición de consumo de macroalgas marinas, pero se realizan intentos para el aprovechamiento de estos recursos, sobretodo aquellos que por su abundancia y valor nutricional permitan ser explotados de forma racional, de ahí la importancia de estudiar aspectos biológicos de las algas marinas en nuestras condiciones. Relacionado con el género Ulva, Diaz-Piferrer & López (1959) destacaron la relativa abundancia del género en las costas cubanas y su valor nutricional en la alimentación de las aves, basado en la estimación de la biomasa de las arribazones y la composición química en la zona aledaña al río Quibú. Estas algas también han sido probadas por Garmendía (1994), como aditivo alimentario en productos de amplio consumo como el pan, por las propiedades en cuanto a estabilidad y durabilidad que le confiere. En la década del 90', se priorizaron otras investigaciones sobre la Biología y el aprovechamiento de algunas macroalgas al norte N de La Habana, en particular la especie Ulva fasciata Delile, teniendo en cuenta los estudios que anteceden, su amplia distribución y relativa abundancia (desde la zona intermareal hasta 2 m de profundidad), si se compara con Ulva lactuca Linnaeus, la otra especie reportada para Cuba (Suárez, 1975) y presente en esta zona, pero que solamente está presente en áreas muy localizadas de la zona intermareal. En este trabajo se abordan algunos aspectos biológicos como parte de un estudio integral realizado entre 1996 y 2000, para el manejo de este recurso en el área, para su uso como suplemento en la dieta animal y/o humana y la industria cosmetológica.
Materiales y métodos
Área de estudio
El estudio se realizó entre el río Quibú (82°27'54" N y 23°53'04" W) y el río Jaimanitas (82°29'25" N y 23°05'34" W) en la costa norte N de La Habana, Cuba donde se asienta un banco natural extenso de Ulva fasciata Delile. Se escogieron para realizar los muestreos de todas las variables, tres zonas con características distintas en cuanto a la protección al oleaje y el aporte de nutrientes: dos de ellas, aledañas a las desembocaduras de los ríos y una zona central entre ambos ríos, donde el aporte de nutrientes procedentes de los mismos es menor y la exposición al oleaje es mayor por las características del área (Fig 1). La determinación y descripción de las poblaciones de Ulva fasciata se hizo consultando los trabajos de Taylor (1960), Litller & Litller (2000) así como los ajustes nomenclaturales de acuerdo con Wynne (1998) y Hayden et al. (2003).

Distribución y porcentaje de cobertura
Se ubicaron siete transectos perpendiculares a la línea de la costa; tres de ellos en la zona aledaña al Río Quibú (T1,T2 y T3), dos en la zona central (T4 y T5) y el resto en la zona aledaña al río Jaimanitas (T6 y T7). En cada transecto se evaluó el porcentaje de cobertura (%) y el tipo de sustrato, colocando una cuerda graduada como guía desde la zona intermareal hasta donde se observaron ejemplares de la especie; excepto en T4 donde se colocó la cuerda 150 m alejada de la costa, por existir una zona de playa (arena) muy extensa donde no hay vegetación. Se tiró al azar un cuadrado cuadriculado (25 cm x 25 cm) por triplicado cada 10 m y en cada cuadrado se estimó el área de cobertura que ocupó la especie y los diferentes sustratos (rocoso, roco-arenoso y arenoso); a partir de estas estimaciones se calculó el porcentaje de cobertura (%) y el tipo de sustrato, en este último caso se empleó la escala siguiente: rocoso > 75% con roca u otro sustrato duro, roco-arenoso -50% roca y 50% arena, y el Arenoso > 75% arena u otro sustrato blando. Se realizaron 10 muestreos en diferentes meses del año entre marzo de 1996 y julio de 1999.
Morfometría
Para el estudio morfométrico (largo/ancho) se colectaron ejemplares en iguales zonas a las descritas en el área de estudio, coincidiendo los lugares muestreados con los seleccionados para estimar el resto de las variables, incluyendo la concentración de pigmentos. En la zona aledaña al río Quibú se ubicaron dos transectos, en este caso, paralelos a la línea de la costa, uno de ellos en la zona intermareal, realizando la colecta desde el límite superior de marea hasta una profundidad menor de 0.50 m. El otro transecto se ubicó a una distancia de la costa entre 21m y 40 m, a 1 m de profundidad. En la zona central y en la zona aledaña al río Jaimanitas se ubicó un transecto paralelo a la línea de la costa; en el primer caso, separado de ella unos 100 m, a una profundidad que fluctuó entre 0.75 y 1 m; en el segundo caso se ubicó a 20 m de la costa, cerca de un caserío, cuyo lugar está protegido de la acción del oleaje por una cresta arrecifal y dos cayos pequeños. En cada uno de los transectos, se colectaron 25 ejemplares al azar a lo largo de 100 m, los cuales fueron trasladados al laboratorio. A cada ejemplar se le realizaron tres mediciones: largo de la mayor de las ramificaciones (l.max.); ancho máximo de dicha ramificación (a.max.) y largo promedio de tres ramificaciones (l.pm). Esta última variable no es más, que el largo promedio de las tres ramificaciones más largas sin tener en cuenta la mayor de las ramificaciones. Este estudio se efectuó en el período comprendido entre noviembre de 1996 y marzo de 1999 y se inició después de obtener los primeros resultados sobre la distribución y el porcentaje de cobertura de este recurso en el área, lo que permitió seleccionar las zonas de mayor interés. Los ejemplares fueron colectados en cuatro veces cada año con el objetivo de conocer la variación estacional que experimentan estos indicadores en cada zona.
