










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.15 no.1 Ciudad de México Jun. 2005
Artículos
Caracterización diatomológica en tres cuerpos de agua de los humedales de Jilotepec-Ixtlahuaca, Estado de México
Diatoms of three bodies of water from wetlands Jilotepec-Ixtlahuaca, Estado de Mexico
María Guadalupe Oliva-Martínez1, José Gabriel Ramírez-Martínez1, Gloria Garduño-Solórzano1, Jaqueline Cañetas-Ortega2 y Martha M. Ortega3
1 Facultad de Estudios Superiores Iztacala-UNAM, Laboratorio de Botánica. Avenida de los Barrios No. 1, Tlalnepantla, Estado de México. C.P. 54090. E-mail: oliva@servidor.unam.mx
2 Instituo de Física, UNAM, Laboratorio de Microscopía Electrónica, Departamento de Física Experimental. Apartado Postal 20-364. Circuito Exterior, Ciudad Universitaria. México.
3 Instituto de Biología, UNAM, Laboratorio de Ficología. Apartado postal 70-233, Circuito exterior, Ciudad Universitaria. México.
Recibido: 16 de mayo de 2004
Aceptado: 17 de febrero de 2005
Resumen
Se representa la caracterización taxonómica y ambiental de las diatomeas de tres cuerpos de agua de los humedales de Jilotepec-Ixtlahuaca, Estado de México, a partir de seis muestreos (enero-octubre de 2001). Se colectaron crecimientos algales bentónicos y planctónicos. Los taxa se estudiaron con microscopio de luz y electrónico de barrido. Se identificaron 45 especies y 3 variedades de diatomeas distribuidas en 32 géneros. Aulacoseira granulata, Cymbella affinis, Gomphonema acuminatum, G. angustatum, Pinnularia microstauron, Synedra ulna y Stephanodiscus niagarae fueron especies dominantes. Por el contrario Craticula halophila, Cymatopleura solea y Neidium iridis fueron raras. Epithemia turgida, Eunotia curvata y Rhopalodia sp. fueron particulares del charco permanente. Achnanthes pinnata, Cyclostephanos dubius, Eunotia faba y Planothidium ellipticum son nuevos registros para la FVTM. La presencia de S. niagarae amplia su distribución como especie viviente hacia el sur de Norte América. El 89% de las especies fueron limnófilas, con tendencia a indiferentes y a reófilas. El 67% de los taxa estuvieron asociados con la diversidad de sustratos y no manifestaron marcada estacionalidad. La riqueza de especies fue mayor durante la época de secas. Craticula halophila, Encyonema caespitosum, Frustulia vulgaris y Sellaphora pupula se encontraron en época de lluvias. Todas las especies fueron meso-euritermales con afinidad templada. El pH fue ligeramente básico (7.8), el 65% de las especies fueron alcalífilas. Eunotia faba, acidófila-circumneutral, E. curvata y Surirella tenera circumneutral-indiferentes. Stephanodiscus niagarae en asociación con Aulacoseira granulata, Fragilaria capucina y F. crotonensis, indican una tendencia a condiciones eutróficas. Todas las especies son características de agua dulce y oligohalobias. Los resultados de las variables ambientales medidas indicaron que el agua del área de estudio es poco mineralizada.
Palabras clave : Microalgas, diatomeas, humedales, bentos, plancton, microscopio electrónico de barrido, México, Faja Volcánica Transmexicana.
Abstract
The taxonomic and environmental characteristics of the diatom flora found in three bodies of water in the wetlands of Jilotepec-Ixtlahuaca, Estado de Mexico are described. Six samplings were made (January-October, 2001). Benthic and planktonic diatoms growth were collected.The samples were analyzed with a Light Microscope (LM) and a Scanning Electron Microscope (SEM). 45 species and 3 infraspecific taxa belonging to 32 genera were identified. Aulacoseira granulata, Cymbella affinis, Gomphonema acuminatum, G. angustatum, Pinnularia microstauron, Synedra ulna y Stephanodiscus niagarae were the dominat species. Craticula halophila, Cymatopleura solea, and Neidium iridis were rare species. Epithemia turgida, Eunotia curvata and Rhopalodia sp. were species specific to the permanent pond. Achnanthes pinnata, Cyclostephanos dubius, Eunotia faba and Planothidium ellipticum are new records for theTrans Mexican Volcanic Belt. We found a living population of S. niagarae from wetlands, suggesting that this species had once a much wider distribution towards the southern region of North America. 89% of the species were limnophilous with an indifferent and rheophilous tendency. 67% of the taxa were associated to the diversity of substrates and did not exhibit marked seasonality. Species richness was higher during dry season. Craticula halophila, Encyonema caespitosum, Frustulia vulgaris y Sellaphora pupula were found during the rainy season. All of the species were found to be meso-eurytherms with temperate affinity. pH was slightly basic (7.8), 65% of the species were alkaliphilous. Eunotia faba, acidophilous-circumneutral, Eunotia curvata and Surirella tenera circumneutral-indifferent. Stephanodiscus niagarae in association with Aulacoseira granulata, Fragilaria capucina and F. crotonensis showed a tendency to eutrophic condition. All of the species were characterized as being from freshwater and oligohalobous. Results from environmental analysis show that the water from study area has a low mineral content.
Key words : Microalgae, diatoms, wetlands, benthos, plankton, scanning, electron microscope, Mexico, Trans Mexican Volcanic Belt.
Introducción
De todas las regiones biológicas en que México está dividido, la Faja Volcánica Transmexicana (FVTM) es una de las zonas más importantes por la biodiversidad que alberga. Esta provincia morfotectónica atraviesa al país de costa a costa por su parte central desde el Golfo de México al este y el Océano Pacífico al oeste, entre los paralelos 17° 30' y 20° 25' de latitud norte y los meridianos 96° 20' y 105° 20' de longitud oeste. Es una franja alargada de unos 900 km con amplitudes variables de 50 a 250 km, cubriendo parte de los estados de Veracruz, Puebla, Tlaxcala, Hidalgo, Estado de México, Distrito Federal, Jalisco, Michoacán, Colima y Nayarit (Ferrusquía-Villafranca, 1993). Su origen se debe a una recurrente y continua actividad volcánica que tiene lugar en el Plio-cuaternario. Morfológicamente la FVTM está formada por una serie de planicies escalonadas, pequeños grupos de volcanes y grandes cadenas montañosas volcánicas (Lugo, 1984). Sus máximas elevaciones se ubican entre los paralelos 19° y 20° de latitud norte donde se levantan los picos montañosos más altos del país, entre ellos los volcanes Pico de Orizaba, Popo-catépetl, Iztaccíhuatl, Nevado de Toluca y Nevado de Colima (Reyes-Matamoros y Martínez-Moreno, 2003). La infiltración del agua subterránea se presenta en las zonas fracturadas de las cadenas montañosas y circula hasta su acumulación en los valles y zonas lacustres (Arriaga et al., 2000). La Faja Volcánica Transmexicana es la zona de contacto y transición de las dos regiones biogeográficas del Continente Americano: Neártica y Neotropical. Su compleja topografía, variabilidad de altitudes, climas, posición geográfica del país y su historia geológica provee un mosaico de hábitats y microhábitats que la hace uno de los principales centros de endemismos y riqueza de especies de algunos organismos tales como mamíferos, encinos y otros grupos de plantas (Fa y Morales, 1993; Nixon, 1993; Rzedowski, 1993). En la FVTM se localizan importantes ambientes acuáticos conocidos como humedales, en los últimos años ha aumentado notablemente el interés por su estudio, estas áreas que se encuentran inundadas temporal o permanentemente por aguas dulces o salinas, son de régimen natural o artificial, cuya profundidad no excede de seis metros (Ramsar,1999). Los humedales son ecosistemas complejos, dinámicos y altamente productivos, son áreas de crianza y reproducción de un número elevado de flora y fauna silvestre, reconocidos por su alto número de endemismos, especialmente de peces e invertebrados y porque dan refugio y protección a una gran cantidad de especies de aves migratorias. Estos ambientes también desempeñan un papel ecológico importante en el control de la erosión, sedimentación, inundaciones, mantenimiento de pesquerías, desarrollo agrícola y turístico (Ramsar, 1998,1999). En los humedales las épocas seca y anegada, tienen rasgos muy diferentes en el panorama vegetal, tanto que pueden considerarse una transición entre sistemas terrestres y acuáticos. En estos la persistencia del sistema terrestre o anegado es variable y depende de la topografía del terreno, del grado de permeabilidad del suelo, del origen del aporte principal del agua y de la lluvia. Las algas perifíticas proliferan abundantemente en estas condiciones, sobre o alrededor de otras plantas, formando masas superpuestas al sustrato, siempre libres o formando costras quebradizas. Los crecimientos algales desempeñan un papel preponderante porque regulan el paso de nutrimentos entre el sustrato, los sedimentos y el agua, asimilan nutrimentos y los liberan al sustrato, de este modo otros organismos dependen de las algas para la obtención de nutrimentos (Goldsborough y Robinson, 1996; Novelo y Tavera, 1999).
Actualmente el estudio de la ficoflora de los ambientes continentales se ha incrementado, sin embargo, aún quedan áreas que no se conocen en absoluto. La alteración de estos ambientes y su contaminación son algunas de las causas que disminuyen la diversidad y abundancia de la ficoflora de México, por lo que es de fundamental importancia realizar estudios monográficos y ecológicos que coadyuven a la búsqueda de especies propias de los cuerpos de agua continentales mexicanos. Tradicionalmente las investigaciones que se realizan sobre humedales, se centran en el estudio de las aves o de algún otro grupo animal como peces y mamíferos, quedando en segundo término las plantas y las algas (Cantoral-Uriza y Aboal, 2001). De manera particular en México, las diatomeas han sido motivo de distintas investigaciones que abarcan aspectos de taxonomía e información ambiental como los realizados por Lozano-García et al. (1993), Valadez et al. (1996), Cantoral-Uriza et al. (1997), Ibarra-Vázquez y Novelo (1997), Vilaclara et al. (1997), García-Rodríguez y Tavera (1998), Rico et al. (1998), Oliva et al. (2001) y Ramírez-Vázquez et al. (2001). Existe muy poca información de las diatomeas de los humedales mexicanos, como el trabajo de Ibarra-Vázquez y Novelo (1997) quienes estudiaron la zona ubicada en la Reserva Ecológica "El Edén" Quintana Roo y en la que registraron 21 géneros y 60 especies.
Este trabajo forma parte del proyecto "Algas Epicontinentales de la Faja Volcánica Transmexicana". En dicha región desde 1854 a 2001 se han citado 472 especies de diatomeas pertenecientes a 77 géneros (Ramírez, 2003). En esta contribución se presenta la caracterización taxonómica y ambiental de las diatomeas de tres cuerpos de agua de los humedales de Jilotepec-Ixtlahuaca, Estado de México, región hidrológica considerada por la CONABIO como prioritaria por su alta biodiversidad (Arriaga et al., 1998, 2000). Hasta el momento no existen para esta zona publicaciones relacionadas con el estudio de este grupo algal.
