










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.13 no.3 Ciudad de México oct. 2003
Artículo
Variabilidad del estado trófico y la biomasa del fitoplancton de Bahía Concepción, Golfo de California (1997-1999)
Variability of the trophic state and phytoplankton biomass of Bahia Concepcion, Gulf of California (1997-1999)
1David J. López-Cortés, 2Ismael Gárate-Lizárraga, 1José J. Bustillos-Guzmán, 1Rosalba Alonso-Rodríguez e 1Iban Murillo-Murillo.
1Centro de Investigaciones Biológicas del Noroeste S.C. (CIBNOR). Apdo. Postal 128, C.P. 23000, La Paz, Baja California Sur, México.
2Centro Interdisciplinario de Ciencias Marinas (IPN-CICIMAR). Laboratorio de Fitoplancton, Apdo. Postal 592, C.P. 23000, La Paz, Baja California Sur, México. E-mail:dlopez@cibnor.mx
Recibido: 8 de septiembre de 2002.
Aceptado: 10 de septiembre de 2003.
Resumen
Con el propósito de determinar la variación de la biomasa del fitoplancton (clorofila a) y el estado trófico de Bahía Concepción ubicada en la parte central oeste del Golfo de California, se realizó un estudio hidrobiológico durante el período de 1997 a 1999. Se determinó la temperatura, la concentración del oxígeno disuelto y de nutrientes inorgánicos (nitrato, nitrito, fosfato y silicato), y la biomasa fitoplanctónica (clorofila a). Las condiciones hidrográficas muestran un período de mezcla (octubre-abril), dos de transición (abril-mayo, septiembre-octubre) y uno de estratificación (junio-septiembre). Las temperaturas más cálidas durante el periodo de estudio fueron observadas durante 1998 y las más frías en 1997 para todos los períodos hidrográficos. Los valores de oxígeno disuelto muestran una zona de hipoxia durante el período estratificado (< 2.7±0.78 ml l-1), por abajo de los 20 metros de profundidad. Durante los períodos de transición se observó un incremento de la biomasa fitoplanctónica similar al que se observa como máximos de primavera y verano de algunas lagunas costeras en latitudes similares, sin embargo la mayor biomasa se detectó en el período de estratificación, particularmente por abajo de los 15 m. La escala de nutrientes mostró un estado de eutrofía para fosfato y silicato todo el año. El índice trófico reveló que la oligotrofía para fosfatos, nitratos y nitritos es el estado dominante, con excepción de los períodos de transición, que es cuando la mesotrofía se presentó. Los resultados sugieren que los incrementos fitoplanctónicos en Bahía Concepción se deben al suministro de nutrientes de origen natural y que las condiciones tróficas (mesotrofía o eutrofía), son determinadas por las condiciones hidrográficas del área.
Palabras clave: Biomasa de fitoplancton, estado trófico, Bahía Concepción, Golfo de California.
Abstract
To determine the effect of the phytoplankton biomass and the trophic status in Bahía Concepción in the central-west part of the Gulf of California an hydrobiologic study was conducted from 1997 to 1999. Temperature, dissolved oxygen, inorganic nutrients (nitrate, nitrite, phosphate and silicate) and phytoplankton biomass (chlorophyll a) concentrations were measured. Hydrographic conditions shown a mixing period (October-April), two transition periods (April-May and September-October) and a stratification period (June-September). Warmest temperatures were observed during 1998 and coldest in 1997. Dissolved oxygen values shown an hypoxic layer (<2.7±0.78 ml l-1), during the stratified period below 20 m depth. During the transition period two peaks of phytoplankton biomass were observed, on spring and summer, which matched with those reported for coastal lagoons at similar latitudes, however the highest biomasses were observed in the stratification period, particularly below 15 m depth. The scale of nutrients showed an eutrophy for phosphate and silicate all the year. The trophic index revealed an oligotrophy state for phosphate, nitrate and nitrite had a dominant status with the exception for the transition periods, when the mesotrophic conditions were presents. These results suggest that phytoplankton biomass is associated with the natural inputs of nutrients and that trophic conditions (mesotrophy or eutrophy) were determined for the hydrographic of the area.
Key words: Phytoplankton biomass, trophic status, Bahía Concepción, Gulf of California.
