










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.13 no.3 Ciudad de México Out. 2003
Artículo
Distribución y abundancia de los quetognatos de la plataforma Tamaulipeca y océano adyacente en abril de 1987
Distribution and abundance of chaetognaths on the continental shelf of Tamaulipas and adyacent oceanic zone during april 1987
Silvia Mille-Pagaza y Jorge Carrillo-Laguna
Escuela Nacional de Ciencias Biológicas-I.P.N. Prol. Carpio y Plan de Ayala S/N. Casco de Santo Tomás. C.P. 11340. México, D.F. jcarrill14@hotmail.com
Recibido: 20 de julio de 2002.
Aceptado: 4 de septiembre de 2003.
Resumen
Se analiza la distribución y composición de las especies de quetognatos de 16 estaciones localizadas frente al estado de Tamaulipas en la parte oeste del Golfo de México. Las muestras fueron colectadas durante el crucero "Justo Sierra" realizado en abril de 1987. Se encontraron en total 13 especies, con la más baja riqueza hacia el norte del área de estudio frente a laguna Madre. Flaccisagitta enflata y Sagitta tenuis fueron las de mayor densidad, con valores entre 2000 y 6000 individuos100m-3 en la porción norte del área. F. enflata junto con Serratosagitta serratodentata estuvieron presentes en todas las estaciones, ésta última con las densidades más altas en la zona sur. Se detectaron tres grupos de especies mediante el Valor de Importancia, el primero incluyó a F. enflata, S. tenuis, S. serratodentata. Con la similitud de Morisita se definieron dos grupos de estaciones; el primero comprendió siete sitios ubicados al norte del área de estudio y la estación más sureña de todas, donde el promedio de la diversidad de Shannon fue bajo (1.3 bits individuo-1) y la dominancia de Simpson tuvo un valor intermedio (0.51). El segundo grupo lo formaron las siete estaciones restantes situadas al sur del área, con una diversidad promedio de 2.2 bits individuo-1 y la media de la dominancia de 0.27. El análisis de componentes principales permitió separar un conjunto de estaciones neríticas y otro de estaciones oceánicas, coincidiendo básicamente con los resultados del índice de similitud. La salinidad y la temperatura fueron los factores abióticos que probablemente influyeron en la distribución y abundancia de las especies de quetognatos en el área.
Palabras clave: Quetognatos, densidad poblacional, similitud, diversidad, Golfo de México.
Abstract
An analysis of Chaetognatha species distribution and abundance is presented for the northwestern region of the Gulf of Mexico. Zooplankton samples were collected during April 1987 at 16 collection sites located in front of the State of Tamaulipas. Thirteen species were recorded. Specific richness was lower at northwestern sites in front of Laguna Madre. Species with the highest density (2000 to 6000 orgs.100m-3) were Flaccisagitta enflata y Sagitta tenuis which were found towards the north of the area. The most frequent species collected throughout the sites were F. enflata and Serratosagitta serratodentata, the last one showed its highest densities in the southern region. Three groups of species were detected by using the Importance Value (IV): the first set included F. enflata, S. tenuis and S. serratodentata. Also two groups were defined when density data were analyzed by Morisita's similarity index. The first included 7 northerly sites and the second, the most southern site. The last one shows a low Shannon diversity value (1.3 bits individual-1) and an intermediate Simpson's dominance value (0.51). The second group included the rest of the 8 southern stations, with an average diversity of 2.2 bits individual-1 and a mean dominance of 0.27. A Principal Component analyses ran on abiotic variables grouped the stations in two sets: oceanic and neritic, this in turn agreed with the results of the similarity index. Chaetognaths species distribution and abundance were probably influenced by salinity and temperature.
Key words: Chaetognaths, specific density, similarity, diversity, Gulf of Mexico.
