










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.14 no.2 Ciudad de México 2004
Artículo
Impacto del cultivo de camarón por succión de larvas de peces y camarón mediante el bombeo de granjas acuícolas en Navachiste, Sinaloa
Impact of the shrimp aquaculture by suction of fish and shrimp larvae by pumping in the aquaculture farms of Navachiste, Sinaloa
Wenceslao Valenzuela Quiñónez,1 José Antonio López Limón2 y Eugenio Alberto Aragón-Noriega3,4
1 Centro Interdisciplinario de Investigación para el Desarrollo Integral Regional-IPN Unidad Sinaloa. Km 1.0 Carretera a las Glorias, Guasave Sinaloa, México. C. P. 81101.
2 Escuela de Biología de la Universidad Autónoma de Sinaloa. Ciudad Universitaria, Culiacán, Sinaloa, México.
3 Centro de Investigaciones Biológicas del Noroeste S. C. Unidad Guaymas. Km 2.35 Camino al Tular, Estero Bacochibampo. Guaymas, Sonora, México 85454
Recibido: 16 de julio de 2003
Aceptado: 31 de marzo de 2004
Resumen
Se evaluó el impacto del bombeo de las granjas de cultivo de camarón de la Bahía de Navachiste, Sinaloa, sobre los estadíos planctónicos de las poblaciones comerciales de peces y camarones que penetran a los estuarios para completar su desarrollo. Se hicieron muestreos mensuales durante 13 meses en los canales de llamada y canales reservorios de dos granjas de camarón seleccionadas al azar, así como en el frente de playa de Navachiste. Se encontró que las larvas de peces se reclutan en los canales de llamada todo el año, pero con dos periodos importantes: primavera y otoño con abundancia mayor a 12 org/m3. La cantidad de postlarvas de camarón capturadas en septiembre fue mayor a 20 org/m3. La densidad de postlarvas en los dos sitios del interior fueron diferentes. Existe una relación directa entre la cantidad de organismos en los canales y la que es capturada en los sistemas de bombeo de las granjas. Se concluye que existe una mortalidad aditiva a la natural por parte de la succión del bombeo acuícola y que la concentración de organismos varía en la misma laguna costera, por lo que el impacto será diferente de acuerdo al sitio en donde se ubique la bomba de succión. Utilizar este tipo de estudios es importante para recomendar sitios de ubicación de las granjas acuícolas en las lagunas costeras.
Palabras clave: zooplancton, bombeo, granjas, impacto, Sinaloa.
Abstract
The impact of shrimp culture over commercial populations of planktonic fish and shrimps that penetrate to the estuaries to complete their development in Bahía Navachiste, Sinaloa was evaluated during pumping. Monthly samplings were made in13 months in the intake water channels and the reservoir channels of two shrimp farms and in the near offshore of Navachiste. It was found that fish larvae are recruited to the intake water channels, throughout the year, but with two important periods: spring and autumn (abundance greater than 12 org/m3). The greatest shrimp postlarva's abundance was in September with 20 org/m3. The abundances in the two interior places were different. There is a direct relationship between the abundance in the intake water channels and the density captured by the pumping systems of the farms. It is concluded that natural mortality is added by shrimp mariculture farms pumping during the early stages of estuarine biota. The impact will be different according to the place where the suction pump is located. This type of studies are important to recommend places of location of shrimp farms in the coastal lagoons.
Key words: zooplankton, pumping, farms, impact, Sinaloa.
Introducción
La pesca de camarón en Sinaloa mantiene el segundo lugar de capturas de altamar y el primero de bahías y esteros, aún cuando en los últimos años la fluctuación de precios ha generado problemática en la comercialización (Aragón-Noriega, 2001). La producción total o producción por área de cultivo de camarón fue positiva hasta los años 1996-1998. Posteriormente se presentaron problemas de enfermedades que limitaron la producción (Lyle-Fritch et al., 2001).
De 1980 al 2000 la acuacultura se consolidó como un nuevo desarrollo de producción de camarón en Sinaloa (Páez-Osuna, 2001); sin embargo, tanto la pesca como la acuacultura se consideran responsables de efectos adversos al ambiente y a las especies (Holloway, 2002). Los efectos desfavorables de la última pueden ser, la destrucción de humedales por la construcción de estanques, eutroficación de los ecosistemas costeros, escape de organismos exóticos, captura de biota estuarina y el uso de químicos por la acuacultura (Hopkins et al., 1995).