Introducción
En general en los ecosistemas marinos semicerrados como las lagunas costeras, esteros y bahías, los flujos de marea, vientos locales, surgencias y el aporte de agua dulce proveniente de ríos, generan condiciones hidrográficas de mezcla y estratificación de la columna de agua (Simpson et al., 1977; Viner, 1985; Kiørboe, 1993), provocando cambios en el balance de nutrientes y en el estado trófico de estos ecosistemas (Figueiras et al., 1986; Peterson & Bellantoni, 1987; Scharek et al., 1994). Un incremento de los niveles de nutrientes en la columna de agua de bahías y esteros, se asocia a los procesos de mezcla (Delgadillo-Hinojosa et al., 1997; Casas et al., 1999) y una disminución de nutrientes en superficie e incrementos por hundimiento y acumulación por abajo de la termoclina en cuerpos de agua profundos se observa bajo condiciones de estratificación (Pingree et al., 1977; Viner, 1985; Casas & Bode, 1999).
La eutroficación o eutrofización es un proceso asociado al enriquecimiento de nutrientes y/o la introducción de material orgánico en los cuerpos de agua. En los últimos años, el incremento de la eutrofización de cuerpos de agua ha sido provocada por el aumento en el suministro de nutrientes a nivel global, proveniente de actividades humanas, lo cual promueve un exceso en la productividad primaria y en algunos casos se generan eventos de hipoxia o anoxia en el fondo de los cuerpos de agua (Justic et al., 1995). Las clasificaciones del estado trófico de ecosistemas marinos se basan en divisiones discretas y arbitrarias (Ignatiades et al., 1992; Alonso-Rodríguez et al., 2000). Por ello se ha elegido una escala (Ignatiades et al. 1992; Smith et al. 1999) y el índice de Karydis et al. (1983). La escala propone la existencia de tres estados tróficos: El estado eutrófico (0.35±0.22 de N-NO3, 0.34±0.28 P-PO4, 1.15±0.90 N-NH3 y 3-5 mg-Chl a m-3) se refiere al enriquecimiento de nutrientes o materia orgánica y está asociado a la presencia de fitoplancton nocivo e hipoxia. El estado mesotrófico (0.26±0.17 de N-NO3, 0.09±0.05 de P-PO4, 0.84±0.47 de N-NH3 y de 1-3 mg-Chl a m-3) es definido como un estado intermedio, relacionado a la concentración de nutrientes y crecimiento fitoplanctónico (Ignatiades et al., 1992; Alonso-Rodríguez et al., 2000). Por último, el estado oligotrófico (0.23±0.21 de N-NO3, 0.03±0.03 de P-PO4, 0.38±0.23 de N-NH3 y de <1 mg-Chl a m-3) se refiere a la ausencia de concentraciones de nutrientes que puedan ser detectadas y a una producción primaria limitada.
Bahía Concepción es un ecosistema costero semicerrado, antiestuarino, en contacto directo con agua superficial de la parte central del Golfo de California (Gilmartin & Revelante, 1978; Martínez-López & Gárate-Lizárraga, 1997). Durante el año se presentan tres condiciones hidrográficas: la de mezcla en otoño-invierno, la de estratificación en primavera-verano y dos períodos de transición que varían según la intensidad y dirección de los vientos (Lechuga-Devéze et al., 2000; López-Cortés et al., 2003). Estas condiciones hidrográficas pueden provocar cambios importantes en los flujos de nutrientes, influyendo en el crecimiento y proliferación de diversos grupos fitoplanctónicos (Viner, 1985). Se ha sugerido, a través del análisis de los parámetros fotosintéticos, que en la parte central del Golfo de California las condiciones tróficas no corresponden a la oligotrofía (Gaxiola-Castro et al., 1999), sin embargo, aún no se ha definido su estado trófico. En este trabajo se evalúa la biomasa de fitoplancton y se hace una aproximación del estado trófico de bahía Concepción basándose en el análisis de la concentración de clorofila a y nutrientes obtenidos de 1997 a 1999 y la aplicación de índices tróficos con validez universal en aguas costeras de cuerpos semicerrados como es el caso. El objetivo del presente trabajo es el de asociar los incrementos del fitoplancton con los diferentes estados tróficos del sistema.