Introducción
De acuerdo con varios autores el grupo Chaetognatha, conformado principalmente por especies planctónicas, constituye un taxa de gran interés por su amplia distribución a nivel mundial; entre las especies reportadas sobresalen algunas cosmopolitas que se han registrado tanto en los océanos del hemisferio norte como en los del sur del planeta (Alvariño, 1969; Ulloa & Palma, 2000). Además se considera uno de los grupos holoplanctónicos más abundantes, tanto en aguas oceánicas como neríticas (Álvarez-Cadena et al., 1996; McLelland, 1989; Stuart & Verheye, 1991). Su abundancia y alta capacidad depredadora, principalmente de copépodos (Álvarez-Cadena et al., 1996; Oresland, 2000), lo convierte en un conjunto importante dentro de las cadenas tróficas del sistema marino (Stuart & Verheye, 1991). Todo lo anterior, sumado al hecho de considerarlos indicadores hidrológicos por su asociación con determinadas condiciones físicoquímicas de las masas de agua (McLelland, 1989), hace de ellos un grupo de gran interés para su estudio.
Sin embargo, los estudios de quetognatos en el Golfo de México se han concentrado fundamentalmente en la zona norte y noreste, en las corrientes cercanas a la Península de Florida (Owre, 1960; Pierce & Wass, 1962); cerca de la costa de Texas (Pierce, 1962); en las proximidades de la desembocadura del río Mississippi (McLelland, 1989); en la zona oceánica del noreste del Golfo (McLelland & Perry, 1989) y en los alrededores de Cuba (Suárez-Caabro, 1955). Asimismo, hay algunos estudios en el mar Caribe (Michel & Foyo, 1976; Gasca et al., 1996). En la Zona Económica Exclusiva Mexicana del Golfo se han realizado varios estudios muy puntuales en regiones cercanas a la costa (Vega-Rodríguez, 1965; Rivero-Beltrán, 1975). Recientes trabajos de este grupo en el Golfo de México ampliaron estos estudios con las publicaciones sobre las especies encontradas en dos cruceros realizados en la Plataforma de Campeche (Mille-Pagaza et al., 1997; Mille-Pagaza & Carrillo-Laguna, 1999) y en la zona suroeste del Golfo abarcando tanto la región cercana a la Plataforma como la oceánica (Mille-Pagaza & Carrillo-Laguna, 2001). En el presente trabajo se analizan las especies encontradas en el muestreo de abril de 1987 en la zona oeste del golfo sobre la plataforma Tamaulipeca y parte de la zona oceánica y se busca establecer con precisión la relación entre las especies de quetognatos y ciertas condiciones de las masas de agua donde se presentan.
Materiales y métodos
Se analizaron 16 muestras recolectadas mediante arrastres oblicuos con red bongo de 333 µm de malla, a bordo del buque oceanográfico "Justo Sierra" en abril de 1987 (JS 8701), por personal del Instituto Nacional de la Pesca. Las muestras proceden de la región oeste del Golfo abarcando la parte norte de la Plataforma tamaulipeca y una porción de la zona oceánica (Fig. 1). La profundidad de las estaciones en general, fue menor a los 100m y solamente siete se ubicaron fuera de la plataforma, entre 400 y 2000 m de profundidad. En cada estación se registraron en superficie la temperatura (ºC), la salinidad (ups) y el oxígeno (ml l-1).

La identificación de las especies de quetognatos se hizo con base en las descripciones de McLelland (1989). Por estación se calculó la densidad total, la riqueza específica de Menhinick (Ludwig & Reynolds, 1988), la diversidad de Shannon y la dominancia de Simpson (Krebs, 1994; Smith & Smith, 2001). Para cada especie se estimó la densidad en organismos por 100 m3 y el Valor de Importancia (VI) (Krebs, 1994), el cual se obtuvo sumando la valores relativos de frecuencia y abundancia. Con el criterio de clasificación de Dajoz (1996) se establecieron las especies constantes, ocasionales y accidentales.