El plancton temporal o meroplancton comprende grupos de importancia para la pesca de escama (sardinas, róbalos, meros, cabrillas, pámpanos, pargos y corvinas), crustáceos (camarones y langostas) y moluscos (caracol y almejas) (Wickstead, 1979). Las lagunas costeras son lugares adecuados para el desarrollo de la industria de camarón y por sus características han permitido el establecimiento de las unidades de producción. La acuacultura en su desarrollo genera modificaciones o alteraciones en el área donde se ubica, entre otras, la competencia por área, modificaciones del hábitat, la succión directa y muerte de organismos y la proliferación de enfermedades que impactan en los cultivos (Pillay, 1992).
Las lagunas costeras son áreas de refugio y criaderos naturales para una gran cantidad de especies de peces, crustáceos y moluscos de interés comercial, así como también de numerosas especies de aves que utilizan estos lugares, ya sea estacionalmente o durante todo el año con fines de reproducción o alimentación (Day et al., 1989). Algunos peces pasan la mayor parte de su ciclo vital en estas áreas, como en el caso de varias especies de las familias Gobiidae y Ariidae, mientras otras pasan solamente parte de su ciclo como las familias Carangidae, Centropomidae, Gerreidae o aquellas que utilizan el área únicamente durante la etapa adulta con fines de alimentación como los Ophichthidae (Wickstead, 1979).
Las especies que se encuentran en los canales que abastecen de agua a las granjas acuícolas (canales de llamada) son succionadas por las bombas en relación directa a la concentración temporal de las mismas en los esteros; sin embargo, esto no se ha evaluado con muestreos de campo y se basan en la especulación (Castro-Félix, 2000). Holloway (2002) por ejemplo, en entrevistas a pescadores obtuvo como respuesta que no se sabe con certeza qué está causado la disminución de peces en la costa de Sonora, pero una de las causas puede ser la acuacultura. En Sonora y en otros lugares, los tubos de absorción pueden llevarse las larvas silvestres, despojando a los pescadores de futuras recolecciones. Aragón-Noriega y García-Juárez (2002) hicieron una evaluación de las postlarvas de camarón presentes en la boca de una laguna costera de Sonora y el canal de llamada de una granja camaronera para brindar una respuesta a las dudas planteadas en el párrafo anterior. Dichos autores concluyen que quizás en esa laguna costera y el lugar en donde se encuentra la granja, no exista un impacto por bombeo que incida en las postlarvas de camarón, pero especulan que en una misma laguna costera el impacto puede ser diferente, ya que el plancton presenta una distribución en "manchas". Aragón-Noriega y García-Juárez (2002) también especulan sobre la posibilidad de que la concentración de postlarvas de camarón no sea la misma en las distintas lagunas costeras y por tanto, el impacto de las granjas acuícolas por succión de organismos puede ser diferente y se debe tener cuidado para no extrapolar los impactos de una laguna a otra y entre las diversas zonas de una misma laguna.
En este trabajo se analizó el número de organismos en estadios planctónicos y juveniles tempranos que son capturados a través del bombeo en granjas de cultivo de camarón. Con el propósito de evaluar la capacidad de absorción por bombeo acuícola sobre estos organismos en dos zonas del interior de la laguna costera Bahía Navachiste, Sinaloa, y comparar la abundancia de estos sitios con la zona marina adyacente.
Materiales y métodos
El estudio se realizó en la laguna costera de Navachiste, Sinaloa, localizada entre los 25º 22' y 25º 35' N, y 109º 05' y 108º 45' W. La extensión del sistema abarca 26,700 ha y comprende principalmente la Bahía de Navachiste con 21,400 ha. La profundidad media es de cuatro metros, tiene una barrera arenosa de 23 km de largo, ocho islas y una plataforma de barrera interna. La comunicación con el mar es por dos bocas (Ajoro y Vasequilla) que conectan al Golfo de California (Contreras, 1985). En esta zona se fijaron tres estaciones de muestreo (Fig. 1), una en un frente de playa a un costado de la boca llamada La Bocanita (25º 23' 21'' N y 108º 47' 43'' W); la segunda en una granja acuícola en la zona norte de la bahía (25º 35' 02'' N y 108º 40' 49'' W) y la tercera en una granja de la zona sur (25º 21' 04'' N y 108º 40' 02'' W). En las instalaciones de las granjas acuícolas se realizó el muestreo en los canales de llamada (canal que conduce el agua desde el estero o mar hasta la estación de bombeo de la granja) y en el canal de distribución (canal que inicia después de la estación de bombeo). En ambos casos la muestra se tomó aproximadamente a cinco metros de la bomba.