Materiales y métodos
Durante tres años consecutivos (1997, 1998 y 1999) se realizaron muestreos en dos estaciones ubicadas en la parte central y más profunda de Bahía Concepción (Fig. 1). Se tomaron muestras de agua con una botella van Dorn en la superficie, 5, 10, 15, 20, 25 y 27 m de profundidad. Para la determinación de los parámetros físico-químicos en 1997 se colectaron muestras de agua de febrero a mayo cada tres a cinco días. En 1998 se intensificaron los muestreos a cada tres días, durante los primeros diez meses del año y cada 15 días para el resto de este mismo año. Durante 1999 los muestreos se realizaron cada 15 días. La temperatura del agua se midió con un termómetro de cubeta (Kalshico). El oxígeno se determinó mediante el método de Winkler (Strickland & Parsons, 1972). Para determinar la biomasa total de fitoplancton (clorofila a) se filtraron 1.5 litros de agua de mar, utilizando filtros de fibra de vidrio Whatman GF/F, los cuales se congelaron a -20°C y posteriormente se analizaron mediante cromatografía líquida de alta resolución, HPLC (Bustillos-Guzmán et al., 1995). También se obtuvieron muestras en cada profundidad para la cuantificación de nutrientes inorgánicos; nitrato, nitrito, fosfato y silicato, los cuales se determinaron siguiendo la metodología propuesta por Strickland y Parsons (1972).
El índice de estratificación térmica IET (°C m-1) se calculó con base en la diferencia térmica del estrato superior e inferior dividido entre su correspondiente profundidad y entre el número de profundidades menos uno (Bustillos-Guzmán et al., 1995) generándose la siguiente escala: IET<0.05 indica mezcla; 0.05>IET<0.2 indica transición e IET>0.2 indica estratificación. Los datos de vientos fueron proporcionados por la Comisión Nacional del Agua (CNA) y se expresan como el cubo de la velocidad del viento (m3 s-1) el cual se considera como un buen indicador de la mezcla en la columna de agua de acuerdo con Gómez-Gutiérrez et al. (1999).
Se estimó el índice trófico mensual I para fosfato, nitrato y nitrito con base a la siguiente ecuación: I (C/C-log x) Log A (Karydis et al.,1983):
donde:
I : es el índice trófico para un nutriente específico.
C : es el logaritmo base 10 del total (para este trabajo se utilizaron los valores integrados de todos los meses, para cada año) de la carga de un determinado nutriente en una área y x es la concentración total (anual) de ese nutriente para la estación muestreada (Estaciones 1 y 2).
A: Es el número de estaciones.
La escala de clasificación es la siguiente: si I > 5 indica estado eutrófico, 3 > I < 5 indica estado mesotrófico e I < 3 indica estado oligotrófico.
Para cada época del año se determinó el estado trófico de la bahía mediante la utilización de la escala de concentración de nutrientes propuesta por Ignatiades et al. (1992). Para este análisis se emplearon los datos de nutrientes correspondientes a los niveles de superficie y 15 metros de profundidad.
Resultados
Índice de Estratificación. De acuerdo con los resultados obtenidos con el índice de estratificación térmica IET (°C m-1) se registraron cuatro períodos hidrográficos para los tres años estudiados: uno de mezcla, uno estratificado y dos de transición (mezcla-estratificación). Estos períodos estuvieron marcadamente influenciados por el viento y el calentamiento del agua de mar (Fig. 2) y presentaron variaciones anuales e interanuales en magnitud, intensidad y tiempo. Aunque para 1997, solo fueron monitoreados los primeros cuatro meses del año, se registró una parte del período de mezcla (27 de febrero al 12 de marzo), coincidiendo con incrementos en la velocidad del viento (1400 m3 s-1). Un período de transición del 14 de marzo al 5 de mayo y el inicio y desarrollo de una columna de agua estratificada en la segunda semana de mayo (Fig. 2A). A principios de 1998 se incrementó la frecuencia de los vientos de máxima velocidad (1400 m3 s-1) prolongando su acción hasta finales de abril, en este año los dos períodos de mezcla abarcaron la época de otoño-invierno (31 de enero al 16 de abril y del 17 de octubre al 28 de noviembre), posteriormente disminuyó la velocidad del viento (600 m3 s-1) y se inició el primer período de transición que fue más extenso que el del año anterior (20 abril-28 de junio) y el segundo, que comprendió del 6 de septiembre al 2 de octubre. Con la reducción del efecto del viento y el incremento en el calentamiento del agua, se originó el período de estratificación de Bahía Concepción (30 junio-30 agosto) (Fig. 2B). En general, durante 1999 se observó una disminución en la velocidad del viento y un acortamiento en la duración de los períodos de transición, además de un alargamiento del período de estratificación (Fig. 2C).