Área de estudio
Los Humedales de Jilotepec-Ixtlahuaca se localizan hacia el norte del Estado de México a una altura de 2560 msnm, las coordenadas que lo delimitan son 20°06'00" y 19º40'12" latitud N y 99º51'00" y 99º19'12" longitud O (Fig. 1). Su composición geológica colindante es el basalto, arenisca y suelo aluvial (CETENAL, 1974, 1975), con suelos de tipo Luvisol, Planosol, Feozem y Andosol, con pendientes, obstrucciones e inundaciones. El clima de la región es templado subhúmedo, semifrío subhúmedo con lluvias en verano y parte del otoño. El Intervalo de temperatura media anual es de 10-16°C. Precipitación total anual de 700-1200 mm. La vegetación de la zona corresponde a bosques de pino-encino y pino, pastizal inducido y tular. El uso del suelo en sus alrededores es la acuicultura, ganadería y agricultura de riego y temporal, presentando una erosión hídrica fuerte (Arriaga et al., 1998)
Materiales y métodos
La colecta en los Humedales de Jilotepec-Ixtlahuaca se realizó de enero a octubre del 2001. Se designaron tres sitios (Fig. 1): la zona litoral del embalse Danxhó localizada a los 19º52'42" latitud N y 99º33'28" longitud O. El embalse tiene una superficie aproximada de 311 ha y una capacidad de almacenamiento de 22.7 Mm3. Pertenece a la región hidrológica del Pánuco, su corriente principal se deriva del río Coscomate, así como una gran cantidad de agua pluvial que se canaliza hacia el cuerpo de agua (Secretaría de Ecología, 2003). El canal Santa Elena localizado a los 19º53'20" latitud N y 99º33'40" longitud O, se caracteriza por ser de corriente muy baja (< 0.1 m/seg casi estancada), baja (0.1-0.35 m/seg) a moderada (0.35- 0.6 m/seg) que varia con la época del año (Carmona-Jiménez y Montejano-Zurita, 1993). Su profundidad es escasa teniendo como máxima 0.90 m y como mínima 0.20 m y 2 m de ancho aproximadamente. El canal corre paralelo al embalse Danxhó, en algunas zonas se encuentran rocas emergentes y cantos rodados cubiertos por lodo y musgos. En sus orillas se desarrollan hidrófitas emergentes como Eleo-charis densa y Ludwigia palustris e hidrófitas enraizadas sumergidas como Myriophyllum aquaticum y Potamogeton no-dosus (García-Calderón y Rzedowski, 2001). El charco permanente localizado al noroeste del embalse a los 19º55'53" latitud N y 99º33'59" longitud O, tiene una superficie aproximada de 9000 m2 y una profundidad de 0.50 -1.00 m que varía con la época del año. La vegetación circundante principalmente constituida por Typha latifolia y otras hidrófitas enraizadas emergentes como Juncus effusus, Ludwigia peploides, Nynphoides fallax, Sagittaria latifolia y el helecho acuático Marsilea mollis, además de Potamogeton nodosus, hidrófita enraizada sumergida (García-Calderón y Rzedowski, 2001). Para su ubicación se utilizó el geoposicionador Magellan, modelo Meridian y apoyo cartográfico (CETENAL, 1974, 1975).
En el campo se colectaron crecimientos algales bentónicos visibles (Goldsborough y Robinson, 1996; Novelo y Tavera, 1999): alrededor de macrófitas (perifiton), sobre macrófitas (epifiton), masas sobrepuestas al sustrato (plocon), sobre de algas como Spirogyra y Oedogonium que no están directamente adheridas al sustrato (metafiton) y organismos libremente suspendidos en la columna de agua (plancton), con la ayuda de una espátula y mediante el filtrado de 20 l de agua de cada localidad, utilizando una red manual de fitoplancton con abertura de malla de 25 µm. Las muestras se fijaron con formol a una concentración final del 4% (Lara et al.1996). Se midieron temperatura ambiental y en el nivel superficial (30 cm de profundidad) in situ, los siguientes parámetros físicos y químicos del agua: visibilidad del disco de Secchi, temperatura y pH con un potenciómetro Cole-Parmer Digi-Sense Modelo 5985, oxígeno disuelto con la técnica de Winkler modificada, alcalinidad total y dureza total con métodos volumétricos (A. P. H. A. et al., 1985). La conductividad específica (K25) se midió con un conductímetro Conductronic CL35. Para el estudio microscópico de las muestras se realizó la técnica de limpieza de frústulas de diatomeas (Hasle y Fryxell, 1970). Con las muestras libres de materia orgánica se siguieron dos técnicas para su observación: En la primera se elaboraron preparaciones permanentes, utilizando Nafrax® como medio de montaje cuyo índice de refracción es de 1.7 ± 0.02 (Hasle y Fryxell, 1970) y se observaron en un Microscopio de Luz (ML) Lobophot-2 marca NIKON con cámara fotográfica integrada. El remanente del material limpio fue montado sobre cinta de doble cara en bases de aluminio y recubierto con plata pura 0.999, se observaron en un Microscopio Electrónico de Barrido (MEB) JSM-5200 marca JEOL, con una distancia de trabajo de 10 mm y 25 kV. Las muestras líquidas se incorporaron al herbario IZTA con los números de registro 726-895 y las laminillas permanentes con los números 1465-1468.
El porcentaje de presencia y distribución de las especies en los tres sitios estudiados se obtuvo utilizando el criterio de Braun-Blanquet (1979) modificado: dominantes (con más del 60% de presencia), raras (menos del 15%) y las particulares que solo se encontraron en el charco permanente.
El sistema de clasificación que se siguió fue el propuesto por Round et al. (1990). La terminología usada en la descripción de las especies fue la de Ross et al. (1979) y Round et al. (1990). La validez de los nombres científicos y su sinonomia se verificó con Van Landingham (1967-1979) y las páginas electrónicas Integrated Taxonomic Information System (2002) y con el Index Nominum Algarum (2000). La caracterización ambiental de las diatomeas determinadas, se hizo por su comparación con los criterios propuestos por Lowe (1974), Patrick (1977), Gasse (1986), Foged (1993), Håkansson (1993), Yang y Dickman (1993), Goldsborough y Robinson (1996) y Novelo y Tavera (1999) (Tabla 1).
Resultados
Se determinaron 45 especies y 3 variedades de diatomeas distribuidas en 32 géneros. Se presenta la lista sistemática de las especies (Apéndice). En la tabla 2 se señala su distribución y porcentaje de presencia, a través de los seis muestreos en las tres localidades estudiadas: embalse (Em), canal (Ca), y charco permanente (Ch). En la tabla 3 se muestran las especies dominantes, las raras y las particulares para el charco permanente. De las especies anteriores se hace su descripción, así como también la de las especies registradas por primera vez para la Faja Volcánica Transmexicana (FVTM) que se indican con un asterisco: Achnanthes pinnata Hustedt, Cyclostephanos dubius (Fricke) Round, Eunotia faba Ehrenberg y Planothidium ellipti-cum (Cleve) Round et Bukhtiyarova. Se señala el hábitat, forma de vida, sinonimia, los registros referidos para la FVTM, información ambiental citada en la literatura y las referencias bibliográficas utilizadas para su determinación.
Descripción de Especies
* Achnanthes pinnata Hustedt (Figs. 2-4)

Valvas lineales o lanceoladas, con ápices redondeados. Estrías paralelas formadas por una hilera de areolas (Figs. 2 y 4). En la vista externa los extremos centrales del rafe son rectos (Fig. 4), los extremos terminales son curvados (Fig. 3). La valva sin rafe no fue observada. Medidas de la valva: eje api-cal de 6.4-10.4 µm de largo, eje transapical de 3.2-4 µm de ancho. Densidad de estrías: 14 estrías en 5 µm.
Muestras de herbario: 772, 781.
Hábitat: embalse y canal.
Forma de vida: creciendo en natas flotantes de Oedogonium y Spirogyra (metafiton).
*Nuevo Registro para la FVTM.
Información ambiental citada en la literatura: pH: alcalifila (?), Hábitat general: arroyos y ríos. Distribución geográ fica: Sur, Sureste y Este de los estados centrales de los Estados Unidos; California (Patrick y Reimer, 1966).
Referencias: Patrick y Reimer, 1966, p. 266, Lám. 17, figs. 17-18; Lange-Bertalot y Krammer, 1989, p.119, fig. 32:19-30.
Aulacoseira granulata (Ehrenberg) Simonsen (Figs. 5-9)
Células con varios plastos discoidales, cilíndricas formando cadenas largas o cortas, rectas, la unión entre las val-vas vecinas es mediante espinas que se encuentran en el borde de la cara valvar circular, poros sencillos y esparcidos con frecuencia hacia la periferia (Figs. 5 y 6), las espinas están ensanchadas en su base y se incrustan con las de la val-va adyacente (Fig. 8). Una a dos espinas largas de 9 -21.6 µm (Fig. 7). Las espinas largas se acoplan en surcos longitudinales que se encuentran entre las líneas de las areolas del manto de la célula vecina (Fig. 9). Las areolas del manto están dispuestas en hileras transversales o longitudinales a lo largo del manto, con una densidad de 10-15 en 10 µm (Fig. 9). Medidas de la valva: 8.5-24 µm de largo, 4.5-10.4 de diámetro. Especie dominante.
Muestras de Herbario: 726, 729, 730, 731, 734, 786.
Hábitat: Embalse, canal y charco.
Forma de vida: Crece en sustrato arcilloso, sobre y alrededor de vegetación acuática sumergida como Potamogeton nodosus y Myriophyllum aquaticum, natas flotantes de Oedo-gonium y Spirogyra y en la columna de agua (plocon, perifiton, epifiton, metafiton y plancton).
Sinonimia: Gaillonella granulata Ehrenberg, Melosira granulata (Ehrenberg) Kützing, Melosira granulata var. granulata (Ehrenberg) Ralfs.
Registros para la FVTM: DISTRITO FEDERAL: Chapulte-pec (Ortega et al., 1994); Xochimilco (Ortega et al., 1994); Río la Magdalena, Dinamo 4 y Camino Santa Teresa (Ramírez-Vázquez et al., 2001). ESTADO DE MÉXICO: Lago de Texcoco (Díaz-Lozano, 1917; Bradbury, 1971); Presa de Guadalupe (Lugo et al.,1998). GUANAJUATO: Embalse Ignacio Allende (Ló-pez-López y Serna-Hernández, 1999). MICHOACÁN: Lago de Pátzcuaro (Osorio-Tafall, 1941, 1941a; Rosas et al., 1993; Sládecék y Vilaclara, 1993; Ortega et al., 1994); Laguna Zacapu (Metcalfe, 1988). MORELOS: Laguna de Zempoala (Kusel-Fetz-mann, 1973).
Información ambiental citada en la literatura: pH: alcalifila, oscila entre 6.3-9, óptimo 7.9 - 8.2. Estado Trófico: eutrófica. Halobionte: oligohalobia (indiferente). Saprobionte: oligosapróbica a mesosapróbica. Corriente: limnófila. Hábitat general: lagos, estanques y ríos. Forma de Vida: euplanctóni-ca. Distribución estacional: verano. Distribución Geográfica: cosmopolita (Lowe, 1974; Foged, 1993; Håkansson, 1993; Yang y Dickman, 1993), con amplia distribución al Este de Africa (Gasse,1986).
Referencias: Hustedt, 1930, p. 87, fig. 44, como Melosira granulata; Germain, 1981, p. 24, Lám. 3, figs. 1-5; Gasse, 1986, p. 77, Lám. 1, figs. 5,8, como Melosira granulata; Krammer y Lange-Bertalot, 1991, p. 22-24. fig. 16:1-2, fig. 17:1-10, fig. 18:1-12, fig.19: 9.
Craticula halophila (Grunow) Mann (Fig. 10)
Valvas lanceoladas con ápices rostrados, área axial estrecha y recta, área central no diferenciada del área axial. Estrías finas ordenadas de una forma paralela al eje apical, rafe recto con nódulos centrales muy evidentes (Fig. 10). Medidas de la valva: eje apical de 23.2-118 µm de largo, eje transapical de 5.6-26.4 µm de ancho. Densidad de estrías: 13-20 estrías en 10 µm. Especie rara.
Muestras de Herbario: 839, 845.
Hábitat: Embalse y canal.
Forma de vida: Crece alrededor de vegetación acuática sumergida o enraizada emergente como Myriophyllum aqua-ticum y Eleocharis densa respectivamente y en sustrato arcilloso (perifíton y plocon).
Sinonimia: Navicula halophila (Grunow) Cleve, Navicula cuspidata var. halophila Grunow.
Registros para la FVTM: ESTADO DE MÉXICO: Lago de Texcoco (Bradbury, 1971). MICHOACÁN: Laguna Zacapu (Metcalfe, 1988).