Para estimar el volumen de agua filtrada se usó una red para plancton de forma cónica con luz de malla de 505 µm y equipada con un medidor mecánico de flujo de agua (General Oceanic, Modelo 2030). Se realizaron arrastres manuales, paralelos a la costa y contra corriente, a intervalos de una hora y con duración de cinco minutos, tanto en playa como en los canales de llamada. Los arrastres en los canales de distribución (después de la estación de bombeo) tuvieron una duración de un minuto y 30 segundos para evitar que la presión de agua dañara a los organismos. La toma de muestra se realizó durante los flujos y reflujos de marea, a una profundidad media de un metro. Se hicieron muestreos mensuales durante 13 meses. Las muestras se fijaron con formol al 4% neutralizado con borato de sodio. La temperatura superficial del agua, el oxígeno disuelto y la salinidad, se registraron en cada arrastre.
Las postlarvas de camarón y los alevines de peces se separaron del resto del zooplancton en el laboratorio. Los grupos fueron identificados con la ayuda de un microscopio óptico, utilizando el objetivo 10X. Las postlarvas de camarón se identificaron con el apoyo de las claves propuestas por Mair (1979), Calderón-Pérez et al. (1989a) y Calderón-Pérez et al. (1989b). La identificación de los alevines y juveniles de peces se llevó a cabo mediante microscopios estereoscópicos, con los que se hizo la observación de las características de valor taxonómico como son: el patrón de pigmentación, número de vértebras, número de radios y espinas en las aletas, longitud del intestino, posición de las aletas, etc. Para la identificación de las familias se utilizaron los trabajos de Dawson (1974) y Gómez-Aguirre et al. (1974). La ordenación sistemática de los especímenes examinados sigue el criterio de Greenwood et al. (1967).
Para el análisis de datos, el volumen de agua filtrado en cada arrastre se estimó a partir de la calibración del medidor de flujo. De las lecturas del medidor del flujo se obtuvo una constante para calcular la distancia (D) en cada arrastre. Por lo que el volumen de agua filtrado es igual a: V=D*A. donde A = área de la boca de la red = 0.2 m2; D = Distancia recorrida (número de revoluciones multiplicada por la constante de calibración). Para analizar la variación de la abundancia de organismos capturados, los datos se expresaron en organismos por metro cúbico de agua filtrada (org/m3), la cual se obtuvo al dividir el número de organismos capturados por muestra entre el volumen de agua filtrada al momento del muestreo. La normalidad de las muestras ya transformadas fue corroborada con la prueba de Kolmogorov-Smirnov, la homogeneidad de varianza se corroboró con la prueba de Levene (Zar, 1984). Las comparaciones se hicieron en dos grandes grupos; postlarvas de camarón y peces.
Los datos no presentaron una distribución normal aún después de trasformados, por lo que se aplicó estadística no paramétrica. La prueba estadística Kruskal-Wallis (Zar, 1984) se usó para evaluar la hipótesis nula de no diferencias entre la abundancia antes y después del sistema de bombeo.
Resultados
En el periodo de enero 2001 a enero del 2002 se realizaron un total de 225 arrastres y se identificaron en total de 34,691 organismos, comprendidos en los grupos de peces, camarones y jaibas.