Temperatura. Las temperaturas más bajas se registraron durante los períodos de mezcla, siendo el período de 1997 el más frío (18.0°C) al ser comparado con 1998 y 1999 (21.0°C) (Fig. 3A). El período de transición más cálido fue durante 1998 y 1999 (>25°C de 0-15 m) con respecto a 1997 (22.0°C de 0-15 m), aún por abajo de los 15 m, 1998 fue mas cálido (25-24°C) que 1997 (19-20°C) y que 1999 (25-22°C) (Fig. 3B). Las más altas temperaturas se registraron en superficie durante el verano, coincidiendo con los períodos de estratificación de la columna de agua. Así en 1998 se presentaron temperaturas elevadas desde superficie a 27 m (30-25°C) comparadas con 1997 (27-20°C) y 1999 (30-23°C). La termoclina se desarrolló entre los 5 y los 20 m de profundidad en 1997 y entre los 10 a 20 m en 1998 y 1999 (Fig. 3C).
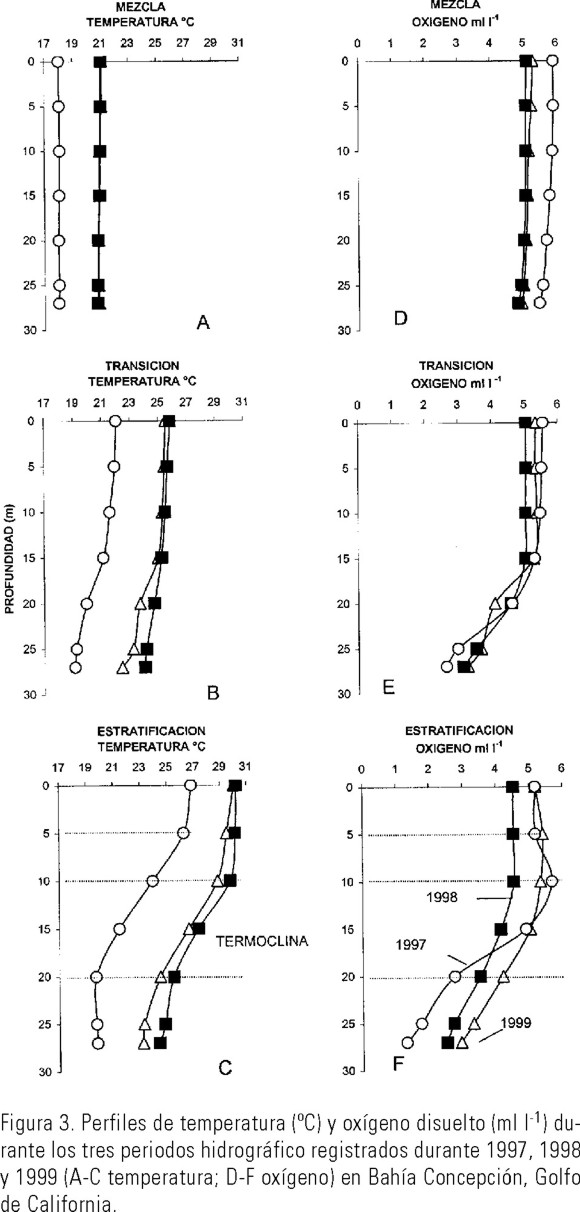
Oxígeno disuelto. Los valores de oxígeno disuelto en el período de mezcla fueron elevados en los tres años, con concentraciones mayores de 5.0 ml l-1, principalmente en 1997 (Fig. 3D). Durante el período de transición, también se presentaron valores altos desde la superficie hasta los 15 m, sin embargo, por abajo de esta profundidad se inició la oxiclina en los tres años estudiados (Fig. 3E). Durante el período de estratificación, en los tres años, las condiciones de hipoxia se registraron por abajo de los 25 m, con valores >1.0<3.0 ml l-1. (Fig. 3F).