Se calculó la similitud entre estaciones con el Índice de Morisita y el dendrograma se elaboró con el ligamiento de promedios no ponderados (UPGMA) (Ludwig & Reynolds, 1988, Krebs, 1999). Con el fin de caracterizar el ambiente abiótico se empleó el análisis de componentes principales (Ludwig & Reynolds, 1988, Krebs, 1999) y para establecer las variables abióticas que posiblemente influyeron en la composición y abundancia de las especies de quetognatos se utilizó el estadístico T2 de Hotelling (Morrison, 1990).
Resultados y discusión
En este crucero se recolectaron trece especies de quetognatos; entre 5 y 7 especies se obtuvieron en las estaciones más norteñas y entre 7 y 8 para las sureñas; sin embargo, en tres estaciones aisladas hubo nueve especies, una de ellas situada al norte del área y dos al sur. Los valores del índice de Menhinick variaron entre 0.06 y 0.16 en la porción norteña y entre 0.19 y 0.27 para la parte sureña por lo que la mayor riqueza específica y el mayor número de especies la mostraron las estaciones sureñas.
Las especies más frecuentes y con mayor densidad promedio en el área (>1077 org.100m-3) fueron Flaccisagitta enflata (Grassi, 1881) y Serratosagitta serratodentata (Krohn, 1853) presentes en todas las estaciones (16); Krohnitta subtilis (Grassi, 1881), aunque se obtuvo en 15 estaciones, presentó poca abundancia (6.25 org.100m-3) mientras que, Mesosagitta decipiens (Fowler, 1905), M. sibogae (Fowler, 1906) y Ferosagitta hispida (Conant, 1895) se encontraron únicamente en una estación y Flaccisagitta lyra (Krohn, 1853) en dos, todas ellas al sur, con abundancias promedio menores a 2 org.100m-3.
Se registraron densidades totales por estación (Fig. 2) entre 3 500 y 14 000 org.100m-3 en la zona norte del área, que de acuerdo con varios autores es una zona con temperatura relativamente baja y mayor concentración de oxígeno debido al afloramiento del agua profunda, lo que se traduce en un florecimiento importante de fitoplancton (Moulin-Ruíz, 1980; Vázquez de la Cerda, 1975) y en consecuencia mayor cantidad de copépodos fitófagos y por tanto, de depredadores como los quetognatos, semejante a lo señalado por Alvariño (1968) para la zona de influencia del Amazonas.
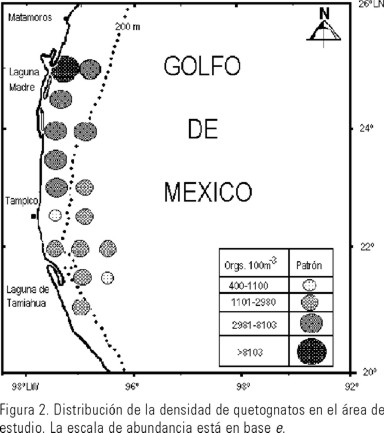
Los valores de temperatura registrados en este mes para las estaciones norteñas fueron más bajos (23.5ºC ±0.5, α = 0.05) que en las sureñas, donde la temperatura fue mayor (24.3ºC ±0.3, α = 0.05), con excepción de la más sureña que mostró 22.1ºC. La masa de agua con menor temperatura estuvo integrada por 7 estaciones de plataforma y la 5 del talud, en esta masa, F. enflata tuvo las mayores densidades (de 1008 a 6810 org.100m-3) ya que de acuerdo con lo señalado por McLelland (1989), esta especie es considerada poco tolerante a las altas temperaturas.
Los valores de densidad total por estación más altos, correspondieron a las estaciones uno y cuatro ubicadas frente a la desembocadura de las bocas de la Laguna Madre y fueron aportados fundamentalmente por F. enflata; sin embargo, Sagitta tenuis Conant 1896, también contribuyó a esta alta densidad con el valor máximo (6003 org.100m-3) en la estación cuatro, confirmando su preferencia por las masas de agua neríticas. Las densidades totales por sitio presentaron un patrón similar al encontrado para abril-mayo de 1986 por Mille-Pagaza y Carrillo-Laguna (2001) en las tres estaciones situadas al noreste del área estudiada. En ambos cruceros el número de especies fue el más bajo y la densidad total más alta la aportaron Flaccisagitta enflata y Sagitta tenuis. Estas dos especies mostraron un comportamiento muy similar para la zona de estudio en ambos años, con un descenso muy marcado en sus densidades en las estaciones oceánicas.