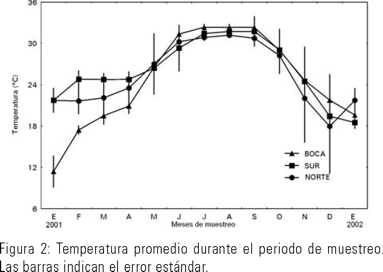
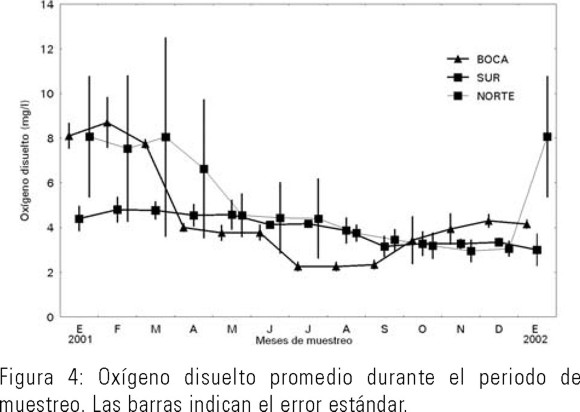
La temperatura promedio del agua en La Bocanita, granja Zona Norte y granja Zona Sur fluctuó entre los 10 y los 30ºC. Los máximos fueron de junio a octubre y los mínimos de noviembre a abril (Fig. 2). La salinidad promedio del agua fluctuó entre las 10 y las 40 ups. Los máximos fueron de junio a octubre y los mínimos de noviembre a abril. La granja norte recibe descargas de agua dulce de drenes agrícolas, lo cual es evidente por presentar menor salinidad y mayor fluctuación de la misma que en los otros puntos de muestreo (Fig. 3). El oxígeno disuelto fluctuó entre los 3 a 10 mg/l. Los máximos fueron de noviembre a marzo y los mínimos de mayo a octubre (Fig. 4).

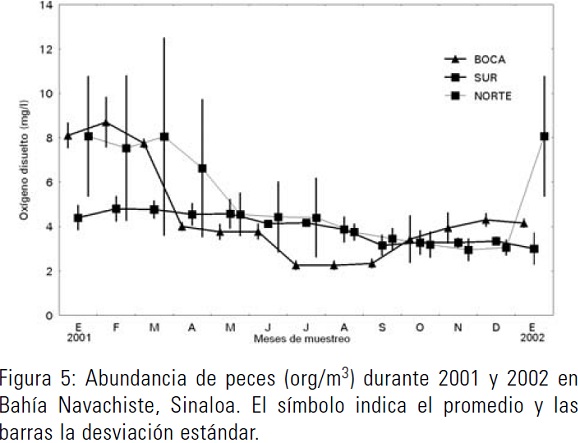
Se identificaron 23 familias de peces en el exterior y el interior de la bahía. Las familias Engraulidae y Gobiidae se capturaron durante todos los meses del año, ocupando el 42 y 25 % de la abundancia total, respectivamente (org/m3). Las familias Gerreidae y Serranidae se presentaron de febrero hasta noviembre, alcanzando 11 y 10 % de la abundancia total, respectivamente. Las familias que siguieron en orden de importancia fueron: Paralichthyidae, Sciaenidae, Gariidae, Bregmacerotidae. Albulidae, Argentinidae, Ammodytidae, Bathylagidae, Bothidae, Carangidae, Cynoglossidae, Gonostomatidae, Myctophidae, Lutjanidae, Mugilidae, Myctophidae, Serranidae, y Tetraodontidae. Éstas fueron encontradas sólo en el periodo de abril a junio en bajas proporciones. Los peces presentaron dos periodos máximos de abundancia, el primero de marzo a mayo y el segundo de agosto a noviembre (Fig. 5). En el frente de playa se encontró la mayor abundancia, siendo el primer máximo en mayo con 8 org/m3 y el segundo con 12 org/m3 en septiembre.
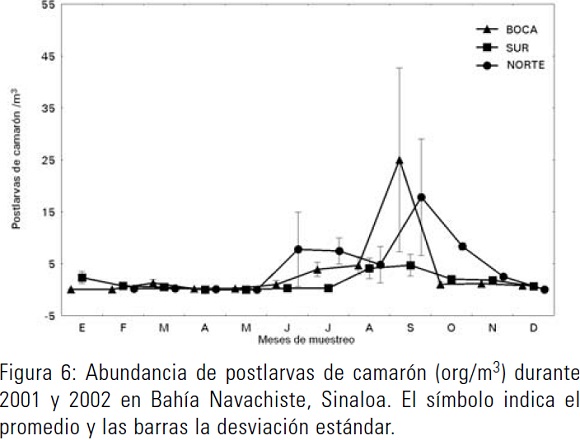
En camarones, se identificaron cuatro especies: camarón café (Farfantepenaeus californiensis) con el 45% del total capturado a lo largo del año, camarón blanco (Litopenaeus vannamei) con el 27%, camarón azul (L. stylirostris) con el 21% y el resto aportado por el camarón rojo (F. brevirostris). Las postlarvas de camarón se presentaron en los meses de junio a noviembre (Fig. 6). En el frente de playa se determinó la mayor abundancia en septiembre con 20 org/m3. En la bahía, la mayor abundancia se presentó en los mismos meses que en el frente de playa.