Información ambiental citada en la literatura: pH : alcalifila, oscila entre 6.7- 7.8. Halobionte: mesohalobia. Corriente: limnófila. Hábitat general: manantiales, lagos y ríos. Temperatura: euritérmica. Distribución Geográfica: cosmopolita, Sur de Carolina, Ohio, Texas Wyoming (Patrick y Reimer, 1966). Es común encontrarla en agua dulce, pero prefiere aguas con alta concentración mineral e indiferente a la composición aniónica (Lowe, 1974; Germain, 1981; Gasse, 1986).
Referencias: Hustedt 1930, p. 268, fig. 436; Germain, 1981, p. 172, Lám. 65, figs. 1-5; Gasse, 1986, p. 98, Lám. 16, fig. 5, como Navicula halophila; Mann y Stickle, 1991, p. 79-107, figs. 17-23.
* Cyclostephanos dubius (Fricke) Round (Figs. 11-17)

Valvas circulares, concéntricamente onduladas. La superficie de la valva con fascículos e interfascículos radiales. Areolas dispuestas en un patrón radial que se inicia desde el centro llegando a tener de dos a tres hileras hacia la unión del manto y la valva, las areolas en el centro tienen un arreglo irregular (Figs. 11 y 12). Las espinas cuando están presentes, se encuentran en los interfascículos cerca del márgen de la valva (Fig. 11). Dos o tres fultopórtulas centrales más pequeñas que las areolas (Fig. 12, flecha), internamente con dos poros satélite (Fig. 14, FPCI). Fultopórtulas marginales externas debajo de las espinas y cerca del margen de la valva (Fig 13, FPME), internamente con dos poros satélite (Fig. 15, FP-MI). La abertura externa de la única rimopórtula se encuentra cerca del manto, en un interfascículo como una hendidura (Fig. 12, RP flecha) e internamente como un labio oblicuo (Fig. 16, RP). La vista interna de la valva presenta cámaras alveolares no bien definidas (Fig. 17). Diámetro de las valvas: 5.4-15.2 µm. Densidad de areolas marginales: 13-15 en 5 µm.
Muestras de Herbario: 738, 741, 743, 755, 758.
Hábitat: Embalse, canal y charco.
Forma de vida: Crece en natas flotantes de Oedogonium, en sustrato arcilloso y en la columna de agua (metafiton, plocon y plancton).
Sinonimia: Cyclotella dubia Fricke, Stephanodiscus dubius (Fricke) Hustedt, Stephanodiscus pulcherrimus Cleve-Euler
*Nuevo registro para la FVTM
Información ambiental citada en la literatura: pH: alcalífila, oscila ente 6.8 - 7.4. Estado Trófico: eutrófica. Halobionte: oligohalobia/halófila. Hábitat general: lagos. Forma de Vida: planctónica y sedimentos. Distribución estacional: primavera, verano y otoño. Distribución Geográfica: cosmopolita, ha sido citada raramente para Norte América, ampliamente distribuida en Europa y al Sur de Africa, es común encontrarla en aguas con alta concentración de sílice y elevada conductividad (Hickel y Håkansson, 1987; Pienaar y Pieterse, 1990; Krammer y Lange-Bertalot, 1991).
Referencias: Hickel y Håkansson, 1987, p. 35-39. figs. 9 -18; Pienaar y Pieterse, 1990, p. 202-203, figs. 1-5; Krammer y Lange-Bertalot, 1991, p. 64-65, fig. 42:1, 2; fig. 67:7-9b; Håkansson, 2002, p. 62, figs. 198-208.
Cymatopleura solea (Brébisson) W. Smith (Figs .18 y 19)
Valvas isopolares, contorno elíptico, estrechas al centro, ápices agudos (Fig.18). El margen de la valva un poco ensanchado, muestra el canal del rafe (Fig. 19). Frecuentemente el eje apical esta marcado por una zona estrecha, lisa y rectilínea, dando lugar a un pseudorrafe. La superficie de la val-va es ondulada, con estrías transversales muy finas (Fig. 19). Medidas de la valva: eje apical de 67-94 µm de largo, eje tran-sapical de 19-22 µm en la parte más ancha y de 16-20 en la parte más estrecha, 8 fíbulas en 10 µm, con 6 crestas y 5 valles (Fig. 18). Especie rara.
Muestras de Herbario: 839, 840,
Hábitat: Embalse y charco.
Forma de vida: Crece en natas flotantes de Oedogonium y Spirogyra y en sustrato arcilloso (metafiton y plocon).
Sinonimia: Cymatopleura librile (Ehrenberg) Pantocksek
Registros para la FVTM: ESTADO DE MÉXICO: Texcoco, Cerro Soltepec (Díaz-Lozano, 1917); Lago de Texcoco (Brad-bury, 1971); Río Lerma (Ortega et al.,1994). MICHOACÁN: Laguna Zacapu (Metcalfe, 1988).TLAXCALA: Paleolago Tlaxcala, Mina Santa Bárbara (Rico et al., 1998).
Información ambiental citada en la literatura: pH: alca-lífila, oscila ente 5.4 - 9.0, óptimo arriba de 8. Estado Trófico: eutrófica. Halobionte: oligohalobia (indiferente). Saprobionte: oligosapróbica a beta-mesosapróbica. Hábitat general: lagos y ríos. Forma de Vida: bentónica, perifítica, epifítica, planctó-nica. Distribución estacional: otoño e invierno. Temperatura: de oligotérmica (0 - 15°C) a euritérmica (15 - 30.5°C). Distribución Geográfica: cosmopolita, Europa, Africa, India (Lowe, 1974; Sarode y Kamat, 1984; Gasee, 1986; Foged, 1993).
Referencias: Hustedt, 1930, p. 425, fig. 823 a; Vinyard, 1979, p. 58, fig.177 a, b; Germain, 1981, p. 374, Lám. 141, figs. 1-8; Sarode y Kamat, 1984, p. 228. Lám. 27, fig. 636; Gasee, 1986, p. 39, Lám. 44, figs. 3-4, como Cymatopleura librile; Krammer y Lange-Bertalot, 1988, p. 168, fig.116:1-4, 117: 1-5, 118: 1-8.
Cymbella affinis Kützing (Figs. 20-23) (21, 22)

Valvas dorsiventrales, margen ventral ligeramente convexo, con extremos, redondeados, área axial angosta (Figs. 20 y 21). Área central ligeramente redondeada. Estrías uniseriadas, en la vista externa compuestas de poros alargados (Figs. 20, 22 y 23). En especímenes con pie mucilaginoso, ambos ápices presentan una área discreta, compuesta de pequeños poros, a través de los cuales el material del tallo es excretado (Fig. 22, flecha). El rafe lateral se localiza a lo largo de la cara valvar, cerca de la línea media de la valva y curvado en forma dorsiventral (Figs. 21 y 22). Los extremos centrales del rafe están formados por ganchos que apuntan hacia el margen dorsal (Fig. 23), mientras que los extremos terminales se doblan hacia el margen ventral (Fig. 22). Uno o dos estigmas alargados se ubican en la región central del rafe (Figs. 20 y 23). Medidas de la valva: eje apical de 20-37 µm de largo, eje transapical de 6.5-10 µm de ancho. Densidad de estrías: 10-13 en 10 µm. Especie dominante.
Muestras de Herbario: 726, 727, 728, 729, 730.
Hábitat: Embalse, canal y charco.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Juncus effusus, Eleocharis densa, natas flotantes de Oedogonium y Spirogyra y en sustrato arcilloso (perifiton, epifiton, metafiton y plocon).
Sinonimia: Cymbella excisa Kützing, Cocconema par-vum W. Smith, Cymbella parva (W.Smith) Kirchner.
Registros para la FVTM: Zona Central de México (Met-calfe, 1988).
Información ambiental citada en la literatura: pH: alcalífila a alcalibióntica, oscila ente 4.3 - 9.0, óptimo 7.8 - 8.5. Ha-lobionte: oligohalobia (indiferente). Saprobionte: saprofóbica a oligosapróbica. Hábitat general: lagos, ríos y estanques. Forma de Vida: bentónica, epilítica, epifítica. Distribución estacional: verano. Distribución Geográfica: cosmopolita, Europa, Africa, India, se desarrolla mejor en tierras cálidas (Lowe, 1974; Patrick y Reimer, 1975; Sarode y Kamat, 1984; Gasee, 1986; Foged, 1993; Krammer, 2002).
Referencias: Hustedt, 1930, p. 362, fig. 671 a, b; Patrick y Reimer, 1975, p. 57, Lám. 10, fig. 7; Germain, 1981, p. 282, Lám. 104, figs. 1-11; Sarode y Kamat, 1984, p. 166. Lám. 19, fig. 440; Maidana, 1985, p. 107, Lám. 7, fig. 11, Lám. 30, figs. 3-5; Gasee, 1986, p. 40, Lám. 27, fig. 20; Novelo, 1998, p. 393, fig. 291; Kram-mer, 2002, p. 40. Lám. 21, figs. 1-21; Lám. 22, figs. 1-20; Lám. 23, figs. 1-18.
Epithemia turgida (Ehrenberg) Kützing (Fig. 24)
Valva isopolar, vista valvar curvada, con polos redondeados o capitados, margen dorsal convexo y ventral cóncavo (Fig. 24). Rafe arqueado en forma de V con el nódulo central en el ápice y dirigido hacia el margen dorsal (Fig. 24). La superficie valvar esta recorrida por costillas transversales, entre las costillas se observan dos o tres hileras de estrías finas y delgadas, con areolas visibles (Fig. 24). Medidas de las valvas: eje apical de 44.8 - 83.2 µm de largo, eje transa-pical de 8.8 -20.2 µm de ancho. Densidad de costillas de 3-4 en 10 µm. Densidad de estrías 12-15 en 10 µm. Especie particular del charco.
Muestras de Herbario: 761, 762, 763, 842.
Hábitat: charco permanente.
Forma de vida: Crece sobre sustrato arcilloso y natas flotantes de Oedogonium y Spirogyra (plocon y metafiton).
Sinonimia: Navicula turgida Ehrenberg, Frustulia pincta Kützing, Eunotia turgida Ehrenberg, Epithemia pinctum (Küt-zing) Brébisson.
Registros para la FVTM: ESTADO DE MÉXICO: Texcoco, Cerro Soltepec (Díaz-Lozano,1917); Lago de Texcoco (Bradbury, 1971); Laguna de Victoria (Mendoza-González, 1985; Ortega et al.,1994). MORELOS-ESTADO DE MÉXICO: Laguna de Zempoala (Kuzel-Fetzmann, 1973; Ortega et al., 1994). TLAX-CALA: Paleolago Tlaxcala, Mina Santa Bárbara (Rico et al., 1998); Zona Central de México (Metcalfe, 1988).
Información ambiental citada en la literatura: pH: alcalífila a alcalibióntica, oscila ente 4.6 - 9.0, óptimo alrededor de 8.2. Halobionte: oligohalobia (indiferente). Saprobionte: saprógena a beta-mesosapróbica. Corriente: limnófila. Hábitat general: lagos, ríos y estanques. Forma de Vida: bentónica, perifítica, epifítica. Distribución Geográfica: cosmopolita, Europa, Estados Unidos (Lowe, 1974; Patrick y Reimer, 1975).
Referencias: Hustedt, 1930, p. 387, fig. 733; Patrick y Reimer, 1975, p. 182, Lám. 25, figs.1 a, b; Germain, 1981, p. 318, Lám. 118, figs. 1 - 4; Novelo, 1998, p. 450, fig. 354.
Eunotia curvata (Kützing) Lagerstedt (Fig. 25)
Valvas ligeramente arqueadas, isopolares, ápices redondeados, margen ventral recto o ligeramente cóncavo, con nódulos polares pequeños. Superficie valvar finamente estriada (Fig. 25). Estrías punteadas o lineadas perpendiculares al eje longitudinal recurvado. Rafe corto poco visible en los nódulos polares. Medidas de las valvas: eje apical de 49-76 µm de largo, eje transapical de 3-4 µm de ancho. Densidad de estrías: 13-16 estrías en 10 µm. Especie particular del charco.