Pigmentos
Para la determinación de la concentración de clorofila a, b y los carotenoides fueron colectados 25 ejemplares enteros a 1m de profundidad, en cada estación, en iguales zonas a las descritas en el área de estudio. Los lugares específicos dentro de cada zona se señalan en la Fig. 1. Las colectas se realizaron trimestralmente entre los meses de mayo de 1998 y julio de 1999. Las muestras fueron trasladadas al laboratorio en bolsas de polietileno negro y se conservaron a -20°C. Para la determinación de las clorofilas a y b los talos completos se cortaron en pequeñas porciones, se pesó un gramo de cada muestra por triplicado y se prepararon los extractos en acetona en un homogenizador Waring Blendor para la extracción total de los pigmentos durante 2 min. Los extractos se centrifugaron a 4000 rpm durante 20 min. Las determinaciones de la clorofila a y b se realizaron con el empleo de la ecuación descrita por Hellebust & Craigie (1978) y por Jeffrey & Humphrey (1975) respectivamente. Para la determinación de carotenoides, se tomó una parte del extracto en acetona y se neutralizó con carbonato de calcio hasta lograr la saturación del extracto, posteriormente se agitó manualmente y se mantuvo en reposo durante 24 h a -20°C.
Para la descripción cualitativa de la composición en pigmentos, se realizó una cromatografía en capa fina sobre sílica gel G como fase estacionaria, y como fase móvil éter de petróleo: acetona en una proporción 75:25. Se realizó un espectro de barrido en espectrofotómetro Shimadzu con el programa acoplado, a los extractos acetónicos con y sin carbonato de calcio.
El análisis de los datos se realizó utilizando el paquete estadístico Statistic Versión 4.2 y las pruebas estadísticas (Spiegel, 1977) realizadas fueron las siguientes: la prueba de Kruskall Wallis se aplicó a los datos del porcentaje de cobertura y las variables morfométricas (largo y ancho máximo, largo promedio) para comparar los datos entre las diferentes zonas y meses del año. Se aplicó la prueba de Spearman para conocer si existe correlación entre las variables morfométricas. Para el análisis de la concentración de pigmentos se utilizó la prueba de Mann-Whitney. En todos los casos si existía diferencia significativa se aplicó la prueba Student Newman Keuls (SNK) con un 95% de confiabilidad.
Resultados
Distribución y porcentaje de cobertura. La Figura 2 muestra el valor medio del porcentaje de la cobertura de Ulva fasciata en la zona aledaña al río Quibú. En esta zona se extiende un gran banco de Ulvaceas durante todo el año, que se deprime al final del verano (agosto-octubre). Su extensión varía con la época y aparece desde la zona intermareal hasta 100 m alejado de la costa (enero y marzo de 1996); los ejemplares fueron observados hasta 2 m de profundidad en marea baja (Tabla 2). El valor medio mas alto se registró en la zona somera, casi siempre en los primeros 80 metros desde la línea de la costa. En general, existe diferencia significativa al comparar esta variable en los distintos meses y entre las zonas (p<0.05). El valor medio hallado indica, que en enero de 1997 Ulva fasciata presentó un porcentaje de cobertura significativamente mayor, seguido de junio de 1997, junio de 1998 y marzo de 1996. Casi siempre, los registros mas bajos se hicieron en agosto y octubre (Tabla 1).

En esta zona los máximos valores del porcentaje de cobertura en los meses más favorables, superan el 50% y difieren significativamente entre sí (p<0.05) al comparar los diferentes sustratos (rocoso, roco-arenoso y arenoso), en la mayoría de los datos contrastados. El valor medio más elevado se reporta donde predomina el sustrato rocoso (máximos entre 80% y 95%), seguido del sustrato roco-arenoso (60%-87%). En el sustrato arenoso, apenas se observan ejemplares de la especie y los valores máximos no exceden del 4% (Tabla 2).