Para evaluar el impacto de la succión de organismos por el bombeo acuícola, se realizó una comparación entre la abundancia de organismos antes y después del bombeo (abundancia en canal de llamada contra abundancia en canal reservorio). Las figuras 7 y 8 para larvas de peces y postlarvas de camarón, respectivamente, muestran que si la densidad de organismos en el canal de llamada es alta, también lo es en el canal reservorio (después de la succión por los sistemas de bombeo).

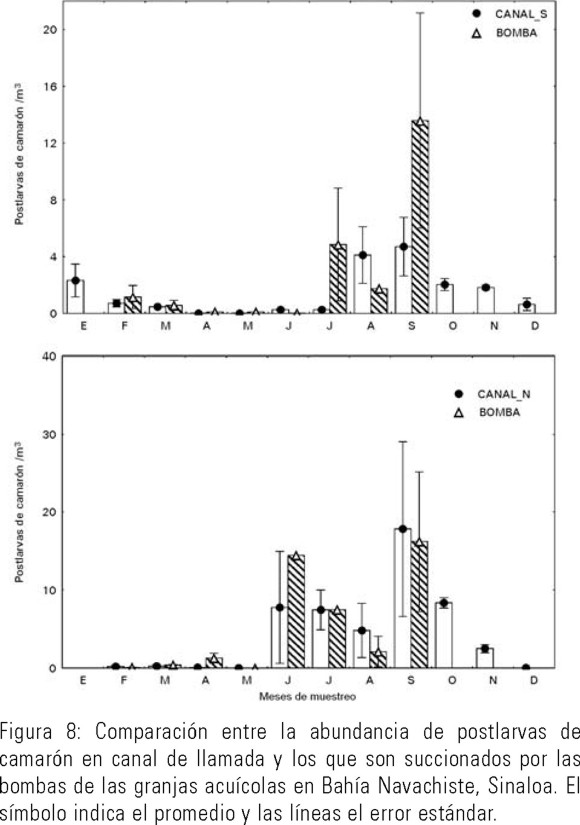
La abundancia de organismos planctónicos varía en el tiempo y entre las zonas de muestreo, pero siempre la cantidad de organismos succionados está en relación directa con la densidad en los canales de llamada (Figs. 7 y 8). La prueba estadística no paramétrica Kruskal-Wallis para las abundancias de organismos entre los canales antes y después del sistema de bombeo mostró que la abundancia media que se observó en las área de crianza y la que ingresa a la granja de camarón no es significativamente diferente (H = 0.38 y P = 0.85). Por lo que se considera que no hay razón para suponer que se evita la succión de organismos por alguna vía de respuesta del organismo o actividad del acuicultor por impedirlo.
Discusión
Las granjas camaronícolas son dependientes del uso de agua de los estuarios para el llenado de estanques y posteriormente para el recambio de la misma durante las actividades diarias. La remoción de la biota de los estuarios en el proceso de transferir agua hacia los estanques causa preocupación por el posible impacto al ambiente (Hopkins et al., 1995). Es concebible que organismos estuarinos puedan ser bombeados a los estanques y posteriormente descargados de nuevo a los esteros con el recambio de agua, pero es probable que esto no ocurra y que la mayoría de los organismos mueran. Este caso ha sido muy discutido sin presentar datos que lo avalen (Castro-Félix, 2000) y sin profundizar en el tema (Hendrickx et al., 1995) o sobre grupos muy específicos como las postlarvas de camarón (Aragón-Noriega y García-Juárez, 2002). La relevancia de este estudio es que aporta datos comparativos entre las abundancias de los primeros estadios de peces y camarones en puntos situados antes y después del bombeo de las granjas acuícolas.