Muestras de Herbario: 876, 880, 883.
Habitat: charco permanente.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, natas flotantes de Oedogonium y Spirogyra y en la columna de agua (perifiton, epifiton, metafiton y plancton).
Sinonimia: Eunotia bilunaris (Ehrenberg) Mills, Eunotia lunaris (Ehrenberg) Grunow.
Registros para la FVTM: ESTADO DE MÉXICO: Lago de Texcoco (Bradbury, 1971). HIDALGO: Mineral del Monte: Cerca de Real del Monte (Ortega et al., 1994). Zona Central de México (Metcalfe, 1988).
Información ambiental citada en la literatura: pH: acidófila a indiferente, oscila ente 4.0 - 9.0, óptimo 5.5 - 6.0. Estado Trófico: eutrófica a oligotrófica. Halobionte: indiferente a halofóbica. Saprobionte: oligosapróbica. Hábitat general: lagos, estanques, charcos, rara en arroyos, ríos. Forma de Vida: bentónica, perifítica, epifítica, ticoplanctónica. Distribución Geográfica: cosmopolita, ampliamente distribuida en aguas de bajo contenido mineral, indiferente al calcio. Europa, Estados Unidos. Este de Africa (Lowe, 1974; Patrick y Reimer, 1966; Sarode y Kamat, 1984; Gasse, 1986; Alles et al.,1991).
Referencias: Hustedt, 1930, p. 183, fig. 249, como Euno-tia lunaris; Patrick y Reimer, 1966, p. 189, Lám. 10, fig.4; Germain, 1981, p. 94, Lám. 32, figs. 7 -10, como Eunotia lunaris; Maidana, 1985, p. 67. Lám. 3, fig. 7, como Eunotia lunaris; Gasse, 1986, p. 52, Lám. 11, figs. 11-12; Novelo, 1998, p. 301, fig. 197, como Eunotia bilunaris.
*Eunotia faba Ehrenberg (Fig. 26)
Valvas ligeramente arqueadas, isopolares, ápices redondeados, margen ventral ligeramente recto, margen dorsal convexo, con nódulos polares subapicales. Superficie valvar finamente estriada (Fig. 26). Estrías lineadas perpendiculares al eje longitudinal recurvado. Medidas de las valvas: eje api-cal de 29-35.2 µm de largo, eje transapical de 4.6-7 µm de ancho. Densidad de estrías: 12-14 estrías en 10 µm.
Muestras de Herbario: 810, 816, 883.
Hábitat: canal y charco permanente.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, Myriophyllum aquaticum, natas flotantes de Oedogonium y Spirogyra y en la columna de agua (perifiton, epifiton, metafiton y plancton).
Sinonimia: Himantidium faba Ehrenberg, Himantidium soleirolii W. Smith non Kützing, Eunotia vanheurckii Patrick.
*Nuevo registro para la FVTM.
Información ambiental citada en la literatura: pH: acidófila a circumneutral, oscila ente 4.0 -9.0, óptimo 5.5-6.0. Estado Trófico: oligotrófica. Halobionte: indiferente a halofóbica. Saprobionte: oligosapróbica. Corriente: limnófila. Hábitat general: lagos, estanques, ciénagas. Forma de Vida: bentónica, perifítica, Distribución Geográfica: cosmopolita, Europa Central, Estados Unidos, Francia (Patrick y Reimer, 1966; Germain,1981; Krammer y Lange-Bertalot, 1991; Håkansson,1993; Alles et al., 1991).
Referencias: Hustedt, 1930, p. 183, fig. 246; Patrick y Reimer, 1966, p. 210-211, Lám. 13, fig. 7, como Eunotia vanheurckii var. vanheurckii; Germain, 1981, p. 92, Lám. 31, figs. 20-21; Krammer y Lange-Bertalot, 1991, p. 225. fig. 164: 1-10; Novelo, 1998, p. 302, fig. 198.
Gomphonema acuminatum Ehrenberg (Figs. 27-30)
Valvas heteropolares, ápice capitado, más ancho que el centro de la valva, rostrado, ápice inferior redondeado (Figs. 27 y 28). Área axial lineal, angosta y la central redondeada con un estigma sobre uno de sus lados (Fig. 29). Estrías paralelas uniseriadas (Figs. 28 y 29), en algunos especímenes con dos hileras de areolas. La ornamentación del cíngulo consiste de pocas bandas, con una o dos líneas transversales de poros (Fig. 30, flecha). Medidas de las valvas: eje apical de 38.4-52.8 µm de largo, eje transapical en la porción más ancha de 12-15.2 µm, de 9.6-12 µm en el centro de la valva y de 5-6 µm en la base. Densidad de estrías: 10-13 en 10 µm. Especie dominante.
Muestras de Herbario: 727, 729, 731, 745, 762.
Hábitat: embalse, canal y charco.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, Myriophyllum aquaticum, natas flotantes de Oedogonium y Spi-rogyra, sustrato arcilloso (perifiton, epifiton, metafiton y plocon).
Sinonimia: Gomphonema brebissonii Kützing, Gomphonema coronatum Ehrenberg, Gomphonema acuminatum var. coronata (Ehrenberg) Rabenhorst.
Registros para la FVTM: DISTRITO FEDERAL: Rio la Magdalena Contreras: Dinamo 4 (Ramírez-Vázquez et al., 2001). ESTADO DE MÉXICO: Lago de Texcoco (Bradbury, 1971); Laguna de Victoria (Mendoza-González, 1985; Ortega, et al., 1994). MORELOS-ESTADO DE MÉXICO: Laguna de Zempoala (Kusel- Fetzmann, 1973; Ortega et al., 1994; García-Rodríguez y Tavera, 1998). Zona Central de México (Metcalfe, 1988).
Información ambiental citada en la literatura: pH: alcalífila, oscila ente 5.4 - 9.0, óptimo 7.8 - 8.5. Estado Trófico: oligotrófica a ligeramente mesotrófica. Halobionte: oligohalobia (indiferente). Saprobionte: oligosapróbica. Corriente: limnófila. Hábitat general: lagos, estanques, charcos, Forma de Vida: bentónica, perifítica, epifítica. Distribución Geográfica: cosmopolita, más abundante en regiones templadas que en regiones tropicales, ampliamente distribuida en aguas de mediana a baja dureza. Europa, Estados Unidos, Este de Afri-ca y Argentina (Lowe, 1974; Patrick y Reimer, 1975; Vinyard, 1979; Germain, 1981; Maidana, 1985; Gasse, 1986; Foged, 1993).
Referencias: Hustedt, 1930, p. 370, fig. 684 como Gomphonema acuminatum var. coronata (Ehrenberg) W. Smith; Patrick y Reimer, 1975, p. 112, Lám.15, fig. 2, 4, 7; Germain, 1981, p. 301, Lám. 110, figs. 1- 4 como Gomphonema acuminatum var. coronata (Ehrenberg) W. Smith; Maidana, 1985, p. 125. Lám. 8, fig. 1; Round et al.,1990, p. 494-495; Novelo, 1998, p. 405, fig. 305.
Gomphonema angustatum (Kützing) Rabenhorst (Figs. 31-33)

Valva lineal o lineal lanceolada, ápices rostrados (Fig. 31). Área axial angosta con un estigma sobre un lado, frente a una estría central (Figs. 31 y 32). Estrías uniseriadas, paralelas en toda la valva, las dos estrías centrales opuestas, más cortas y más distantes de las otras (Fig. 31). Las aberturas de las areolas pueden ser ligeramente alongadas, redondas o en forma de "c" principalmente cerca del rafe (Fig. 33, flecha). La valva presenta un campo de poros pequeños en el polo basal, por los cuales se excreta el mucílago que conforman los pedículos (Fig. 33, flecha). Rafe filiforme central, con los extremos finales ligeramente curvados (Fig. 33). En vista externa los extremos centrales del rafe son rectos (Fig. 32). Medidas de la valvas: eje apical de 18.4-33.6 µm de largo, eje transapical de 5-7 µm de ancho. Densidad de estrías: 12-17 en 10 µm. Especie dominante.
Muestras de Herbario: 726, 731, 740, 755, 763.
Habitat: embalse, canal y charco.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, Myriophyllum aquaticum, natas flotantes de Oedogonium y Spirogyra y en sustrato arcilloso (perifiton, epifiton, metafiton y plocon).
Sinonimia: Sphenella angustata Kützing.
Registros para la FVTM: ESTADO DE MÉXICO: Lago de Texcoco (Bradbury, 1971). MICHOACÁN: Laguna Zacapu (Metcalfe, 1988).
Información ambiental citada en la literatura: pH: alcalífila, oscila entre 6.0-9.0, óptimo 7.5-7.7. Estado Trófico: eutrófica. Halobionte: oligohalobia (indiferente). Saprobionte: oligosapróbica, alfa-mesosapróbica y polisapróbica. Corriente: indiferente. Hábitat general: lagos, estanques, manantiales y arroyos. Forma de Vida: bentónica, perifítica, epifítica. Distribución estacional: otoño e invierno. Temperatura: euri-termal (15 °C o más) a metatermal (5 - 15 °C), oligotermal (0 y 15 °C) a mesotermal (15 °C y 30 °C). Distribución Geográfica: cosmopolita, Europa, Estados Unidos, Este de Africa, Argentina (Lowe, 1974; Patrick y Reimer, 1975; Germain, 1981; Maida-na, 1985; Gasse, 1986; Wasell y Håkansson, 1992; Foged, 1993).
Referencias: Hustedt, 1930, p. 373, fig. 690; Patrick y Reimer, 1975, p. 125, Lám. 17, figs. 17-19; Germain, 1981, p. 306, Lám. 114, figs. 1-21; Maidana, 1985, p. 128, Lám. 8, fig. 17; Round et al., 1990, p. 494-495. Wasell y Håkansson, 1992, p. 174. figs. 28-36, 79-88, como Gomphonema angustatum /parvulum.
Neidium iridis (Ehrenberg) Cleve (Figs. 34, 35)
Valvas elípticas o lanceoladas, con polos redondeados (Fig. 34). Los extremos centrales del rafe terminan en ganchos apuestos (Fig. 35). La superficie de la valva ornamentada con estrías punteadas. Área axial estrecha. Área central ovalada y transversal al eje apical (Fig. 35). Estrías perpendiculares al rafe, muy raramente oblicuas. Con surcos longitudinales cerca del margen de la valva (Fig. 35, flecha). Medidas de la valvas: eje apical de 36-88.7 µm de largo, eje transapical de 9.6-21 µm de ancho. Densidad de estrías: 17-22 estrías en 10 µm. Especie rara.
Muestras de Herbario: 842, 892.
Habitat: embalse y charco.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, Myriophyllum aquaticum, natas flotantes de Oedogonium y Spirogyra (perifiton, epifiton y metafiton).
Sinonimia: Navicula iridis Ehrenberg, Navicula firma Kützing, Pinnularia iridis (Ehrenberg) Rabenhorst.
Registros para la FVTM: DISTRITO FEDERAL: Río La Magdalena Contreras: Dinamo 2 y 4 (Ramírez-Vázquez, 2001). ESTADO DE MÉXICO: Lago de Texcoco (Bradbury, 1971). HIDALGO: Mineral del Monte, Cerca de Real del Monte (Ortega et al., 1994). MORELOS-ESTADO DE MÉXICO: Laguna de Zempoala (Kusel- Fetzmann, 1973; Ortega et al., 1994). Zona Central de México (Metcalfe, 1988).
Información ambiental citada en la literatura: pH: indiferente, oscila entre 4.0 - 9.0, óptimo alrededor de 6. Halobionte: halófobo a Indiferente. Saprobionte: oligosapróbica. Corriente: limnobióntica. Hábitat general: lagos, estanques, pantanos, ciénagas y ocasionalmente ríos. Forma de Vida: bentónica, perifítica epifítica. Distribución Geográfica: cosmopolita, Europa, Estados Unidos, India y Este de Africa (Lowe, 1974; Patrick y Reimer, 1966; Germain, 1981; Sarode y Kamat, 1984; Gasse, 1986; Foged, 1993).