En la zona central (Fig. 3) el banco de Ulva fasciata aparece en determinados lugares (T4) a partir de los 100 m de la costa y se observaron ejemplares hasta 1.5 m de profundidad. Este banco alcanzó una extensión máxima de 120 m en enero de 1997 (Tabla 2). Se encontraron diferencias significativas entre los meses de máximo y mínimo porcentaje de cobertura (p<0.05). En este lugar el mes más favorable es junio, seguido de enero y marzo y los registros mas bajos para la zona central en general coincidieron con los reportes de la zona aledaña al río Quibú (agosto y octubre) (Tabla 1).

En el Transecto 5, el banco de Ulva fascista aunque tiene una extensión máxima de 120 m, el porcentaje de cobertura es muy bajo y se ubica desde los 90 m de distancia de la costa hasta los 210 m, excepto en el mes de enero de 1997, cuando las algas aparecen más cerca de la costa (a partir de 30 m de la orilla entre 0.5 m y 1.5 m de profundidad). Se encontraron diferencias significativas al contrastar esta variable en la zona central en los distintos meses y diferentes sustratos (p<0.05) (Tabla 2).
En la zona aledaña al río Jaimanitas (Fig.4), la extensión del banco de Ulva varió con el lugar y los diferentes meses. En el transecto 6 la especie se observó durante todo el tiempo (excepto en junio de 1997), a una profundidad de 0.5 m y tuvo una extensión de 10 m. Al contrastar la media en los diferentes meses del año (p<0.05) se observaron diferencias significativas en el porcentaje de cobertura y los meses con mayores porcentajes fueron: junio/96, seguido de enero(97/99) y marzo/98 (Tabla 2).

En el transecto más cercano al río Jaimanitas (T7), Ulva fasciata apareció desde la zona intermareal hasta 170 m alejado de la costa. El mayor porcentaje de cobertura se evidenció en los primeros 50 metros separados de la costa y a partir de los 90 m; en correspondencia con los lugares más bajos y con el sustrato rocoso. En general, se presentaron diferencias significativas al comparar esta variable en los distintos meses del año (p<0.05); junio de 1996 fue el mes de mayor cobertura, seguido del mes de enero del 1997 y enero del 1998; el resto de los meses alcanzaron valores mucho más bajos y los mismos no difirieron entre sí. Al comparar el valor de la cobertura en los diferentes sustratos, variaron significativamente (p<0.05). Esta variable mostró un valor moderado en el sustrato roco-arenoso, aunque no excedió los valores máximos hallados en la zona aledaña al río Quibú, pero si fueron superiores a los obtenidos en la zona central (Tabla 2).
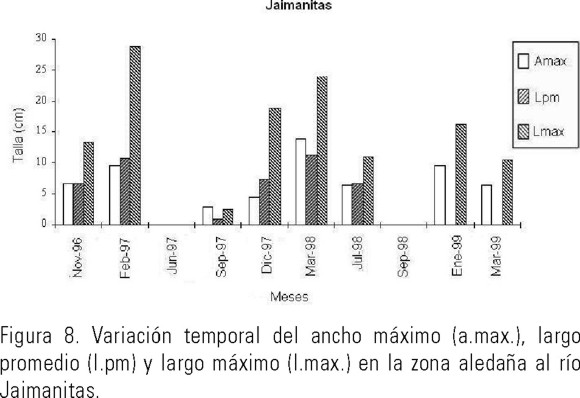
Morfometría. Los valores medios de la longitud y el ancho máximo de la ramificación más larga y el largo promedio del resto de las ramificaciones se muestran en las Figuras: 5-8 (6, 7). En las tres variables se observaron diferencias entre las medias en los diferentes meses y zonas.



Los ejemplares que crecen cercanos al río Quibú en la zona mas baja (Fig. 5), mostraron los mayores valores promedios de la longitud máxima en los meses de febrero y junio de 1997 y marzo de 1998. En el mes de septiembre desaparecieron las algas y en diciembre de ese mismo año se presentó una recuperación del banco con ejemplares muy pequeños cuyas tallas difirieron del resto de los meses. Se observó que el ancho medio de la ramificación más larga, es mucho menor que su longitud y menor que en el resto de las zonas. Los ejemplares colectados entre 21 y 40 m alejados de la costa (Fig. 6) en esta misma zona, alcanzaron también los valores mas altos de longitud máxima en los mismos meses y dichas frondas alcanzaron las máximas longitudes de toda el área, en algunas ocasiones duplican o triplican tanto el largo de la ramificación más larga como el resto de sus ramificaciones; sin embargo se apreciaron los valores más bajos del ancho al comparar estos resultados con las otras zonas. Se presentaron diferencias significativas en el largo entre los meses muestreados. En septiembre de 1997 no se observaron ejemplares a simple vista, pero en diciembre se pudo apreciar una recuperación del banco al igual que en la zona intermareal. Este fenómeno se presenta de forma similar al siguiente año en la misma época (Tabla 1).