Estudios poblacionales sobre especies de camarón con hábitos de vida similares a los del Golfo de California indican que las pesquerías tradicionales de este recurso, son más afectadas por la falta de espacio y condiciones ambientales durante las fases de juvenil y subadultos, que por la reproducción y el reclutamiento (Lam et al., 1989). A esto hay que agregar el hecho de que la depredación en juveniles y adultos es intensa y que la extracción de camarones influye en las relaciones depredador-presa.
El impacto de la extracción de postlarvas en las poblaciones de camarón varía en función de la edad y es menor cuando la extracción se realiza en las primeras fases y se incrementa exponencialmente en los juveniles (Gracia, 2001). Este mismo autor concluye que el efecto de extraer postlarvas es menor cuando se captura a los organismos en las zonas cercanas a las bocas de las lagunas costeras y en el momento cuando inmigran a las áreas de crianza. Una vez que se han establecido en las áreas de crianza, el efecto se incrementa exponencialmente. También señala que los modelos de simulación para Litopenaeus setiferus con relación a una extracción de organismos de 29 mm de longitud total para sembrar 4,000 ha, podría reducir hasta un 40% la producción pesquera. Hay que recordar que las postlarvas de camarón capturadas durante este estudio son organismos ya establecidos en las áreas de crianza. De hecho se han capturado en los esteros de la parte más interna de la laguna costera de Navachiste, por lo que de acuerdo con Gracia (2001) su impacto es muy alto sobre la población silvestre explotada.
Aragón-Noriega y García-Juárez (2002) mencionan que para probar la hipótesis del posible impacto de las granjas sobre la producción natural de camarón, se debe incluir un número mayor de granjas distribuidas en varios sistemas. Ellos argumentan con sus resultados que hay sitios donde las granjas acuícolas no impactan negativamente. Esto lo atribuyen al patrón de dispersión de las postlarvas silvestres además de que en el sistema donde ellos realizaron su trabajo (Guásimas, Sonora) no se concentran las postlarvas en los canales de llamada de las granjas; sin embargo, en Navachiste, Sinaloa (lugar donde se realizó el presente estudio) se encontró que la densidad de postlarvas de camarón es muy superior (20 PL/m3) a la que ellos encontraron en Guásimas (5 PL/m3). Aragón-Noriega y García-Juárez (2002) mencionan que una posible explicación para la baja abundancia que encontraron, fue la alta salinidad. Los resultados del presente estudio contradicen su idea, ya que en Navachiste se encontraron dos sitios con diferente salinidad y no hubo diferencia en la densidad de postlarvas de camarón. Incluso la salinidad evaluada a lo largo del estudio en la granja sur de Navachiste es similar a la de Guásimas. El patrón de temperatura en Guásimas es el mismo al de Navachiste, por lo que tampoco es ésta una variable ambiental que explique la diferencia en abundancia.
Lo importante de comparar este estudio con el de Aragón-Noriega y García-Juárez (2002) radica en valorar la relevancia de conocer la abundancia de postlarvas de camarón de los sistemas donde se practica la acuacultura de camarón, para no extrapolar los efectos de un sistema a otro, ya sea negativo como en este caso o sin efecto aparente como en Guásimas. Se resalta que es necesario comparar los posibles impactos de un mismo sistema en diferentes sitios, ya que éste puede presentar diferencias en la abundancia de postlarvas de camarón como el caso de este estudio.
El ingreso de huevos, larvas y juveniles de peces, que luego serán depredadores potenciales del camarón, es una preocupación constante de los acuicultores (Hendrickx et al., 1995). Los filtros necesarios para retener los huevos de peces no son selectivos como un control contra los depredadores, ya que otras especies son destruidas por la presión del agua ejercida contra la mallas de los filtros. Los acuicultores filtran el agua que ingresa a los estanques con mallas de 200 a 300 micras para quitar los huevos de peces y crustáceos que pueden además ser portadores de virus u otros patógenos. Los organismos capturados en las granjas donde se hizo el presente estudio comprenden peces de la familia Albulidae con valor en la pesca deportiva con caña, y de la familia Clupeidae quienes constituyen una de las pesquerías más importante del área, y Engraulidae que son usados para la fabricación de harina y aceite. Hendrickx et al. (1995) mencionan que por el tamaño de la malla protectora en las compuertas de entrada y tomando en cuenta el tamaño de las larvas de peces es posible que éstas puedan entrar al estanque de cultivo. Durante este estudio se observó que en las mallas de filtrado prevalecieron larvas y juveniles de peces lo que indica que muchos organismos mueren en estas estructuras y jamás regresan al estero como se pensaba.