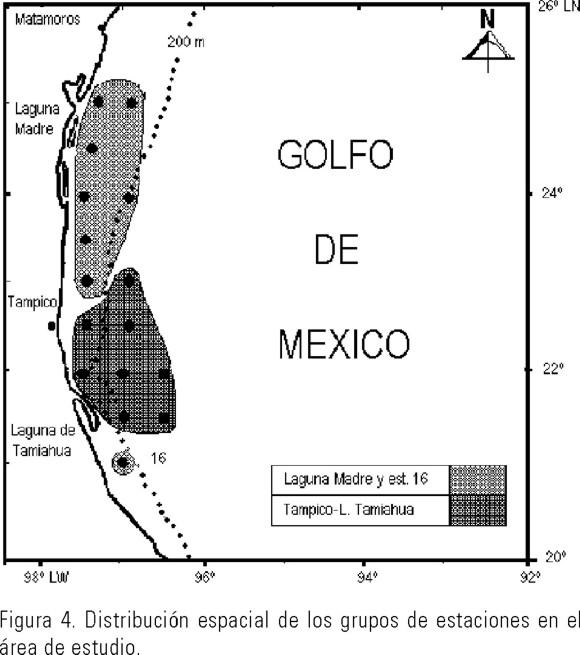
Por el contrario las densidades totales más bajas de ambas especies se presentaron en las estaciones oceánicas o en estaciones de plataforma influenciadas por la masa de agua oceánica, particularmente en el sitio nueve, ubicado sobre la plataforma frente a Tampico, se tuvo la densidad total más baja del área de estudio (648 org.100m-3) donde varias especies mostraron densidades inferiores a 200 org.100m-3, incluso aquí fue donde F. enflata presentó la densidad más baja (206 org.100m-3), mientras que Sagitta bipunctata de estirpe oceánica, indicadora de salinidades elevadas (McLelland, 1989), mostró una de las densidades más altas. En la estación 11, situada sobre la plataforma, hubo un comportamiento similar en cuanto a las densidades de estas dos especies, lo que nos hace suponer que estas dos estaciones ubicadas en la parte mas estrecha de la plataforma estuvieron influenciadas por la masa de agua oceánica, lo que coincide con el análisis de componentes principales ya que la estación nueve se unió a las oceánicas (Fig. 5).
Los datos de Valor de Importancia (Tabla 1), señalan a F. enflata como la especie más importante en la zona, dada su calidad de cosmopolita; en segundo lugar se sitúa a S. tenuis confirmando su estirpe eurihalina y nerítica (Alvariño, 1969; McLelland, 1989) y en tercer lugar a Serratosagitta serratodentata la cual puede ser considerada como indicadora de aguas del talud y corrientes del Golfo al igual que F. enflata (Alvariño, 1965). De acuerdo con McLelland (1989) esta última especie puede tolerar bajas salinidades, lo que podría explicar su amplia distribución en esta zona, con las más altas densidades en las estaciones localizadas sobre la plataforma. Para la primavera de 1986, Mille-Pagaza y Carrillo-Laguna (2001) reportan a Sagitta tenuis como integrante del segundo nivel de importancia, esta diferencia podría ser atribuible a que predominaron las estaciones oceánicas en abril-mayo de 1986.

En el segundo nivel de importancia aparecen cuatro especies con valores entre 13.4 y 15.4; este conjunto está conformado por especies de estirpe oceánica, indicadoras de mezcla de aguas costeras y oceánicas (Alvariño, 1969; McLelland, 1989) o bien, que se encontraron en sitios cercanos al talud en la zona oceánica. El tercer nivel lo conformaron el resto de las especies, de origen claramente oceánico, cuyos valores son inferiores a 8.6; incluso algunas de ellas no llegan a uno.