Referencias: Hustedt, 1930, p. 245, fig. 379; Patrick y Reimer, 1966, p. 366, Lám. 34, figs.1; Germain, 1981, p. 148, Lám. 57, figs. 1-4; Sarode y Kamat, 1984, p. 82, Lám. 9. fig. 192; Gasse, 1986, p. 123, Lám. 9, fig. 10, como Neidium iridis var. amphigomphus.
Pinnularia microstauron (Ehrenberg) Cleve (Figs. 36, 37)
Valvas lineales, lanceoladas con polos redondeados a ligeramente rostrados. Área axial de casi 1/4 del ancho de la valva (Fig. 36). Estrías radiales en el centro, cortas en ambos lados y convergentes en los extremos (Fig. 37). El área central de forma rómbica, constituye una fascia transversal (Fig. 37). Rafe filiforme con los extremos finales rectos o ligeramente expandidos, en los extremos distales, el rafe se curva hacia el mismo lado (Fig. 36). Medidas de las valvas: eje apical de 44-63.2 µm de largo, eje transapical de 8-11.2 µm de ancho. Densidad de estrías: 9-11 en 10 µm. Especie dominante.
Muestras de Herbario: 726, 728, 730, 731, 734.
Hábitat: embalse, canal y charco.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, Myriophyllum aquaticum, Sagittaria latifolia, natas flotantes de Oedogonium y Spirogyra y sobre sustrato arcilloso (perifiton, epifiton, metafiton y plocon).
Sinonimia: Stauroptera microstauron Ehrenberg, Navicula bicapitata var. hybrida Grunow.
Registros para la FVTM: DISTRITO FEDERAL: Río La Magdalena Contreras: Dinamo 2 y 4 (Ramírez-Vázquez, et al., 2001). ESTADO DE MÉXICO: Lago de Texcoco (Bradbury,1971). GUANAJUATO: Río Lerma (Ortega et al., 1994). HIDALGO: Atotonilco el Grande; Mpio. Mineral del Monte, cerca de Real del Monte (Ortega et al., 1994).
Información ambiental citada en la literatura: pH: indiferente, alrededor de 7 o ligeramente debajo de 7. Estado Trófico: oligotrófica, mesotrófica. Halobionte: oligohalobia. Corriente: limnobióntica. Hábitat general: lagos, estanques, pantanos, y ocasionalmente ríos. Forma de Vida: bentónica, epifítica, epipélica. Temperatura: euritérmica (15°C o más). Distribución Geográfica: cosmopolita, Europa, Estados Unidos, India, Argentina y Este de Africa (Patrick y Reimer, 1966; Germain, 1981; Sarode y Kamat, 1984; Maidana, 1985; Gasse, 1986; Foged, 1993; Yang y Dickman, 1993).
Referencias: Hustedt, 1930, p. 320, fig. 582; Patrick y Reimer, 1966, p. 597, Lám. 55, fig.12; Germain, 1981, p. 249, Lám. 90, figs. 8 - 11; Sarode y Kamat, 1984, p. 149, Lám. 17. fig. 392; Maidana, 1985, p. 189, Lám. 16, fig. 5; Gasse, 1986, p. 159, Lám 23, figs. 16, 17; Wasell y Håkansson, 1992, p. 168, fig 54, p. 180, fig. 100; Krammer, 1992, p. 98, fig. 32: 9-17; fig. 33:1-19; fig. 34:1-8; fig. 35:9-13; Novelo, 1998, p. 376, fig. 274.
*Planothidium ellipticum (Cleve) Round et Bukhtiyarova (Figs. 38, 39)

Valvas elípticas con ápices redondeados. Estrías muy finas, ligeramente radiales hacia los polos, rafe filiforme con los extremos finales rectos (Fig. 38). Valva sin rafe interrumpida en la parte central y en uno de sus lados, por una depresión en forma de pezuña, "hoof mark" (Fig. 39), según Round y Bukhtiyarova (1996). Medidas de la valva: eje apical de 5 - 10.4 µm de largo, eje transapical de 2.5- 6.2 µm de ancho. Densidad de estrías: 12-15 estrías en 5 µm.
Muestras de herbario: 862, 890.
Hábitat: embalse y canal.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Potamogeton nodosus, Myriophyllum aquaticum, natas flotantes de Oedogonium y Spirogyra y sobre sustrato arcilloso (perifiton, epifiton, metafiton y plocon).
Sinonimia: Achnanthes elliptica (Cleve) Cleve-Euler, Achnanthes elliptica var. elliptica (Cleve) Cleve-Euler, Achnanthes lanceolata var. elliptica Cleve.
*Nuevo registro para la FVTM.
Información ambiental citada en la literatura: pH: alcalífila. Estado Trófico: mesoeutrófica. Halobionte: oligohalobia (Indiferente). Corriente: reófila. Hábitat ríos, lagos, estanques. Forma de Vida: bentónica, epifítica, epipélica.Temperatura: mesotermal. Distribución Geográfica: cosmopolita, Europa, Estados Unidos, India y Este de Africa (Patrick y Reimer, 1966; Germain, 1981; Sarode y Kamat, 1984; Gasse, 1986; Foged, 1993; Håkansson, 1993).
Referencias: Hustedt, 1930, p. 208, fig. 306 c; Patrick y Reimer, 1966, p. 270; Germain, 1981, p. 116, Lám. 44, Figs. 17-18; Sarode y Kamat, 1984, p. 56, Lám. 5, fig. 117; Gasse, 1986, p. 20, como Achnanthes lanceolata var. elliptica; Lange-Bertalot y Krammer 1989, p. 73, Lám. 86, Figs. 33-43, como Achnanthes lanceolada var. elliptica y A. elliptica; Round y Bukhtiyarova 1996, p. 351-352.
Rhopalodia sp. (Figs. 57-59)
Valva isopolar, vista valvar con el margen dorsal convexo (Fig. 57), la zona central ensanchada, margen ventral recto, ápices curvos en vista conectiva (Figs. 58 y 59). Medidas de las valvas: eje apical de 48-53.2 µm de largo, eje transapi-cal de 5 - 13 µm de ancho. Particular del charco permanente.
Muestras de Herbario: 747, 762, 787.
Habitat: charco permanente.
Forma de vida: Crece sobre sustrato arcilloso y natas flotantes de Oedogonium y Spirogyra (Plocon y metafiton).
Stephanodiscus niagarae Ehrenberg (Figs. 40-50)(41, 42, 43, 44, 45, 46, 47, 48, 49)
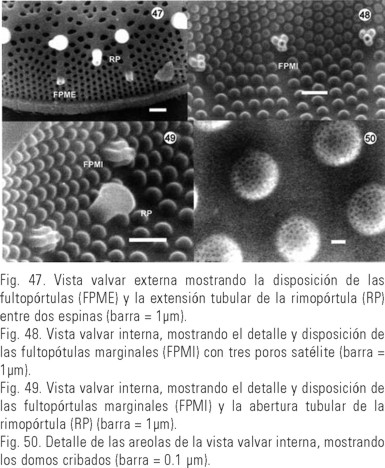

Valvas circulares con numerosos plastos discoidales (Fig. 40). Cara valvar con el centro ligeramente cóncavo o convexo (Fig. 41). Las areolas de la cara valvar están arregladas en fascículos, separados por interfascículos. Los fascículos con dos o tres hileras de areolas hacia el margen, perdiendo su arreglo entre la unión del manto y la valva y una hilera de areolas hacia el centro de la valva. Areolas del área central con arreglo irregular (Figs. 41-43). En la unión del manto y la valva se encuentran las espinas dispuestas cada dos o tres interfascículos (Figs. 41 y 43). Dos a seis fultopór-tulas centrales (Figs. 42 y 44), externamente con un ligero engrosamiento (Fig. 45), internamente están rodeadas por dos o tres poros satélite (Fig. 46), raramente cuatro. Fultopórtulas marginales, una debajo de cada espina (Figs. 41 y 47, FPME), internamente son tubulares, con tres poros satélite (Figs. 48 y 49, FPMI). Entre las espinas se encuentran las rimopórtulas, de tres a cinco por valva, con una extensión tubular larga (Figs. 41 y 47, RP), internamente terminan en una abertura tubular en forma de labio, en dirección oblicua o paralela al margen de la valva (Fig. 49, RP). Las areolas de la cara valvar interna están cubiertas por un domo cribado (Fig. 50). Diámetro de las valvas: 31- 46 µm. Densidad de areolas marginales: 10-18 en 10 µm. Densidad de espinas: 20-44 por valva. Especie dominante.
Muestras de herbario: 735, 757, 762, 769, 776, 786.
Hábitat: embalse, canal y charco.
Forma de vida: Crece sobre natas flotantes de Oedogonium y Spirogyra, sobre sustrato arcilloso y en la columna de agua (metafiton, plocon y plancton).
Registros para la FVTM: ESTADO DE MÉXICO: Lago de Texcoco (Bradbury, 1971); Chalco (Caballero y Ortega, 1998); HIDALGO: Laguna de Tecocomulco (Caballero et al., 1999). MICHOACÁN: Laguna Zacapu (Metcalfe, 1988); Lago Cuitzeo (Israde et al., 2002); Lago Pátzcuaro (Bradbury, 2000). TLAX-CALA: Paleolago Tlaxcala, minas Santa Bárbara y El Lucero (Vilaclara et al., 1997); Paleolago Tlaxcala, Mina Santa Bárbara (Rico et al., 1998).
Información ambiental citada en la literatura: Estado Trófico: oligotrófica, eutrófica. Habitat General: lagos, estanques, ríos. Forma de Vida: bentónica, sedimentos fósiles, plancton. Distribución Geográfica: abundante en los lagos de Norte América (reciente y fósil), sedimentos fósiles de Asia, Africa, Europa y América del Sur (Hakånsson y Locker, 1981; Theriot y Stoermer, 1981, 1984; Theriot et al. 1988; Round et al., 1990; Håkansson, 2002).
Referencias: Håkansson y Locker, 1981, p. 117-149, Lám. 2, figs. 6, 7, Lám. 4, figs. 13-18, Lám. 5, figs. 19-22, Lám. 10, figs. 57-62; Theriot y Stoermer, 1981, p. 64- 72, figs. 1 -21; Theriot y Stoermer, 1984, p. 97-111, fig. 3;Theriot, 1987, p. 359 -373, figs. 3-13;Theriot et al. 1988, p.159-167, figs. 1-10; Round et al., 1990, p. 148, a-d, f-h, i; Krammer y Lange-Bertalot, 1991, p. 67, fig. 68:1-3, 5; fig. 69: 1 a, b; fig. 70: 1; Håkansson, 2002, p. 25, figs. 54-60.
Synedra ulna (Nitzsch) Ehrenberg (Figs. 51-56)
Valvas lineales, con extremos atenuados (Fig. 51). Estrías perpendiculares con respecto al esternón que es estrecho, algunas veces ausentes o poco evidentes en el área central (Fig. 52). Estrías compuestas por hileras de areolas simples, elongadas o redondeadas, ocasionalmente biseria-das, se continúan hasta el manto valvar, el cual forma un ángulo recto con respecto a la cara valvar (Fig. 53). En los ápices de las células se observa un campo de pequeños poros, que con frecuencia estan agrupados debajo de la superficie de la valva (Fig. 54), en la parte superior del campo de poros, se encuentran dos pequeñas proyecciones en forma de cuerno (Fig. 54, flecha). Una rimopórtula esta localizada cerca de cada ápice, y externamente presenta una abertura redonda o ligeramente alargada, como una pequeña depresión (Fig. 55), internamente es una estructura tubular en forma de labio localizada en la penúltima estría (Fig. 56). Medidas de las valvas: eje apical de 115-181.5 µm de largo, eje transapical de 5.7-11.6 µm de ancho. Densidad de estrías: 10-12 estrías en cada 10 µm. Especie dominante.