Nutrientes. Durante el periodo de mezcla, los perfiles de nitrato mostraron concentraciones homogéneas en toda la columna de agua, excepto en 1998, donde se observó un pequeño incremento por abajo de los 25 m (Fig. 4A), en la tabla 1 se observa un valor medio mayor (0.33±0.21) comparado con el de 1997 (0.17±0.10 µM) y 1999 (0.23±0.22 µM) (Tabla 1).
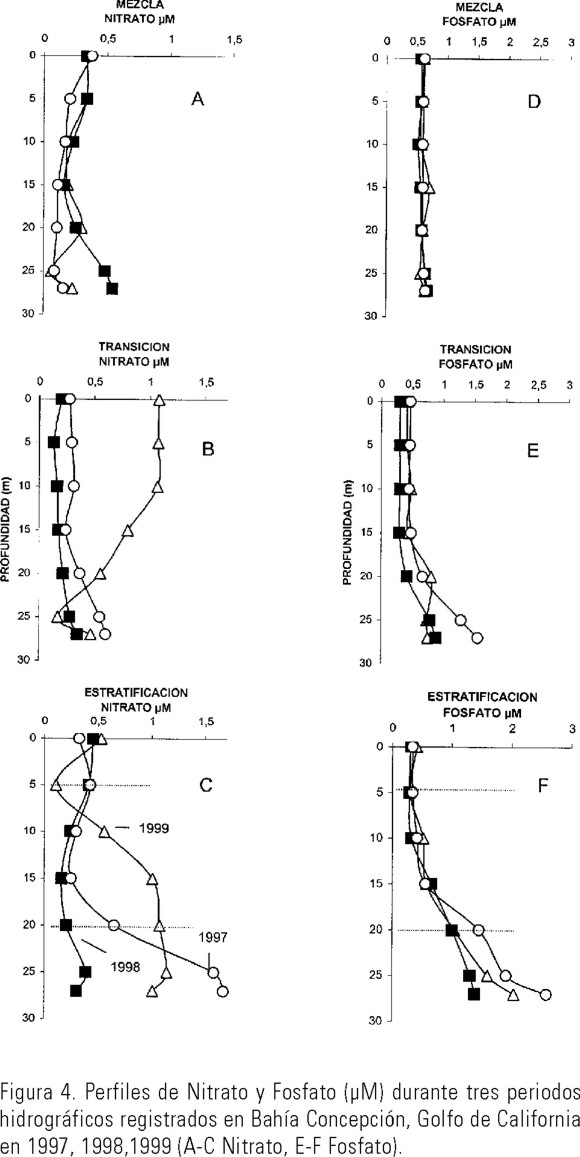
En el período de transición hidrográfica se cuantificaron concentraciones de nitrato mayores de 1 µM-NO3 en 1999 disminuyendo por abajo de los 10 m, en cambio se determinaron las menores concentraciones de nitrato en 1998 (Fig. 4B). Durante la estratificación hubo incrementos por abajo de 5 y 15 m en 1999 y 1997 respectivamente, en cambio en 1998 se mantuvo toda la columna de agua con concentraciones menores de 0.5 µM-NO3 (Fig. 4C). Las concentraciones de nitrito en los tres años analizados fueron elevadas por abajo de 20 m de profundidad (0.21-0.35 µM-NO2) durante el período de estratificación (Tabla 1).
Las concentraciones de fosfato no presentaron variaciones importantes durante los períodos de mezcla en los tres años (Fig. 4D). Sin embargo en el período de transición se incrementaron los valores de fosfato por abajo de los 20 m de profundidad en los tres años analizados (Fig. 4E), aunque en 1998 se registró para este periodo el valor medio más bajo de 0.67±0.5 µM-PO4 comparado con 1997 y 1999 (Tabla 1). Los valores más elevados de fosfato en estos tres años se registraron en los períodos de estratificación, particularmente por abajo de los 15 m. Sin embargo, en 1998, en este mismo periodo las concentraciones fueron bajas con valores menores de 1.5 µM-PO4 por abajo de la termoclina, comparadas con 1997 y 1999 donde se registraron concentraciones ≥ 2 µM-PO4 (Fig. 4F).