La clasificación de Dajoz (1996) aplicada a las especies registradas en el área de estudio (Tabla 1), confirma la posición de éstas en el cuadro de Valor de Importancia, aparecen en los dos primeros niveles las catalogadas como constantes, ya que se recolectaron en más del 50% de las estaciones de muestreo, en el tercer nivel se reúnen: Flaccisagitta hexaptera (d'Orbigny, 1843) y Krohnitta pacifica (Aida, 1897), la primera es una especie constante y la segunda ocasional, además de las accidentales que aparecieron en menos del 25% de las estaciones.
Los valores de diversidad de Shannon variaron desde 0.95 bits individuo-1 en la estación tres, ubicada frente a la boca sur de la Laguna Madre, donde S. tenuis alcanza el segundo valor de densidad más alto, comparte el área fundamentalmente con Flaccisagitta enflata cuya abundancia es de las más bajas, y otras tres especies con densidades inferiores a 100 org.100m-3. La dominancia de la primera se ve confirmada con el valor de Simpson (0.609) para esa estación. La diversidad más alta (entre 1.77 y 2.38 bits individuo-1) se encontró en los sitios de colecta situados al sur del área en las estaciones oceánicas, donde no se observa una clara dominancia por parte de ninguna de las especies.
Por el contrario los valores más altos de dominancia (de 0.39 hasta 0.75) se sitúan al norte de la zona de estudio donde la densidad de Flaccisagitta enflata impera sobre la de las otras especies, o bien, como sucede en la estación tres, la densidad de Sagitta tenuis es la más elevada. En las estaciones dos y cuatro se observan valores medios de diversidad y dominancia donde hubo una codominancia entre estas dos especies.
El dendrograma obtenido para la Similitud de Morisita separó dos conjuntos de estaciones con un nivel de corte ubicado de manera arbitraria a 0.7 de semejanza (Fig. 3). El primer grupo lo integraron ocho estaciones sureñas situadas frente a la costa de Tampico y la Laguna de Tamiahua, la mayoría de ellas colocadas en la zona oceánica o cerca del talud (Fig. 4), donde las densidades totales por estación fueron bajas. En este conjunto de estaciones una abundancia de 1000 org.100m-3 representó el valor más bajo de F. enflata y se registró el más alto de Serratosagitta serratodentata que fue de 1021 org.100m-3, la diversidad media fue alta (2.2 bits individuo-1) y por tanto la dominancia promedio fue baja (0.27). El segundo grupo estuvo integrado por siete estaciones norteñas y la más sureña, ubicadas sobre la plataforma y una en el veril, con densidades totales por estación elevadas; con un claro predominio de F. enflata o de S. tenuis sobre el resto de las especies; la diversidad media fue de 1.33 bits individuo-1 y la dominancia promedio de 0.507.

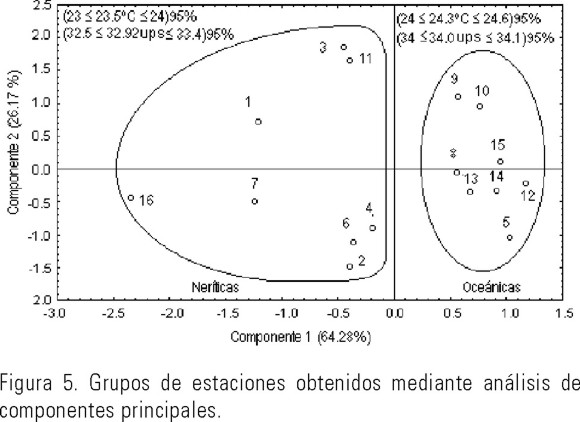
El análisis de componentes principales aplicado a los valores de los factores abióticos separó las estaciones en dos grupos: neríticas y oceánicas, con una varianza explicada del 90.45 % en los dos primeros ejes (Fig. 5). Los intervalos de confianza (=0.05) tanto de temperatura (nerítico: 23 ≤ 23.5ºC ≤ 24; oceánico 24 ≤ 24.3ºC ≤ 24.6) como de salinidad (nerítico: 32.5 ≤ 32.92ups ≤ 33.4; oceánico 34 ≤ 34.05ups ≤ 34.1) no se sobrepusieron entre grupos lo que indica que se trata de dos masas de agua con características diferentes.