Los valores medios del largo que experimentaron las frondas en la zona Central, se refleja en la Figura 7. Los mayores valores promedios alcanzados por la longitud máxima se expresaron en el período primavera-verano (junio/97, febrero/97 y marzo/98). Los meses de septiembre y diciembre reportaron tallas más pequeñas. En noviembre/96 desaparecieron los ejemplares en la zona, sin embargo la población se recuperó al finalizar el invierno. Se pudo comprobar que, tanto la longitud promedio de la ramificación más larga, como el resto de las ramificaciones no llegan a alcanzar, en la mayoría de los casos, valores tan altos como los que alcanzaron las frondas que se desarrollaron en la zona cercana a ambos ríos en un mismo mes y en profundidades similares.
Los mayores valores obtenidos del ancho máximo se reportaron en los meses de junio/97, febrero/97 y marzo/98. Los resultados demostraron diferencias entre los meses para todas las variables; se pudo comprobar también que en el caso de la longitud máxima, que febrero/97 y marzo/98, alcanzaron los valores más altos, seguidos de junio/97, septiembre/97 y diciembre/97 (Tabla 1).
En la Figura 8, se presenta el valor medio de las variables morfométricas calculadas en ejemplares que crecieron en la zona aledaña al río Jaimanitas. Los meses más favorables que estas algas alcanzaron las mayores tallas fueron: febrero/97 y marzo/98; junio/97 y septiembre/98 los meses más desfavorables, ya que no se encontró ningún ejemplar fijo al sustrato y las algas presentes estaban desprendidas. El largo promedio del resto de las ramificaciones, alcanzaron valores similares durante todo el tiempo de estudio, excepto el mes de septiembre/97 donde las algas estaban muy pequeñas, por lo que se infiere que el banco comenzó su recuperación. El ancho máximo de la ramificación más larga de los ejemplares que crecen en este lugar, difirieron entre sí en casi todos los meses y presentó un valor mayor que las frondas del río Quibú. El largo promedio alcanzado por del resto de las ramificaciones indicaron que las tallas fueron similares durante todo el año, excepto en el mes de Septiembre. El ancho de las frondas en este lugar muestra diferencias entre todos los meses y su máximo valor lo expresa en el mes de marzo/98 (Tabla 1).
El análisis de los resultados de las correlaciones realizadas entre las variables morfométricas mostró que: en la zona aledaña al río Quibú (0-20 m y 21-40 m) la longitud y el ancho de las frondas mas largas tuvieron una correlación inversa no significativa (rs = -0.18 y rs = -0.09). En este lugar (21-40 m), la longitud máxima y el largo promedio de la mayoría de las ramificaciones tuvieron una correlación directa significativa (rs = 0.58); la correlación entre estas dos variables fue directa no significativa en el resto del área. En los ejemplares que se desarrollaron cerca del río Jaimanitas, se evidenció una correlación directa significativa entre el largo y el ancho de las frondas mas largas (rs = 0.48*) y entre el largo máximo y el largo promedio del resto de las ramificaciones fue directa no significativa (rs = 0.35), ocurriendo de forma similar en la zona central entre estas variables (rs = 0.35) y entre el largo y el ancho de las frondas mas largas (rs = 0.33).
Los ejemplares de la zona del Quibú, mostraron una coloración verde intensa, la textura coreácea y su forma es alargada con el borde bastante rizado. Las frondas que se desarrollaron en Jaimanitas presentaron una coloración verde menos intenso, con bordes más lisos y la textura menos coriácea. En la zona central se observaron especímenes alargados y frondas mas anchas que los que crecieron cerca del río Quibú y su color es verde medio con menos rizos y su textura más suave. Estos especímenes en el invierno presentaron tallas pequeñas y en forma de roseta (Tabla 2).
Pigmentos. El espectro de absorción obtenido del extracto acetónico de U. fasciata ofreció 3 picos máximos: 432.7; 455.6 y 662.5 nm característicos de clorofila a, carotenoides y clorofila b.
El espectro de la muestra precipitada con carbonato de calcio solamente mostró tres máximos a 430, 546 y 665 nm, lo que indicó la presencia de carotenoides en U.fasciata. En la Figura 9 se observó el valor medio de la concentración de clorofila a, b y los carotenoides en la zona aledaña al río Quibú. Se observó diferencia significativa (p<0.05) al contrastar la media de los meses en los niveles de clorofila a y b. El máximo valor de estas variables se obtuvo en las muestras colectadas en los meses de septiembre de 1998, enero y marzo de 1999 (Tabla 1).