Los máximos de abundancia de peces encontrados en Navachiste, Sinaloa, coinciden con los periodos de llenado de las granjas de camarón, el primer ciclo inicia en febrero y marzo y el segundo de junio a agosto. La cantidad de organismos zooplanctónicos succionados por las bombas de la zona norte y sur está directamente relacionada con la densidad en las zonas adyacentes. Lo que hace suponer el efecto aditivo de la captura y muerte de postlarvas en el bombeo de las granjas de camarón.
En el estado de Texas, E. U. se ha intentado controlar la destrucción de la fauna estuarina con medidas de prevención impuestas a los acuacultores. Los permisos para operar granjas acuícolas son concedidos después que se ha instalado un sistema de barreras de diversos tamaños de luz de malla antes del sistema de bombeo. Otro tipo de restricción consiste en mantener la velocidad de absorción en las tomas de agua por debajo de 10 cm/s. En este trabajo durante las observaciones de campo el material bombeado que es retenido en los sacos en luz de malla de 200 a 300 micras consistió principalmente de detritus orgánico de origen vegetal. Los organismos dominantes en las redes de filtrado fueron larvas de cangrejos destrozadas y peces grandes. Lo que imposibilita el hecho de que una limpieza constante de los sacos pueda representar una solución al problema, ya que habría que añadir que los organismos de tamaño pequeño y cuerpo más frágil son presionados contra la malla de filtrado antes de entrar a los estanques.
Finalmente, el aporte de este estudio ya no deja lugar a la especulación como lo menciona Holloway (2002). Este trabajo presenta de forma cuantitativa el tipo de organismos que están siendo succionados por el bombeo acuícola en la laguna costera de Navachiste, Sinaloa. Se hace mención sobre los peces y camarones en forma separada para hacer más efectiva la comparación que Holloway (2002) menciona en su estudio.
Se concluye que la concentración de organismos varia en la misma laguna costera, por lo que el impacto será diferente de acuerdo al sitio en donde se ubique la bomba de succión. El conocimiento de la abundancia del plancton en el espacio y el tiempo debe ser utilizado por los acuacultores para programar sus actividades buscando reducir la extracción de postlarvas de camarón. Se cree que una opción viable para evitar la captura de alevines de peces podría ser la disminución de la potencia de los motores, ya que el periodo de presencia de larvas de peces en esta laguna costera es muy prolongado.
Agradecimientos
A la Coordinación General de Postgrado e Investigación del Instituto Politécnico Nacional con registro No 200343 y al Consejo Estatal de Ciencia y Tecnología del Estado de Sinaloa por el apoyo económico. Alan Alfredo Zavala Norzagaray y Pindaro Álvarez Ruiz por su colaboración durante el desarrollo del proyecto. Edgar Alcántara Razo revisó la versión final del manuscrito. David Urías Laborín editó las figuras.