Las concentraciones de silicato no presentaron grandes cambios durante el período de mezcla, con valores < 10 µM-SiO4 (Fig 5A), sin embargo durante el período de transición por abajo de 20 m hubo incrementos hasta 20 µM-SiO4 (Fig. 5B) los valores medios fueron de 14.37±8.61 µM-SiO4 en 1997, de 9.97±6.15 µM-SiO4 en 1998 y de 11.56 ±5.70 µM-SiO4 en 1999. Durante el período estratificado, por abajo de 15 m las concentraciones se incrementan (Fig. 5C), las cuales siempre fueron más elevados en 1999 (22.06 ±8.17 µM-SiO4) que en 1997 (8.81±8.48 µM-SiO4) y que en 1998 (14.48±5.15 µM-SiO4) (Tabla 1).

Biomasa fitoplanctónica. De acuerdo con los perfiles de la biomasa total de fitoplancton (clorofila a), la distribución vertical fue homogénea durante el período de mezcla, observándose diferencias interanuales importantes. Los valores promedio más bajos se detectaron en 1998 (<1 µg l-1) y los más elevados se determinaron para 1997 (>1.5 µg l-1) (Fig. 5 D). Durante el período de transición no se cuantificaron diferencias interanuales importantes, no obstante, se observó un incremento de la biomasa fitoplanctónica a partir de los 10 m de profundidad (0.81-1.25 µg l-1) (Fig. 5E). La estratificación de la columna de agua trajo como resultado el incremento de la biomasa total (Fig. 5F). Este hecho es más notable durante el período de estratificación de 1997, donde se observó un máximo sub-superficial de clorofila a de 4.33 µg l-1. Para 1998-1999 éstos incrementos se observaron por abajo de la termoclina, pero no fueron tan pronunciados.
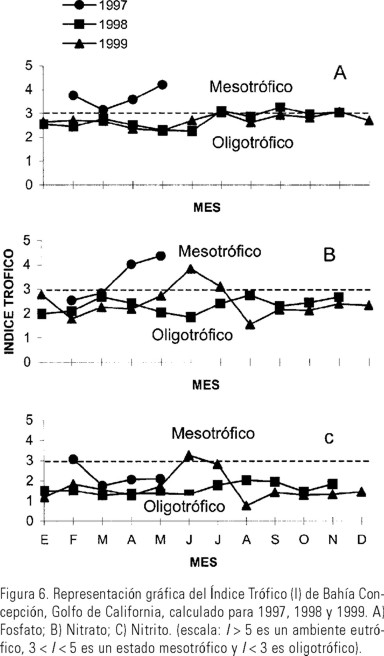
Índice trófico. En los dos estratos de la columna de agua (superficie y 15 m de profundidad), considerados para la aplicación de la escala para determinar las condiciones tróficas de cada nutriente en los diferentes períodos climáticos anuales, se encontró un predominio de la eutrofía para fosfato en las cuatro épocas de los tres años estudiados. El nitrato mostró condiciones principalmente eutróficas y mesotróficas. La oligotrofía generalmente predominó con respecto al nitrito (Tabla 2).
La aplicación de la escala del índice trófico mensual, mostró que durante el período comprendido entre febrero y mayo de 1997, predominó un estado mesotrófico para el fosfato, sin embargo, durante 1998 y 1999 este período fue oligotrófico, con excepción del mes de septiembre de 1998 (Fig. 6A). Las concentraciones de nitrato en 1997 y 1999 presentaron dos estados de mesotrofia en los períodos de transición y de estratificación. En el resto del año predominó la oligotrofía, principalmente durante 1998 (Fig. 6B). El estado trófico con respecto al nitrito fue de oligotrofia en los tres años estudiados, excepto durante julio de 1999 donde hubo un pequeño incremento (Fig 6C). Las condiciones de mesotrofía se registraron principalmente durante los procesos de transición hidrográfica (mezcla-estratificación).
Discusión
Los procesos de mezcla y estratificación de la columna de agua de los ecosistemas costeros son controlados por la estacionalidad del efecto de los vientos, cambios en los flujos de mareas y la topografía (De la Lanza-Espino & Rodríguez-Medina 1993; Simpson & Rippeth, 1993; De la Lanza-Espino & Flores-Verdugo, 1998). En Bahía Concepción estos procesos de estabilidad e inestabilidad en la columna de agua son causados por la intensidad de los vientos del NO durante otoño e invierno y del SE en primavera-verano. Aunque los resultados de este trabajo coinciden parcialmente con lo descrito por Lechuga-Devéze et al. (2001), no obstante, se observó que el patrón de vientos provoca cambios anuales e interanuales en la estructura de la columna de agua, originando períodos de mezcla, de transición y de estratificación de diferente intensidad y duración.