Los resultados de la concentración de pigmentos de los ejemplares colectados en la Playa Francesita aparecen en la Figura 10. El nivel de clorofila a en este lugar fue significativamente superior en los meses de mayo y julio (1998) y enero y julio (1999), si se comparan con el resto de los meses contrastados. La clorofila b, alcanzó valores mucho mas altos y no hubo diferencia significativa entre los meses. Se observó además que el valor máximo se obtuvo en septiembre (1998), seguido de julio (1998/1999) y enero y marzo (1999) (Tabla 1) .

Los valores medios de la concentración de clorofila a, b y carotenoides en el Club Habana se reflejan en la Figura 11. Las muestras evaluadas en este lugar, presentaron diferencias significativas (p<0.05) en el nivel de clorofila a y se destacaron con los valores mas altos los meses de julio (1998/1999) y mayo (1999). En el caso de la concentración de clorofila b no hubo diferencias significativas (p<0.05) al comparar los meses y los valores mas altos se reportaron en mayo (1999) seguido de julio y septiembre (1998) (Tabla 1).

La concentración de carotenoides en esta zona alcanzó su máximo en julio (1998) y difiere significativamente del resto de las medias contrastadas.
Los resultados obtenidos en las concentraciones de los distintos pigmentos en zona aledaña al río Jaimanitas aparecen en la Figura 12. No se observó diferencia significativa en el nivel de clorofila a y los carotenoides, pero sí en la concentración de clorofila b al contrastar los diferentes meses. En enero (1999), se obtuvo la mas alta concentración de este pigmento (Tabla 1).

En la Tabla 1 aparece un resumen que muestra el valor medio de las variables calculadas y las características de la especie en las tres zonas de estudio, para una mejor comprensión y manejo de la información.
Discusion
Distribución y porcentaje de cobertura. Los resultados de este estudio referentes a la distribución y abundancia relativa de Ulva fasciata, corroboran lo planteado por diferentes autores en cuanto a la permanencia del banco natural toda el área y la variación temporal y espacial de la comunidad algal (Díaz-Piferrer, 1959; Cano, 1996; Cano & Hernández, 1997 y Gómez, 1998). Se confirmó además que estos resultados fundamentan y permiten actualizar la descripción realizada por Díaz Piferrer, (1959) sobre las características de la zona aledaña al río Quibú, específicamente en lo concerniente a que el banco de Ulva está en el período de mayor productividad de enero a julio y la época de máximo rendimiento es de febrero a junio. Este autor comprobó además que a partir del mes de agosto a octubre hay un descenso paulatino de la productividad en términos de volumen de algas por colecta.
Los resultados obtenidos en este estudio sobre la distribución, el porcentaje de cobertura y el estudio morfométrico en la zonas central y aledaña al río Jaimanitas, permiten fundamentar que la especie tiene en general en toda el área como período favorable, la época de enero a junio y menos favorable de agosto a octubre.
La zona central a diferencia de las zonas aledañas a ambos ríos se reportaron los registros mas bajos del porcentaje de cobertura en la época mas favorable, pudiendo esto atribuirse a que esta zona está mas expuesta al oleaje y pueden desprenderse mayor número de los ejemplares, además de estar menos influenciada por el aporte de los ríos, por lo cual un número mayor de especies encuentran condiciones propicias y las especies de Ulva pueden afectarse, fenómeno descrito por Díaz-Piferrer (1959), para la zona. Algunas observaciones hechas en los muestreos a la zona, pueden ser considerados como es el caso de la ubicación de un extenso banco de Gracilaria dominguensis y otras algas que compiten con Ulva fasciata, es por ello que aunque exista un sustrato adecuado, la especie no manifiestó un porcentaje de cobertura alto.
Morfometría. Los valores medios de las variables morfométricas evaluadas y sus diferencias en los distintos meses y zonas, sugieren que las condiciones naturales de las tres zonas son diferentes y que las mayores tallas se alcanzan en la época de enero a junio. Se observó que generalmente en la zona aledaña al río Quibú las frondas de Ulva fasciata alcanzan las máximas longitudes de toda el área, en los meses de máximo desarrollo y en algunas ocasiones duplican o triplican tanto el largo de la ramificación más larga como el resto de sus ramificaciones; sin embargo en general se aprecian los valores más bajos del ancho de sus frondas al comparar estos resultados con el resto de las zonas. El valor medio del ancho, se mantiene estable durante todos los meses evaluados, lo que hace suponer que existen uno o varios factores que condicionan esta estabilidad y su influencia sobre dicha variable es más fuerte que la ejercida por el resto. La forma que adopta esta especie en el lugar, ya descritas por Díaz Piferrer y López en 1959, son frondas largas y estrechas rizadas en los bordes. Algunos autores han demostrado los efectos de las condiciones ambientales sobre el polimorfismo en U. fasciata (Mishigeni & Kajumulo,1979), como estrategia adaptativa de la especie a los cambios ambientales.