El estadístico T2 de Hotelling aplicado a las variables abióticas (p = 0.00394) mostró que las densidades y la composición de especies de la comunidad de quetognatos probablemente estuvieron influenciadas por la temperatura y la salinidad (Fig. 6). En un trabajo previo los autores (Mille-Pagaza & Carrillo-Laguna, 2001) determinaron también a la temperatura como un factor que influye la composición y abundancia de los quetognatos. La concentración de oxígeno no fue diferente (p = 0.6117) entre los dos grupos de estaciones.

El hecho de que las estaciones agrupadas por similitud y componentes principales sean básicamente las mismas, así como que exista coincidencia entre los factores que contribuyeron con mayor varianza en el análisis de componentes principales y los detectados mediante la prueba T2 de Hotelling, permiten establecer que la composición y abundancia de quetognatos se ve influenciada en buena medida por el ambiente abiótico y que, en el área, confluyen dos masas de agua con características diferentes en temperatura y salinidad, las cuales corresponden esencialmente a dos ambientes marinos distintos: el nerítico y el oceánico.
Agradecimientos
Los autores agradecemos al Instituto Nacional de Pesca la donación de las muestras y a los revisores anónimos por sus atinadas observaciones.
Referencias
ÁLVAREZ-CADENA, J. N., E. SUÁREZ-MORALES & J. A. MCLELLAND. 1996. Observations on an isolated population of Sagitta hispida Conant (Chaetognatha) in a tropical lagoon system of northeast Yucatan (Mexico). Gulf Research Reports 9(3):197-204. [ Links ]
ALVARIÑO, A. 1965. Chaetognaths. Annual Review Oceanography Marine Biology 3:115-194. [ Links ]
ALVARIÑO, A. 1968. Los quetognatos, sifonóforos y medusas en la región del Atlántico ecuatorial bajo la influencia del Amazonas. Anales del Instituto de Biología, Universidad Nacional Autónoma de México 39, Serie Ciencias del Mar y Limnología 1:41-76. [ Links ]
ALVARIÑO, A. 1969. Los quetognatos del Atlántico. Distribución y notas esenciales de sistemática. Trabajos Instituto Español Oceanografía. Madrid 37: 209 p. [ Links ]
DAJOZ, R. 1996. Précis d'écologie. Dunod, Paris. 290 p. [ Links ]
GASCA, R. J. N. ÁLVAREZ-CADENA & E. SUÁREZ-MORALES. 1996. Chaetognath assemblages in the Mexican Caribbean Sea (1991). Caribbean Marine Studies 5: 1-10. [ Links ]
KREBS, J. C. 1994. Ecology. The Experimental Analysis of Distribution and Abundance. 4th ed. Harper Collins College Publishers, New York. 900 p. [ Links ]
KREBS, J. C. 1999. Ecological Methodology. Addison Wesley Longman, Inc. California. 605 p. [ Links ]
LUDWIG, J. A. & J. F. REYNOLDS. 1988. Statistical Ecology. Wiley, New York. 337 p. [ Links ]
MCLELLAND, J. A. 1989. An ilustrated key to the Chaetognatha of the northern Gulf of Mexico with notes on their distribution. Gulf Research Reports 8(2): 145-172. [ Links ]
MCLELLAND, J. A. & H. M. PERRY. 1989. Records of deep-water chaetognaths from the northern Gulf of Mexico. Gulf Research Reports 8(2): 181-187. [ Links ]