Cuando los vientos fuertes del noroeste inciden en Bahía Concepción se desarrollan los períodos de mezcla (IET <0.05) provocando concentraciones altas de oxígeno disuelto (5.02-5.76 ml l-1) en la columna de agua. La misma situación de homogenización se presentó en las concentraciones de nitrato y fosfato, las cuales no rebasaron los 0.7 µM. Con la disminución del efecto de los vientos, la energía de la turbulencia de la columna de agua disminuye paulatinamente (Simpson et al., 1977) lo cual produce un período de transición hidrográfico, cuya duración está en función de la intensidad de los vientos que la preceden, originando aumentos en las concentraciones de nutrientes y de la biomasa fitoplanctónica (Kiørboe, 1993). Con la disminución de la velocidad de los vientos del noroeste, en Bahía Concepción se iniciaron los períodos de transición y estratificación, registrándose incrementos de nitrato, fosfato y silicato por abajo de los 20 m principalmente en 1997 y 1999. El aumento notable de NO3 y PO4 durante la estratificación trajo como consecuencia el incremento de la biomasa fitoplanctónica (clorofila a) con valores medios por encima de los registrados superficialmente en agosto de 1972 (0.4 µg-Chla l-1) en Bahía Concepción por Gilmartin y Revelante (1978). Bajo estas condiciones de estratificación hidrográfica y de nutrientes, en otras áreas del mundo también se han encontrado incrementos subsuperficiales de clorofila a (Pingree et al., 1976, 1977; Holligan et al., 1984; Revelante & Gilmartin, 1990; Kiørboe, 1993). Sin embargo en 1998, aun bajo condiciones de estratificación, las concentraciones de estos dos nutrientes y de la clorofila a, fueron bajas comparadas con 1997 y 1999, incluso en los períodos de mezcla de 1998. Esto puede explicarse por la presencia de agua ecuatorial superficial que avanzó hasta la parte central y norte del Golfo de California durante el evento El Niño 1997-1998 (Thunell et al.,1999; Lavín et al., 2003). Los datos de temperatura de este estudio muestran un retorno a condiciones más frías en 1999, probablemente asociadas con condiciones de no-Niño (McPhaden, 1999; Bograd et al., 2000; Castro et al., 2000; Lavín et al., 2003), con incrementos en Bahía Concepción de nutrientes y de biomasa fitoplanctónica (López-Cortés et al., 2003). Una disminución rápida de la temperatura así como un incremento por arriba de lo normal de nutrientes y clorofila a también fueron registrados bajo condiciones no-Niño después del evento El Niño 1997-1998, en el Pacífico Ecuatorial (Chávez et al., 1999).
El incremento de la estabilidad de la columna de agua produce una acusada estratificación térmica (Simpson et al.,1977) y la formación de la nutriclina, generándose una barrera térmica, señalada por la termoclina, por encima de la cual, los nutrientes generalmente son escasos debido al consumo y por abajo de esta barrera, se incrementan con la profundidad (Pingree et al., 1976; Karydis et al., 1983). Con el abatimiento de la fuerza eólica y el incremento de la temperatura superficial del mar, en Bahía Concepción se generó la estratificación de la columna de agua y la formación de la termoclina (10-20 m) debajo de esta barrera térmica, las concentraciones de fosfato y nitrato se incrementaron principalmente en 1997 y 1999. Delgadillo-Hinojosa et al. (1997) determinaron en una laguna costera del Golfo de California, que durante la estratificación aumentaban los nutrientes y la biomasa fitoplanctónica, estos incrementos de nutrientes durante la estratificación Babenerd (1991) los asoció con el aumento de la actividad bacteriana. Ammerman y Azam (1985) encontraron que algunas bacterias poseen enzimas en las membranas que son capaces de liberar fósforo inorgánico a partir de compuestos orgánicos, así también Stal et al. (1996), estudiando dos cuencas marinas costeras encuentra diferentes concentraciones de Fe+3 y PO4-3 atribuyéndolo a la estacionalidad del crecimiento de tapetes microbianos formados principalmente de cianobacterias. Para Bahía Concepción, Bustillos-Guzmán et al. (2000) sugieren que el predominio de bacterias