El valor medio obtenido para las variables del largo en los ejemplares de la zona Central, refleja que en la mayoría de los casos, no se alcanzaron valores tan altos como las frondas del Quibú y Jaimanitas en un mismo mes y profundidades similares.
El valor medio de crecimiento alcanzado por mes, en los ejemplares que crecen en el banco de Jaimanitas indica que el largo promedio de las ramificaciones en esta zona, alcanzó valores similares durante todo el tiempo de estudio. El ancho máximo de la ramificación más larga de los ejemplares que crecen en este lugar, difieren entre sí en casi todos los meses y presentó un valor mayor que las frondas del Río Quibú.
Al analizar las tallas de los especimenes en esta zona se aprecia que la época más favorable para el desarrollo es la primavera y que durante el verano cuando se producen intensas lluvias, resulta desfavorable para el desarrollo ya que se provoca el desprendimiento de los ejemplares y se cubre todo el fondo con una capa espesa de sedimento procedente del río, esta situación se presentó en el mes de junio/97, resultado que concuerda con lo planteado por Sfriso & Pavoni (1994). Este lugar por sus características de aguas tranquilas y ser una zona protegida del oleaje, posibilita que el promedio de la longitud máxima se mantenga en valores superiores a los 12 cm aun en diciembre, cuando en las otras zonas están afectadas y el valor medio de las tallas no sobrepasa los 5 cm. La especie en esta zona tiene un morfotipo diferente al descrito anteriormente y esto puede ser debido a que las condiciones ambientales donde se desarrollan estas algas no son iguales. Los datos hidroquímicos reportados por Montalvo et al. (1998) para el área, permiten confirmar la influencia de estos ríos sobre las distintas zonas en cuanto a niveles de nutrientes y fluctuaciones de la salinidad.
Las observaciones en cuanto a la diferencia de coloración de las frondas en las tres zonas, pueden fundamentarse por los resultados de las determinaciones de la concentración de clorofila en cada zona.
Cabe señalar que, las tallas pequeñas en forma de rosetas que se observaron en la zona central en el invierno es característico de las algas que crecen en lugares con gran exposición a la acción de las olas. Similares resultados fueron obtenidos por Mshigeni y Kajumulo en 1979 al comprobar los efectos del movimiento del agua sobre la morfología de las algas; estos autores plantearon que las frondas que crecen en lugares con relativa calma, muestran la forma típica de U. fasciata, mientras que la forma de roseta se presenta en lugares muy expuestos al oleaje confundiéndose a veces con la especie U. rígida.
Resulta interesante la relación inversa que se establece entre el ancho y cada una de las variables referidas al largo en los transectos ubicados en la zona aledaña al Río Quibú; posiblemente como respuesta al movimiento de agua constante que existe en el lugar, de forma tal que se produce una reducción del área de exposición de las ramificaciones, lo que les permite resistir el movimiento del agua. Sin embargo en la zona aledaña al río Jaimanitas donde las aguas son más tranquilas, debido a la morfología de la costa, se obtiene un coeficiente de correlación positivo entre todas las variables y es mayor en el caso del largo y ancho máximo, presentando en este caso una diferencia con las algas que se desarrollan cerca del río Quibú y con un morfotipo descrito como: ramificaciones menos largas pero más anchas y con bordes lisos.
Pigmentos. El espectro de absorción obtenido del extracto acetónico de U. fasciata ofreció 3 picos máximos, característicos de clorofila a, carotenoides y clorofila b, coincidiendo estos resultados con los publicados para Ulva japónica por Lobban y Harrison (1994), en los que aparece la sifoxantina de igual forma que en U. fasciata. El espectro obtenido en la muestra precipitada con carbonato de calcio indica la presencia de carotenoides en esta especie.
Hubo variación en la concentración de pigmentos (clorofila a, clorofila b y carotenoides) entre los diferentes meses, resultados muy similares a los obtenidos por Henley & Ramus, (1989 a y b), en Ulva rigida. El valor de la concentración de clorofila a y b relativamente alto en las zonas aledañas al río Quibú y al río Jaimanitas, se justifica en parte por el elevado grado de contaminación orgánica causantes de la eutroficación observada en las zonas costeras cercanas a sus desembocaduras (Montalvo, et al. 1998 y Cano, et al. 2000). Los datos hidroquímicos (amonio y nitratos) permiten caracterizar tanto los cuerpos de agua donde crece Ulva fasciata en la zona cercana a los ríos como la zona mas alejada y justifican la variación en la concentración de pigmentos en las tres zonas. Resultados similares fueron expuestos por Lahaye (1995) al estudiar la influencia de la concentración de amonio en la coloración verde intenso característica de la clorofila en Ulva rigida. Según Fillit (1995) las Chlorophyceae presentan una composición clorofílica idéntica a las fanerógramas por ser particularmente ricas en clorofila b, con valores entre 1 y 1,8 para la relación clorofila a : clorofila b. De acuerdo con lo explicado por Lobban y Harrison, (1994), la clorofila b está presente en niveles muy altos en las Ulvaceas y en otras Chlorophyceas así como en plantas superiores. Duke et al (1986) demostraron la correlación significativa inversa de la iluminación con el contenido de pigmentos en el talo.