Muestras de Herbario: 726, 730, 731, 734, 735.
Hábitat: embalse, canal y charco.
Forma de vida: Crece sobre o alrededor de vegetación acuática como Typha latifolia, Potamogeton nodosus, Myriophyllum aquaticum, natas flotantes de Oedogonium y Spirogyra, sobre sustrato arcilloso y en la columna de agua (perifiton, epifiton, metafiton, plocon y plancton).
Sinonimia: Bacillaria ulna Nitzsch, Frustulia splendens Kützing, Synedra lanceolada Kützing, Synedra splendens Küt-zing, Synedra ulna var. lanceolada Grunow, Fragilaria ulna (Ehrenberg) Lange-Bertalot.
Registros para la FVTM: ESTADO DE MÉXICO: Texcoco; Cerro Soltepec (Díaz-Lozano, 1917); Manantial de Almoloya; Manantial de Alta Empresa (Santacruz, 1948); Lerma, ciénagas, lagunas y canales (Rioja y Herrera, 1951); Lago de Texcoco (Bradbury, 1971); Laguna de Victoria (Mendoza-González, 1985; Ortega et al. 1994). GUANAJUATO: Embalse Ignacio Allende (López-López y Serna-Hernández, 1999). HIDALGO: Puente de Dios (Ehrenberg, 1854); MORELOS-ESTADO DE MÉXICO: Laguna de Zempoala (Kusel-Fetzmann, 1973; Ortega et al., 1994; García-Rodríguez y Tavera,1998). MORELOS: Los Manantiales (Valadez et al., 1996). MICHOACÁN: Laguna Zacapu (Metcalfe, 1988); TLAXCALA: Paleolago Tlaxcala, minas El Lucero y Santa Bárbara (Vilaclara et al., 1997; Rico et al., 1998).
Información ambiental citada en la literatura: pH: alcalífila, entre 5.7-9.0. Estado Trófico: eutrófica. Halobionte: indiferente. Saprobionte: oligosapróbica a beta-mesosapróbica. Corriente: oligohalobia (indiferente). Hábitat general: lagos, estanques. Forma de Vida: bentónica, perifítica, epifítica, epipélica, planctónica. Temperatura: euritermal y oligotermal a mesotermal. Distribución estacional: primavera, verano y otoño. Distribución Geográfica: Cosmopolita, Europa, Estados Unidos, India y Este de Africa (Patrick y Reimer, 1966; Lowe, 1974; Germain, 1981; Sarode y Kamat, 1984; Gasse, 1986; Foged, 1993; Håkansson, 1993).
Referencias: Hustedt,1930, p. 151, figs. 158,159; Patrick y Reimer, 1966, p. 148, Lám. 7, figs. 1, 2; Germain, 1981, p. 76, Lám. 24, fig. 1 - 6, fig. 168, fig. 8; Sarode y Kamat, 1984, p. 31, Lám. 2. fig. 37; Poulin et al., 1986 p. 99-112; Williams, 1986, p.133, figs. 1-9; Round et al., 1990, p. 370, figs. a-e; Round, 1991, p. 147-154; Novelo, 1998, p. 298, fig. 196.
Caracterización ambiental
Los valores de las variables físicas y químicas determinadas en la zona litoral superficial del embalse, del canal y del charco se presentan en las tabla 4: en enero y febrero, época de frío y sequía, la temperatura del agua fue de 17.7 y 16 °C respectivamente. Desde abril se fue incrementando (18 y 19.7 °C), hasta alcanzar el valor máximo en el mes de agosto 23.3°C en la época cálida y lluviosa. En octubre, al final de esta misma época, la temperatura presentó un marcado descenso (12.7 °C). En enero y febrero el pH osciló entre 8.6 y 9.0, en abril y junio los valores disminuyeron a 7.6 y 6.3 respectivamente y en los meses de agosto y octubre el pH fue de 8.4 y 7.2. Los valores registrados con el disco de Secchi oscilaron entre 22.0 cm en febrero y 33.3 cm en abril. Los valores de oxígeno disuelto (OD) fueron de 7.8 mg l-1 en febrero, las concentraciones más bajas de OD fueron registradas en abril, junio y agosto de 6.1, 5.6 y 5.7 mg l-1 respectivamente, presentándose un aumento considerable en el mes de octubre de 11.6 mg l-1. Los valores de alcalinidad total oscilaron entre 31.7 mg l-1 de CaCO3 en junio y 55.7 mg l-1 de CaCO3 en abril. Los valores de dureza total oscilaron entre 50.0 mg l-1 de CaCO3 en octubre y 173.3 mg l-1 de CaCO3 en junio. Los valores de conductividad específica (K25) medidos fueron de 140-158.6 µS cm-1. En la tabla 5 se presenta la caracterización ambiental de las diatomeas encontradas en las localidades estudiadas de los Humedales de Jilotepec-Ixtlahuaca.
Discusión
Los resultados indican que el 41.5 % (32) del total de los géneros registrados (77) en la FVTM (Ramírez, 2003) están representados en los tres ambientes estudiados, los cuales corresponden al 13% de los recursos hídricos de la región de los Humedales de Jilotepec Ixtlahuaca, lo que señala la importancia biológica de esta zona (Arriaga et al., 1998). Achnan thes pinnata, Cyclostephanos dubius, Eunotia faba y Planothidium ellipticum son nuevos registros para la Faja Volcánica Transmexicana. Considerando el número de especies (45) los géneros mejor representados en este estudio fueron Gomphonema (4) y Surirella (4). Las especies que dominaron durante todo el período de estudio fueron: Cymbella affinis, Gomphonema angustatum y Synedra ulna (77.8%), Aulacoseira granulata, (66.7%), Gomphonema acuminatum (66.7%) Pinnularia microstauron (66.7%) y Stephanodiscus niagarae (61%). El estudio con MEB nos permitió observar los detalles microestructurales de S. niagarae, especie frecuentemente registrada en los lagos profundos de Norte América y Canadá como especie actual y fósil, y en Centro América sólo como fósil (Theriot y Stoermer,1981, 1984, Theriot et al. 1988). En la región Central de México, esta especie es abundante en material fósil, en lagos profundos (Bradbury,1971, 2000; Caballero y Ortega, 1998; Metcalfe, 1988; Caballero et al., 1999; Israde et al. 2002). En los sitios estudiados de los Humedales de Jilotepec, se encontró una población viva importante de esta especie, en niveles de agua poco profundos (Z< 1 m). La presencia de S. niagarae en esta zona amplía su distribución como especie viviente hacia el sur de Norte América.
La comparación de nuestras observaciones con los datos citados en la bibliografía consultada, ha permitido ampliar la información sobre los requerimientos ecológicos de las especies estudiadas. La mayoría de las especies encontradas en este estudio pueden ser consideradas como cosmopolitas, sin embargo, algunas de ellas como Cymbella tumida es frecuentemente encontrada en regiones tropicales (Silva-Benavides, 1996; Krammer, 2002). García-Rodríguez y Tavera (1998) mencionan a Gomphonema acuminatum para ambientes lénticos y a Synedra ulna para ambientes lénticos y lóticos, distribuidas en zonas templadas y tropicales. En las localidades estudiadas el 89% de las especies fueron limnó-filas, con tendencia a indiferentes (18%) y a reófilas (13%), Epithemia turgida, Eunotia curvata, Cymatopleura solea, Nei-dium iridis y Rhopalodia sp. (11%) se encontraron en el embalse o en el charco permanente por lo que en este estudio se consideraron como limnobiónticas. Hippodonta capitata, Neidium affine y Planothidium ellipticum han sido señaladas en la bibliografía como reófilas y Surirella angusta como reobionte (Lowe, 1974; Gasse, 1986). El 67% (31) de las especies estuvieron ampliamente distribuídas entre los diferentes tipos de sustratos bentónicos (perifiton, epifiton, metafiton o plo-con), el 33% (15) de las especies, también se encontraron en el plancton, por lo que son consideradas como ticoplanctóni-cas. Existen referencias previas sobre la forma de vida de los 45 taxones estudiados, el 88.8% (40) fueron bentónicas y el 11.2% (5) planctónicas (Patrick y Reimer, 1966, Lowe,1974, Gasse, 1986), lo cual coincide con nuestras observaciones, en relación al mayor y menor porcentaje de especies. Aulacoseira granulata, Cymatopleura elliptica y Fragilaria croto-nensis tratadas como planctónicas (Lowe,1974; Germain, 1981), resultaron ticoplanctónicas en este estudio. Las formas de vida de Cymbella affinis, Gomphonema angustatum, Aulacoseira granulata, Pinnularia microstauron y Stephanodiscus niagarae han sido registradas como bentónicas, epilíticas, epifíticas, planctónicas y en microambientes de remanso y deslizamiento (Valadez et al.1996; Cantoral-Uriza et al. 1997; Ibarra-Vázquez y Novelo, 1997; Novelo, 1998; Ramírez-Vázquez et al. 2001; Israde et al. 2002).
Con respecto a las variables físicas y químicas medidas, la temperatura en los tres ambientes estudiados tuvo un comportamiento temporal: octubre a febrero, la época de frío y sequía (15°C), abril y junio época de transición (18.8°C) y en agosto (23°C) la época más cálida y lluviosa. El 54% (25) de los taxa, se encontró durante todo el periodo de estudio, la riqueza de especies fue mayor durante la época de secas. Craticula halophila, Encyonema caespitosum, Frustulia vulgaris y Sellaphora pupula se encontraron en época de lluvias. En general la temperatura ambiental y del agua en los humedales de Jilotepec-Ixtlahuaca fue más baja de lo señalado para ambientes tropicales, en los que se ha registrado más de 20°C (Esteves,1988), presentando afinidad templada. En la bibliografía consultada no hay información sobre los requerimientos de temperatura de todos los taxones tratados. De acuerdo con las temperaturas del agua máximas y mínimas registradas durante el muestreo (11-25 °C) y siguiendo la clasificación propuesta por Patrick (1977), las especies estudiadas son meso-euritermales, el 71.7% (33) se encontró en un intervalo de 11-25°C, es decir presentan una tendencia de aguas frías a templadas, y el 28.3% (13) en un intervalo 15-25°C, caracterizadas como de aguas templadas. Los cuerpos de agua tuvieron un pH tendiente a básico, solo en junio se registraron valores ácidos. Sobre la base de los valores de pH medidos (5.7-9.5), el 65% (30), las especies estudiadas se caracterizaron como alcalífilas, el 37% (11) con tendencia a circumneutrales-alcalífilas, el 33% (10) indiferentes-alcalífilas y el 10% (3) alcalífilas-alcalibiónticas; Eunotia faba se encontró en un pH de 5.7-7.0 (acidófila-circumneutral), Eunotia curvata en un pH de 7 y Surirella tenera en un pH de 5.7-7.5 (circumneutral-indiferente), coincidiendo con la información bibliográfica dada para el 78% (35) de los taxa estudiados (Lowe,1974; Patrick, 1977; Alles et al.1991; Foged, 1993; Håkansson, 1993; Yang y Dickman, 1993). La literatura consultada señala el estado trófico de 24 (53%) de los taxones estudiados, 71% (17) fueron eutróficos: Fragilaria crotonensis y Pla-nothidium ellipticum consideradas como mesoeutróficas, Eunotia curvata, Stauroneis phoenicenteron y Stephanodis-cus niagarae como oligoeutróficas. El 17% (4) de las especies se mencionan como mesotróficas, Gomphonema acuminatum y Pinnularia microstauron fueron oligomesotróficas y Gomphonema clavatun oligotrófica (Lowe,1974; Theriot y Stoermer, 1984; Gasse, 1986; Yang y Dickman, 1993). La presencia de Stephanodiscus niagarae en asociación con Aulacoseira granulata, Fragilaria capucina y F. crotonensis en los sitios de estudio, indica la tendencia a condiciones eutróficas (Theriot et al. 1988; Israde et al. 2002). Considerando los valores de conductividad registrados en los sitios de estudio (140-158.6 µS cm-1) y según la clasificacion de Patrick (1977) todas las especies son de agua dulce y oligohalobias. Los ambientes estudiados se caracterizaron por tener baja visibilidad del disco de Secchi la mayor parte del período de estudio, buena oxigenación, alcalinidad total de baja a moderada y dureza total moderada, por lo que se puede estimar que el agua en las tres zonas de muestreo está poco mineralizada (Vilaclara et al. 1993, Gaytan et al. 1997; Martínez, 1998).