Referencias
ARAGÓN-NORIEGA, E. A., 2001. Estado actual de producción de camarón en el Pacífico Mexicano: Pesca y Cultivo. In Castro-Castro V., M. Y. Sokolov Y A. de Jesús-Navarrete (Eds.) Memorias del Primer Foro de Pesca y Acuacultura de la Costa de Chiapas. El Colegio de la Frontera Sur, pp. 53-59. [ Links ]
ARAGÓN-NORIEGA, E. A. & A. R. GARCÍA-JUÁREZ, 2002. Incidencia de postlarvas de camarón (Crustacea: Penaeidae) en el canal de llamada de una granja camaronera del Estado de Sonora. In: M. E. Hendrickx (Ed.) Contribuciones a los crustáceos del Pacifico Este Vol. 1. Instituto de Ciencias del Mar y Limnología, UNAM, pp. 145-154. [ Links ]
CALDERÓN-PÉREZ, J. A., E. MACÍAS-REGALADO & S. RENDÓN-RODRÍGUEZ, 1989a. Clave de identificación para los estadios de postlarva y primeros juveniles de camarón del género Penaeus (CRUSTACEA: DECAPODA.) del Golfo de California. México. Ciencias Marinas 15 (3): 57-70. [ Links ]
CALDERÓN-PÉREZ, J. A., E. MACIAS-REGALADO, F. A. ABREU-GROBOIS & S. RENDÓN-RODRÍGUEZ, 1989b. Antennular flagella: A useful character for distinguishing subgenera among postlarval shrimp of the genus Penaeus (Decapoda) from the Gulf of California. Journal of Crustacean Biology 9 (3):482-491 P. [ Links ]
CASTRO FÉLIX A., 2000. Dos actividades económicas importantes y un problema ambiental y social sin resolver. Panorama Acuícola 5 (4): 50-52. [ Links ]
CONTRERAS, F., 1985. Lagunas costeras. Centro de Ecodesarrollo y Secretaría de Pesca, México. 235 p. [ Links ]
DAWSON, E. C., 1974. A review of Microdesmidae. COPEIA 2: 409-448 [ Links ]
DAY, JR. J. W., C. A. S. HALL, W. M. KEMP & A. YAÑEZ-ARANCIBIA, 1989. Estuarine Ecology. John Wiley & Sons. Nueva York. 558 p. [ Links ]
GÓMEZ-AGUIRRE, S., S. LICEA-DURAN & C. FLORES-COTO, 1974. Ciclo anual del plancton en el sistema Huizache-Caimanero, México. Anales del Centro de Ciencias del Mar y Limnología de la UNAM 1:1-304 [ Links ]
GRACIA, A., 2001. Interacción entre la utilización de postlarvas silvestres para cultivo y las pesquerías de camarón. In: Páez-Osuna (Ed.) Camaronicultura y medio ambiente. Instituto de Ciencia del Mar y Limnología. Programa Universitario de Alimentos y El Colegio de Sinaloa. pp 397-412. [ Links ]
GREENWOOD, P. H., D. E. ROSEN, S. H. WEITZMAN & G. S. MYERS, 1967. Named main division of teleostean fishes. Proceedings of Biological Society of Washington 80: 227-248. [ Links ]
HENDRICKX, M. E., J. SALGADO-BARRAGÁN & M. A. MEDA-MARTÍNEZ, 1995. Abundance and diversity of macrofauna (fish and decapod crustaceans) in Penaeus vannamei culture ponds in Western Mexico. Aquaculture 143: 61-73. [ Links ]
HOLLOWAY, M. 2002. Revolución Azul. Discover 6 (10): 49-55. [ Links ]
HOPKINS, J. S., M. R. DEVOE & A. F. HOLLAND, 1995. Environmental impacts of shrimp farming with special reference to the situation in the continental United States. Estuaries 18 (1A): 25-42 [ Links ]
LAM, C. F., J. D. WHITAKER & F. L. LEE, 1989. Model for white shrimp landings for the central coast of South Carolina. North American Journal of Fisheries Management 9: 12-21.G [ Links ]
LYLE-FRITCH, L., E. ROMERO-BELTRÁN & J. A. BECT VALDÉS, 2001. Desarrollo y características de las granjas camaronícolas en Sinaloa. In Páez-Osuna (Ed). Camaronicultura y medio ambiente. Instituto de Ciencia del Mar y Limnología. Programa Universitario de Alimentos y El Colegio de Sinaloa, pp 351-372. [ Links ]
MAIR, J. M., 1979. The identification of postlarvae of four species of Penaeus (CRUSTACEA: DECAPODA) from the Pacific Coast of México. Journal of Zoology 188: 347-351. [ Links ]
PÁEZ-OSUNA, F., 2001. Impacto ambiental de la camaronicultura: Causas, efectos y alternativas de mitigación. In: Páez-Osuna (Ed.) Camaronicultura y medio ambiente. Instituto de Ciencia del Mar y Limnología. Programa Universitario de Alimentos y El Colegio de Sinaloa, pp 413-426. [ Links ]
PILLAY, T., V. R., 1992. Aquaculture and the Environment. John Wiley & Sons, Nueva York. 189 p. [ Links ]
WICKSTEAD, J. H., 1979. Zooplancton marino. Ediciones Omega. Barcelona. 70 p. [ Links ]
ZAR, J. H., 1984. Biostatistical analysis. Prentice-Hall, Englewood. 718 p. [ Links ]