El valor medio de la concentración de carotenoides en toda el área fue baja, siendo un poco mas elevada en la zona central, los valores hallados en este estudio se corresponden con los obtenidos por Fillit, (1995) para Ulva rígida y es conocido que estos carotenoides son pigmentos accesorios que sirven para proteger al vegetal contra altas intensidades lumínicas, en la absorción de luz y la transferencia de energía (Israel et al. 1982).
Conclusiones
La especie Ulva fasciata no se distribuye de igual forma en toda el área: el mayor porcentaje de cobertura y las mayores tallas se observaron en los meses de enero a junio y el menor porcentaje de agosto a octubre. El sustrato rocoso favorece la cobertura algal.
La morfometría de los ejemplares de Ulva fasciata difieren en los distintos ambientes naturales y las frondas mas largas y estrechas y de color verde intenso crecen cerca del río Quibú. En la zona aledaña al río Jaimanitas, tienen las frondas largas y anchas y son de color verde y las que crecen en la zona central presentan características intermedias.
Se comprobó la riqueza de pigmentos fotosintéticos de Ulva fasciata en la zona. El valor medio la clorofila b varío entre 93.20 mg/g (b.s) - 1.10 mg/g (b.s), seguidos de la clorofila a con valores que oscilaron entre 7.83 mg/g (b.s) y 0.03 mg/g (b.s) y los carotenoides (1.2 mg/g (b.s)-0.03 mg/g (b.s)). Las diferencias de concentraciones de pigmentos en las distintos ambientes y meses, justifica las diferencias de tonalidades del color verde, que se pudieron observar en el vegetal con anterioridad. Los aspectos biológicos de U. fascista abordados en este estudio, además de contribuir al conocimiento de la ecología de la especie y su relación con las condiciones ambientales, permitió obtener información valiosa para el manejo de las poblaciones naturales, con vistas a su aprovechamiento futuro como suplemento dietético o como materia prima en la industria cosmetológica, por su abundancia relativa y cantidad y calidad de pigmentos.
Se recomienda realizar otros estudios para evaluar la biomasa disponible, los componentes químicos mayoritarios, la calidad higiénica sanitaria entre otros aspectos necesarios para el manejo y aprovechamiento de este recurso en la región.
Agradecimientos
Este trabajo se realizó gracias al financiamiento del PNAP "Aprovechamiento de las Algas Marinas cubanas". Los autores desean agradecer a J. Rendón, C. Díaz, L. M. Hernández por su ayuda en el trabajo de campo y el procesamiento de las muestras en el laboratorio.
Referencias
CANO, M. 1996. Potencialidad nutricional de las algas marinas cubanas : Ulva Linnaeus y Enteromorpha Link Tesis presentada en opción del Título maestro en Biología Marina. Universidad de la Habana, Cuba. [ Links ]
CANO, M. Y L.M. HERNÁNDEZ. 1997. Distribución del género Ulva en la costa norte de la Ciudad de La Habana. Memorias del IV Congreso de Ciencias del Mar. 5 pp. [ Links ]
CANO, M.; DÍAZ, J. VALDÉS, O. Y T. CHOPIN. 2000. Distribución y cobertura de Ulva fasciata Delile y el fenómeno de eutroficación. Contribución a la Educación y la Protección Ambiental. Volumen 1: 70-73 p. [ Links ]
DE LARA ISSASSI, G. & ALVAREZ-HERNÁNDEZ S. 1994. Actividad Biológica de las algas marinas mexicanas. Rev. Hist. Nat., 45: 51-60 [ Links ]
DÍAZ PIFERRER. M, M. LÓPEZ. 1959. Taxonomía, ecología y valor nutricional de algas marinas cubanas I. ICIT. Serie de Estudios sobre Trabajos de Investigación. No.6. [ Links ]
DÍAZ-PIFERRER, M. 1964. Adiciones a la flora marina de las Antillas Holandesas, Curazao y Bonaire. Ibid. 4(4):513-543. [ Links ]
DUKE C. S. ; LAPOINTE B. E. & RAMUS J. 1986. Effects of light on growth, Rubpcasi, activity and chemical composition of Ulva species (Chlorophyta) Journal of Phycology 22: 362-370. [ Links ]