MICHEL, H. B. & M. FOYO. 1976. Caribbean zooplankton. Part 1 -Siphonophora, Heteropoda, Copepoda, Euphausiacea, Chaetognatha and Salpidae. Office of Naval Research Departament of Navy, U.S. Govt. Printing Office, Washington, D.C. 549 p. [ Links ]
MILLE-PAGAZA, S. & J. CARRILLO-LAGUNA. 1999. Los quetognatos (Chaetognatha) del banco de Campeche en abril-mayo de 1986. Revista de Biología Tropical 47 (1-2): 101-108. [ Links ]
MILLE-PAGAZA, S. & J. CARRILLO-LAGUNA. 2001. The Chaetognatha of the southwestern Gulf of Mexico during April-May, 1986. Gulf and Caribbean Research 13: 59-65. [ Links ]
MILLE-PAGAZA, S., R. REYES-MARTÍNEZ & M. E. SÁNCHEZ-SALAZAR. 1997. Distribution and abundance of Chaetognatha on he Yucatan Shelf during May, 1986. Gulf Research Reports 9: 263-275. [ Links ]
MORRISON, D. F. 1990. Multivariate Statistical Methods. McGraw-Hill Publishing Co., Singapore. 495 p. [ Links ]
MOULIN-RUÍZ, J. 1980. Observaciones hidrobiológicas frente al litoral de Tamaulipas en el Golfo de México. Secretaría de Marina, Dirección General de Oceanografía. México. 48 p. [ Links ]
ORESLAND, V. 2000. Diel feeding of the chaetognath Sagitta enflata in the Zanzibar Channel, western Indian Ocean. Marine Ecology Progress Series 193: 117-123. [ Links ]
OWRE, H. B. 1960. Plankton of the Florida Current. Part IV: The Chaetognatha of the northern of the Gulf of Mexico. Bulletin of Marine Science 10 (3): 255-322. [ Links ]
PIERCE, E. L. 1962. Chaetognatha from the Texas coast . Publications of the Institute of Marine Science, University of Texas 8: 147-152. [ Links ]
PIERCE, E. L. & M. L. WASS. 1962. Chaetognatha from the Florida Current and coastal water of the southeastern Atlantic states. Bulletin of Marine Science 12: 403-431. [ Links ]
RIVERO-BELTRÁN, C. 1975. Distribución de los quetognatos en la Bahía de Campeche. Memorias 1 Simposium Latinoamericano Oceanografía Biológica (México): 306-324. [ Links ]
SMITH, R. L. & T. M. SMITH. 2001. Ecología. 4ª Ed. Adison Wesley, España. 642 p. [ Links ]
STUART, V. & H. M. VERHEYE. 1991. Diel Migration and feeding patterns of the chaetognath, Sagitta friderici, of the west coast of South Africa. Journal of Marine Research 49: 493-515. [ Links ]
SUÁREZ-CAABRO, J. A. 1955. Quetognatos de los mares Cubanos. Memorias Sociedad Cubana Historia Natural 22:125-180. [ Links ]
ULLOA, R. H. & S. PALMA. 2000. Distribución vertical de los quetognatos y su relación con eventos de surgencia pp. 59-60. In: RÍOS JARA, J. E., E. SUÁREZ CARRILLO, M. PÉREZ PEÑA, E. LÓPEZ URIARTE, E. G. ROBLES JARERO, D. U. HERNÁNDEZ BECERRIL & M. SILVA BRIANO (Eds.). Estudios sobre plancton en México y el Caribe. Sociedad Mexicana de Planctología, A. C. (SOMPAC). México. [ Links ]
VÁZQUEZ DE LA CERDA, A. M. 1975. Currents and waters of the upper 1200 meter of the southwestern Gulf of Mexico. M. S. thesis. Texas A & M University, College Station, TX. 180p. [ Links ]
VEGA-RODRÍGUEZ, F. 1965. Distribución de Chaetognatha en Veracruz, Ver. Anales Instituto Biología Universidad Nacional Autónoma México, Serie Zoología 36: 229-247. [ Links ]