El charco presentó diferencias ambientales con respecto a las otras dos localidades (Tabla 4). La falta de corriente en el charco favoreció el crecimiento de natas flotantes de Spirogyra y Oedogonium, lo que seguramente produjo un microhabitat particular, para Epithemia turgida, Eunotia curvata y Rhopalodia sp. como especies exclusivas a lo largo del estudio, sin embargo, se require un mayor número de estudios ecológicos en la región de los Humedales de Jilotepec, que confirmen lo anterior, además de conocer los intervalos precisos de tolerancia ambiental de las especies estudiadas, que nos permita utilizarlas como indicadores ecológicos.
Agradecimientos
Los autores expresan su agradecimiento a la Dra. Ma. Esther Meave del Castillo, Dr. David Uriel Hernández Becerril, Dr. David A. Siqueiros Beltrones y al Dr. Alfonso Lugo Vázquez, por su revisión, sugerencias y comentarios a este trabajo y a la M. en C. María Patricia Jáquez Ríos por su colaboración en la identificación de las plantas acuáticas que nos permitieron caracterizar el área de estudio.
Referencias
ALLES, E., M. NÖRPEL-SCHEMPP Y H. LANGE-BERTALOT, 1991. Zur Systematik und ökologie charakteristischer Eunotia-Arten (Bacillariophyceae) in elektrolytarmen bachoberläufen. Nova Hedwigia 53(1-2) : 171-213. [ Links ]
A.P.H.A, A.W.W.A. y W.P.F.C. 1985, Standard methods for the examination of water and waste water. 16a. ed. American Public Health As-sociation, Washington, D.C. 1268 p. [ Links ]
ARRIAGA, C.L., V. S. AGUILAR, J. D. ALCOCER, R. R. JIMÉNEZ, E. L. MUÑOZ Y E. D. VÁZQUEZ. (COORDINADORES). 1998. Regiones hidrológicas prioritarias, fichas técnicas y mapa (Escala 1:4 000 000). CONABIO. México. 142 p. [ Links ]
ARRIAGA, C.L., V. S. AGUILAR Y J. D. ALCOCER, 2000. Aguas continentales y diversidad biológica de México. CONABIO. México. 327 p. [ Links ]
BRADBURY, J. P., 1971. Paleolimnology of lake Texcoco, México. Evidence form diatoms. Limnology and Oceanography 16(2) : 180-200. [ Links ]
BRADBURY, J. P., 2000. Limnologic history of Lago Patzcuaro, Michoacan, Mexico for the past 48,000 years: impacts of climate and man. Paleogeography, Paleoclimatology, Paleoecology 163 : 69-95. [ Links ]
BRAUN-BLANQUET, J., 1979. Fitosociología. Bases para el estudio de las comunidades vegetales. H. Blume Ediciones. Madrid. pp. 47-76. [ Links ]
CABALLERO, M. Y B. ORTEGA, 1998. Lake levels since about 40,000 years ago at lake Chalco, near Mexico City. Quaternary Research 50: 69-79. [ Links ]
CABALLERO, M., S. LOZANO, B. ORTEGA, J. URRUTIA Y J. L. MACIAS, 1999. Environmental characteristics of Lake Tecocomulco, northern basin of Mexico, for the last 50,000 years. Journal of Paleolimnology 22: 399-411. [ Links ]
CANTORAL-URIZA, E.A. Y M. ABOAL, 2001. El Margal Pego-Oliva: evolución temporal de la flora de macroalgas. Limnetica 20(1): 159-171. [ Links ]
CANTORAL-URIZA, E. A., J. CARMONA-JIMÉNEZ Y G. MONTEJANO, 1997. Diatoms of calcareous tropical springs in the central region of Mexico. Cryptogamie Algologie 18(1) : 19-46. [ Links ]
CARMONA-JIMÉNEZ, J. Y G. MONTEJANO-ZURITA, 1993. Caracterización ficológica en manantiales de la cuenca baja del sistema hidrológico del Pánuco, México. Boletín de la Sociedad Botánica de México 53 : 21-41. [ Links ]
CETENAL, 1974. Carta topográfica. México E-14-A-18. Escala 1:50 000. S.P.P. [ Links ]
CETENAL, 1975. Carta geológica. México E-14-A-18. Escala 1:50 000. S.P.P. [ Links ]
DÍAZ-LOZANO, E., 1917. Diatomeas fósiles mexicanas. Anales del Instituto Geológico de México 1(1) : 1-27. [ Links ]
EHRENBERG, C., 1854. Microgeologie das erden und felsenchsffende wiekenselbstsandinge lebens auf der erde. Leopold voss. Léipzing. 374 p. [ Links ]
ESTEVES, F. DE A, 1988. Fundamentos de limnología. Interciencia /FINEP, Río de Janeiro. 575 p. [ Links ]
FA, J. Y L. M. MORALES, 1993. Mammals of Mexico: priorizing diversity. In: T.P. Ramamoorthy, R. Bye, A. Lot y J. Fa (Eds.). Biological Diversity of Mexico: origins and distribution. pp. 319-361. Oxford University Press. New York. [ Links ]
FERRUSQUÍA-VILLAFRANCA, I.,1993. Geology of Mexico: A sinopsis. In: T.P. Ramamoorthy, R. Bye, A. Lot y J. Fa (Eds.). Biological Diversity of Mexico: origins and distribution. pp. 3-103. Oxford University Press. New York. [ Links ]
FOGED, N., 1993. Some diatoms from Siberia especially from lake Baikal. Diatom Research 8(2) : 231-279. [ Links ]
GARCÍA-CALDERÓN, G. Y J. RZEDOWSKI, 2001. Flora Fanerogámica del Valle de México. Instituto de Ecología A.C., Centro Regional de Bajío. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. 1406 p. [ Links ]
GARCÍA-RODRÍGUEZ, J. Y R. S. TAVERA, 1998. Fitoplancton del lago Zempoala. Boletín de la Sociedad Botánica de México 63 : 85-100. [ Links ]
GASSE, F., 1986. East African diatoms. Taxonomy, ecological distribution. Bibliotheca Diatomologica. J. Crammer. Berlin - Stuttgart. 201 p. [ Links ]
GAYTAN, M., T. CASTRO, P. BONILLA, A. LUGO Y G. VILACLARA, 1997. Preliminary study of selected drinking water samples in Mexico city. Revista Internacional de Contaminación Ambiental 13(2) : 73-78. [ Links ]
GERMAIN, H., 1981. Flore des diatomées Diatomophycées eaux douces saumâtres du massif armoricain et des contrées voisines d'Europe occidentale. Société Nouvelle Des éditions Boubée Collection. Paris. 444 p. [ Links ]
GOLDSBOROUGH, G. Y G.G.C. ROBINSON, 1996. Pattern in wetlands. Algal ecology. Freshwater benthic ecosystems. pp. 77-117. In: R. J. Stevenson, M.L. Bothwell & R.L. Lowe (Eds). Academic Press. San Die-go, New York, London. [ Links ]
HÅKANSSON, H., 2002. A compilation and evaluation of species in the general Stephanodiscus, Cyclostephanos and Cyclotella with a new genus in the family Stephanodiscaceae. Diatom Research 17(1) : 1-139. [ Links ]
HÅKANSSON, H. Y S. LOCKER, 1981. Stephanodiscus Ehrenberg 1846, a revision of the species described by Ehrenberg. Nova Hedwigia 35: 117-149. [ Links ]
HÅKANSSON, S., 1993. Numerical methods for the inference of pH variations in mesotrophic and eutrophic lakes in southern Sweden - a progress report. Diatom Research 8(2) : 349-370. [ Links ]
HASLE, G. Y G. FRYXELL., 1970. Diatoms: Cleaning and mounting for light and electron microscopy. Transactions of the American Microscopical Society 89(4) : 469-474. [ Links ]
HICKEL, B. Y HÅKANSSON, H., 1987. Dimorphism in Cyclostepanos dubius (Bacillariophyta) and the morphology of initial valves. Diatom Research 2(1) : 35-46. [ Links ]
HUSTEDT, F., 1930. Die süsswasser-flora mitteleuropa. Heft: 10. Bacillariophyta (Diatomeae). Otto Moentz Scientte Publisher. Germany. 465 p. [ Links ]
IBARRA-VÁZQUEZ, C. Y E. NOVELO, 1997. Diatomeas de Texala, Puebla. Boletín de la Sociedad Botánica de México 61 : 49-57. [ Links ]
IBARRA -VÁZQUEZ, C. Y E. NOVELO,1997. Flora diatomológica de una zona de humedales ubicada en la Reserva Ecológica "El Edén" Quintana Roo, México. http://maya.ucr.edu/pril/el_eden/works-hop/diatomesp.html [ Links ]
INDEX NOMINUM ALGARUM., 2000. Página electrónica. http://www.128.32.109.44/e-ina.htlm
INTEGRATED TAXONOMIC INFORMATION SYSTEM., 2002. Página electrónica. http://www.itis.usda.gov
ISRADE, I., V.H. GARDUÑO-MONROY Y R. ORTEGA, 2002. Paleoambiente lacustre del cuaternario tardío en el centro del lago de Cuitzeo. Hidrobiológica 12(1) : 61-78. [ Links ]
KRAMMER, K., 1992. Pinnularia eine monographie der europäischen taxa. Bibliotheca Diatomologica, Bd. 26. J. Cramer. Berlin Stuttgart. 353 p. [ Links ]
KRAMMER, K., 2002. Diatoms of Europa. Diatoms of the european Inland Waters and Comparable Habitats. Cymbella. V.3. In : H. Lange-Bertalot (Ed.). A.R.G. Gantner Verlag K.G. 584 p.