FILLIT,H. 1995. Seasonal changes in the photosynthetic capacities and pigment content of Ulva rigida in a Mediterranean Coastal Lagoon. Botánica Marina.(38): 271-280. [ Links ]
GARMENDIA, R.G. 1994. Estudio preliminar de secado en dos especies de algas marinas. Tesis de Diploma. Instituto de Farmacia y Alimentos. Univ. de la Habana, Cuba. [ Links ]
GÓMEZ, M. 1998. Variación del crecimiento y la morfología de Ulva fasciata Delile en el litoral rocoso de la C. De La Habana. Tesis de Diploma. Universidad de la Habana 46pp. [ Links ]
HAYDEN, H.S; J. BLOMSTER; C. A. MAGGS, P. C. SILVA, M.J. STANHOPE Y J.R. WAALAND. 2003. Linnaeus was right all along: Ulva and Enteromorpha are not distinct genera. Eur J. Phycol. (38): 277-294. [ Links ]
HELLEBUST J.A. & CRAIGIE JS. 1978. Handbook of Phycological Methods. Physiological & Biochemical Methods. Sponsored by the phycological Society of america Inc. Cambrigge University Press. [ Links ]
HENLEY, W.J. & RAMUS, J. 1989 A. Photoacclimation of Ulva rotundata (Chlorophyta) under natural irradiance. Mar. Biol:, 103: 261-266. [ Links ]
HENLEY, W.J. & RAMUS, J. 1989 B. Optimization of pigment content and the limits of photoacclimation for Ulva rotundata (Chlorophyta).Mar. Biol., 103: 267-274. [ Links ]
ISRAEL, A., H. ROMO, K.ALVEAL Y S. ANDRADE. 1986. Variación anual del contenido de clorofila a y carotenos totales en macroalgas bentónicas de Chile central. Anais do simpósio internacional. Algas: a energía do amanha. Instituto Oceanográfico da Universidad de Sao Paulo, 12-16 de Julho 73-78. [ Links ]
JEFFREY, S. W. & HUMPHREY, G.F. 1975. New spectrophometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phitoplankton. Biochem. Physiol. 167,191-194. [ Links ]
LAHAYE, M. 1995. Natural decoloration, composition and increase in dietary fibre content of and edible marine algae, Ulva rigida (Chlorophyta), grown under different nitrogen conditions. J.Sci: Food. agric.(68): 99-104. [ Links ]
LITTLER, D. S. Y M. M. LITTLER. 2000. Caribbean Reef Plants. An identification guide to the reef plants of the Caribbean, Bahamas, Florida and Gulf of Mexico. Offshore Graphics, Inc., 542 pp. [ Links ]
LOBBAN, C.S. Y P.J. HARRINSON. 1994. Seaweed ecology and physiology. Cambridge University Press. Cambridge. 139-241 366pp. [ Links ]
MABEAU, S. Y J. FLEURENCE. 1993. Seaweed in food products: biochemical and nutritional aspects. Tends Food Sci Technol. (4): 103-107. [ Links ]
MISHIGENI, K.S Y KAJUMULO, A. A. 1979. Effects of the environment on polymorphism in Ulva fasciata (Chlorophyta, Ulvaceae). Bot: Mar. (22):145-148. [ Links ]
MONTALVO, J.F; M.CANO; M. PERIGÓ, E. ESPONDO, M.E. CÉSAR Y C.M. PEÓN. 1998. Evaluación de factores hidroquímicos relacionados con la contaminación orgánica en tres cuerpos costeros del litoral oeste de Ciudad de La Habana. En Memorias del taller sobre medio ambiente. ISCTN. Contribución a la Educación y la Protección Ambiental Editorial Academia, La Habana: 93-96. [ Links ]
SFRISO, A. Y B. PAVONI. 1994. Macroalgae and phytoplankton competition in the central Venice Lagoon of Venice. Enviromental Technology (15): 517-528. [ Links ]
SPIEGEL, M.R. 1977. Teoría y problemas de estadística. Editorial Pueblo y Educación. Cuarta edición 358 pp. [ Links ]
SUÁREZ, A.M. 1975. Catálogo de las Algas Marinas Cubanas. Rev. De Ciencias Marinas Universidad de La Habana.
TAYLOR, W. R. 1960. Marine algae of the easthern tropical and subtropical coasts of the Americas. Univ. Mich. Press, Ann Arbor, 870 pp. [ Links ]
WYNNE, M.J. 1998. A checklist of benthic marine algae of the tropical and subtropical western atlantic: first revision. Nova Hedwigia 116: 1-155. [ Links ]