KRAMMER, K. Y H. LANGE-BERTALOT, 1988. 2/2 Bacillariophyceae. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In: Ettl H., Gerloff J., Heynig H. y Mollenhauer D. (Eds.). Süßwasserflora von Mitteleuropa. Stuttgart, Gustav Fischer Verlag. 596 p. [ Links ]
KRAMMER, K. Y H. LANGE-BERTALOT, 1991. 2/3 Bacillariophyceae. 3. Teil Centrales, Fragilariaceae, Eunotiaceae. In: Ettl H, Gerloff J. Heyning H. y Mollenhauer D. (eds), Süßwasserflora von Mitteleuropa. Stutt-gart, Gustav Fischer Verlag. 576 p. [ Links ]
KUSEL-FETZMANN, E., 1973. Beitrage zur Kenntnis der Algenflora der Hochgebirge Zentralamerikas und der nordlichen Anden. Hochgebirgsforschung 3 : 29-68. [ Links ]
LANGE-BERTALOT, H. Y K. KRAMMER, 1989. Achnanthes, eine monographie der gattung mit definition der gattung Cocconeis und nachträgen zu den Naviculaceae. Bibliotheca Diatomologica, 18, J. Cramer, Berlin, Stuttgart. 393 p. [ Links ]
LARA, V. M., J. MORENO Y E. AMARO, 1996. Fitoplancton, conceptos básicos y técnicas de laboratorio. UAM Iztapalapa. México. 227 p. [ Links ]
LÓPEZ-LÓPEZ, E. Y J. SERNA-HERNÁNDEZ, 1999. Variación estacional del zooplancton del embalse Ignacio Allende, Guanajuato, México y su relación con el fitoplancton y factores ambientales. Revista de Biología Tropical 47(4) : 643-657. [ Links ]
LOWE, R. L., 1974. Environmental requirements and pollution tolerance of freshwater diatoms. Cincinnati, Nacional environmental research center office of research and development U.S. Environmental protection agency, 334 p. [ Links ]
LOZANO-GARCÍA, M. S., B. ORTEGA-GUERRERO, M. CABALLERO-MIRANDA Y J. URRUTIA-FUCUGAUCHI, 1993. Late pleistocene and holocene paleoenvironments of Chalco Lake. Quaternary Research 40 : 332-342. [ Links ]
LUGO, H. J., 1984. Geomorfología del Sur de la Cuenca de México. Serie Varia T. 1. Núm. 8 Instituto de Geografía, Universidad Nacional Autónoma de México. México, D.F. 95 p. [ Links ]
LUGO, A., L. BRAVO-INCLÁN, J. ALCOCER, M. L. GAYTÁN, MA. G. OLIVA, MA. DEL R. SÁNCHEZ, M. CHÁVEZ Y G. VILACLARA, 1998. Effect on the planktonic community of the chemical program used to control water hyacinth (Eichhornia crassipes) in Guadalupe Dam, Mexico. Aquatic Ecosystem Health and Management 1 : 333-343. [ Links ]
MAIDANA, N., 1985. Contribución al estudio taxonómico de las diatomeas (Bacillariophyceae) de la provincia de Misión, República de Argentina. Tesis de Doctorado en Ciencias Biológicas. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. Argentina. 276 p. [ Links ]
MANN, D.G. Y A. J. STICKLE, 1991. The genus Craticula. Diatom Re-search 6(1): 79-107. [ Links ]
MARTÍNEZ, C. L. R., 1998. Ecología de los sistemas acuícolas. AGT Editor, México. 227 p. [ Links ]
MENDOZA-GONZÁLEZ, A., 1985. Estudio florístico y ficológico estacional de la laguna de Victoria y Santiago Tilapa, México. Phytologia 58 (7): 479-487. [ Links ]
METCALFE, S., 1988. Modern diatom assemblages in Central Mexico: the role of water chemistry and other environmental factors as indica-te by TWINSPAN and DECORANA. Freshwater Biology 19 : 217-233. [ Links ]
NIXON, C.K., 1993. The genus Quercus in Mexico. pp. 447-458. In: T.P. Ramamoorthy, R. Bye, A. Lot y J. Fa (Eds.). Biological diversity of Mexico: origins and distribution. Oxford University Press. New York. [ Links ]
NOVELO, E., 1998. Floras ficológicas del Valle de Tehuacán, Puebla. Tesis de Doctorado en Ciencias (Biología), Facultad de Ciencias, UNAM, México. 599 p. [ Links ]
NOVELO, E. Y R. S. TAVERA, 1999. Algas y humedales de Quintana Roo. Ciencias 55-56 : 44-45. [ Links ]
OLIVA, MA. G., A. LUGO, J. ALCOCER, L. PERALTA Y MA. DEL R. SÁNCHEZ, 2001. Phytoplankton dynamics in a deep, tropical, hyposaline lake. Hydrobiologia 466 : 299-306. [ Links ]
ORTEGA, M., J. L. GODÍNEZ, G. GARDUÑO Y MA. G. OLIVA, 1994. Ficología de México, algas continentales. AGT Editor. México. 221 p. [ Links ]
OSORIO-TAFALL, B., 1941. Polimorfismo y epifitismo en diatomeas planc-tónicas de Pátzcuaro. Revista de la Sociedad Mexicana de Historia Natural 2 :137-146. [ Links ]
OSORIO-TAFALL, B., 1941A. Materiales para el estudio de microplancton del Lago de Pátzcuaro. Anales de la Escuela Nacional de Ciencias Biológicas 2 : 331-383. [ Links ]
PATRICK, R. Y C. W. REIMER, 1966. The diatoms of the Unites States exclusive of Alaska and Hawaii. Volume I. Monographs of the Academic of Natural Sciences of Philadelphia Number 13. 688 p. [ Links ]
PATRICK, R. Y C. W. REIMER, 1975. The diatoms of the Unites States exclusive of Alaska and Hawaii. Volume II Part 1. Monographs of the Academic of Natural Sciences of Philadelphia Number 13. 213 p. [ Links ]
PATRICK, R., 1977. Ecology of freshwater diatoms and diatom communities. pp. 284-332. In: D. Werner (Ed.). The Biology of Diatoms. Botanical Monographs. Vol. 13. University of California Press. Berkeley and Los Angeles, California. [ Links ]
PIENAAR, C. Y PIETERSE, A. J. H., 1990. Observations on the morphology of Cycostephanos dubius from the Vaal River, South Africa. Diatom Research 5(1) : 201-205. [ Links ]
POULIN, M., L. BÉRARD-THERRIAULT Y A. CARDINAL, 1986. Fragilaria and Synedra (Bacillariophyceae): A morphological and ultrastructural approach. Diatom Research 1(1) : 99-112. [ Links ]
RAMÍREZ, M. J. G., 2003. Catálogo de Bacillariophyta de ambientes epicontinentales de la Faja Volcánica Transmexicana con atlas genérico y estudio diatomológico en el Municipio de Jilotepec, Estado de México. Tesis de Licenciatura en Biología, Facultad de Estudios Superiores Iztacala, UNAM, México. 177 p. [ Links ]
RAMÍREZ-VÁZQUEZ, M., J. BELTRÁN-MAGOS, M. BOJORGE-GARCÍA, J. CARMONA-JMÉNEZ, E. CANTORAL-URIZA Y F. VALADEZ C., 2001. Flora algal del Río la Magdalena, Distrito Federal, México. Boletín de la Sociedad Botánica de México 68 : 45-67. [ Links ]
RAMSAR, 1998. ¿Qué son los Humedales?. http:/www.ramsar.org/about_infopack_types_1e.htm [ Links ]
RAMSAR, 1999. Marco estratégico y lineamientos para el desarrollo futuro de la Lista de Humedales de Importancia Internacional de la Convención sobre Humedales (Ramsar, Irán 1971). http://www.Ramsar.org/key_guide_list_s.htm [ Links ]
REYES-MATAMOROS, J. M. Y D. MARTÍNEZ-MORENO, 2003. Clasificación de los tipos de Vegetación de México. Benemérita Universidad Autónoma de Puebla. Dirección General de Fomento Editorial. México. 11-20 p. [ Links ]
RICO, R., G. VILACLARA, J. MIRANDA Y J. CAÑETAS, 1998. Origin of laminations in Tlaxcala paleolake, Mexico. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie 26(2) : 838-841. [ Links ]
RIOJA, E. Y T. HERRERA, 1951. Ensayo ecológico sobre el limnobio de Lerma y sus alrededores. Anales del Instituto de Biología. Universidad Nacional Autónoma de Mexico 22(2) : 565-591. [ Links ]
ROSAS, I., A. VELASCO, R. BELMONT, A. BAEZ Y A. MARTÍNEZ, 1993. The algal community as indicador of the trophic status of lake Patzcuaro, Mexico. Environmental Pollution 80 : 255-264. [ Links ]
ROSS, R., E. J. COX, N. I. KARAYEVA, D. G. MANN, T. B. B. PADDOCK, R. SIMONSEN Y P. A. SIMS, 1979. An Amended Terminology for the Siliceous Components of the Diatom Cell. Nova Hedwigia, Beihefte 64 : 513-533. [ Links ]
ROUND, F. E., 1991. On stria patterns in Fragilaria and Synedra. Diatom Research 6(1) : 147-154. [ Links ]
ROUND, F. E., R. CRAWFORD Y D. MANN, 1990. The Diatoms. Biology and morfology of the genera. Cambridge University Press. USA. 747 p. [ Links ]
ROUND, F. E. Y L. BUKHTIYAROVA, 1996. Four new genera based on Achnanthes (Achnanthidium) together with a re-definition of Achnanthidium. Diatom Research 11(2) : 345-361. [ Links ]
RZEDOWSKI, J., 1993. Diversity and origins of the phanerogamic flora of Mexico. pp. 129-144. In: T.P. Ramamoorthy, R. Bye, A. Lot y J. Fa (Eds.). Biological Diversity of Mexico: origins and distribution. Oxford University Press. New York. [ Links ]
SANTACRUZ, A., 1948. Estudios del proyecto de abastos de agua de Lerma y recomendaciones para su tratamiento. Órgano Oficial de la Asociación Interamericana de Ingeniería Sanitaria 1:287-299. [ Links ]
SARODE, P. T. Y N. D. KAMAT, 1984. Freshwater diatoms Maharashtra. Institute of Science Aurangabad (Maharashtra, India), 338 p. [ Links ]
SECRETARÍA DE ECOLOGÍA, 2003. Santuarios del agua y forestales. Presa Danxhó, Santa Elena y Xhimojay. Región Hidrológica del Pánuco (RH26). Gobierno del Estado de México. [ Links ]
SILVA-BENAVIDES, A.M., 1996.The epilithic diatom flora of a pristine and a polluted river in Costa Rica, Central America. Diatom Research 11(1) : 105-142. [ Links ]
SLÁDECÉK, V. Y G. VILACLARA, 1993. A waterbloom in Lake Patzcuaro (Michoacan, México). Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie 25 : 431-434. [ Links ]
THERIOT, E., 1987. Principal component analysis and taxonomic interpretation of environmentally related variation in silicification in Stephanodiscus (Bacillariophyceae). British Phycological Journal 22 : 359 - 373. [ Links ]
THERIOT, E. Y E. F. STOERMER, 1981. Some aspects of morphological va-riation in Stephanodiscus niagarae (Bacillariophyceae). Journal of Phycology 17 : 64 -72. [ Links ]
THERIOT, Y E. F. STOERMER, 1984. Principal component analysis of character variation in Stephanodiscus niagarae Ehrenb.: Morphological variation related to lake trophic status. pp. 97-111. In: D. G. Mann (Ed.). Proceedings of the Seventh International Diatom Symposium. Koeltz, Koenigstein. [ Links ]
THERIOT, E., Y. QI, J. YANG Y L. LING, 1988. Taxonomy of the Diatom Ste phanodiscus niagarae from a fossil deposit in Jingyu County, Jilin Province, China. Diatom Research 3(1) : 159 - 167. [ Links ]
VALADEZ, C. F., J. CARMONA-JIMÉNEZ Y E. CANTORAL-URIZA, 1996. Algas de ambientes lóticos en el Estado de Morelos, México. Anales del Instituto de Biología. Universidad Nacional Autónoma de México, Serie Botánica 67(2) : 227-282. [ Links ]
VAN LANDINGHAM, S. L., 1967-1979. Catalogue of the fossil and recent genera and species of diatoms and their synonyms, Vol. 1-8. Cramer, Lehre. [ Links ]
VILACLARA, G., M. CHÁVEZ, A. LUGO, H. GONZÁLEZ Y M. GAYTÁN, 1993. Comparative description of crater lakes basic chemistry in Puebla State, México. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie 25 : 435-440. [ Links ]
VILACLARA, G., R. RICO Y J. MIRANDA, 1997. Effects of perturbations on diatom assenblages in Tlaxcala paleolake, Mexico. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie 26 : 846-851. [ Links ]
VINYARD, W., 1979. Diatoms of Noth America. Eureka Printing Company, Inc. Eureka, California. 179 p. [ Links ]
Wasell, A. y H. Håkansson, 1992. Diatom stratigraphy in a lake on Horseshoe Island, Antartica. A marine-brackish-fresh water transition with comments on the systematics and ecology of the most common diatoms. Diatom Research 7(1) : 157-194. [ Links ]
WILLIAMS, D.M.,1986. Comparative morphology of some species of Synedra Ehrenb. with a new definition of the genus. Diatom Research 1(1) : 131-152. [ Links ]
YANG, J.R. Y M. DICKMAN, 1993. Diatoms as indicators of lake trophic status in central Ontario, Canada. Diatom Research 8(1):179-193. [ Links ]