Chlorobiaceas durante el verano, puede tener un papel importante en la remineralización de nutrientes, a partir del incremento de material orgánico que se sedimenta durante la estratificación, provocando un aumento natural y no antropogénico de nutrientes (Lechuga-Devéze et al., 2001). Sin embargo también se ha sugerido la influencia del agua superficial de la parte central del Golfo de California (Gilmartin & Revelante, 1978) que aunque su contenido de nutrientes es muy variable (Tabla 3), en este estudio se registraron concentraciones medias relativamente bajas en febrero-marzo de 1997-1999 en la superficie, <0.30 µM-NO3, < 0.6 µM-PO4 y < 9.0 µM-SiO4 y de < 0.78 µM-NO3, <1.20 µM-PO4, < 15.9 µM-SiO4 a 27 m, no obstante, las concentraciones de fosfato y de silicato son comparables con algunos valores señalados para el Golfo Central y la misma bahía. Es probable que el agua superficial de la parte oeste del Golfo Central, adyacente a Bahía Concepción, influya en la regulación de los aportes de nutrientes principalmente durante los procesos de mezcla y que también se generen en mayor proporción en el periodo de estratificación dentro de la bahía, por lo tanto es necesario hacer un estudio más detallado del balance de nutrientes para establecer las cargas que se importan o se exportan en este ecosistema costero.
Los excedentes del material orgánico que se generan por la proliferación del fitoplancton propician además de la liberación de nutrientes, el incremento en las concentraciones de sulfuro de hidrógeno (H2S) (Bustillos-Guzmán et al., 2000; Lechuga-Devéze et al., 2001) generándose, a finales del verano y mediados del otoño, un ambiente subóxico o anóxico propicio para los procesos de desnitrificación (Palmer et al., 2000), lo cual es posible inferir que se lleven a cabo debido al incremento de nitrito en el fondo de la bahía durante el periodo de estratificación (Tabla 1). Goering y Cline (1970) y Cline y Richards (1972) demuestran que las máximas concentraciones de nitrito en el ambiente marino son originadas principalmente por la desnitrificación bacteriana.
Aunado al incremento de nutrientes durante la estratificación, las concentraciones de oxígeno disuelto disminuyen con la profundidad hasta llegar a estados de hipoxia (1-2 mg l-1) o anoxia (0-1 mg l-1) e incrementos de sulfuro de hidrógeno de hasta 1.63 mg l-1 en el otoño (Lechuga-Devéze et al., 2001). En este trabajo se tiene evidencia acerca de la presencia de sulfuro de hidrógeno en este período hidrográfico (datos no publicados) y la presencia de eventos de hipoxia, los cuales se presentaron en 1997 abajo de los 20 m y para los años de 1998 y 1999 por abajo de 25 m. Es probable que las condiciones de hipoxia y anoxia de este estrato (20-30 m) sean el resultado del hundimiento y sedimentación de materia orgánica generada por la proliferación masiva de fitoplancton durante la época de invierno (Gárate-Lizárraga et al., 2001a) y su posterior sedimentación durante el verano.
El incremento en la velocidad del viento a principios de otoño, aunado al enfriamiento del agua superficial de Bahía Concepción, provocó el rompimiento de la termoclina en el mes de septiembre de 1998 y 1999, dando lugar al segundo periodo de transición hidrográfica. Este hecho se corrobora con la disminución del índice de estratificación térmica (IET < 0.2) y la resuspensión de nutrientes, originando condiciones para la proliferación rápida del fitoplancton, originando el máximo fitoplanctónico característico de lagunas costeras. Estas particularidades hacen que los periodos hidrográficos de transición presenten condiciones propicias para el desarrollo del fitoplancton lo cual trae como consecuencia una elevada biomasa (Robinson, 1970; Kiørboe 1993; Gárate-Lizárraga et al., 2001b).
Bahía Concepción ha sido descrita como un ecosistema costero, con escasos aportes de nutrientes de origen terrígeno y antropogénico (Bustillos-Guzmán et al, 2000) y básicamente oligotrófico en el verano (Gilmartin & Revelante, 1978). Los resultados de este trabajo muestran variaciones en las concentraciones de nutrientes, lo cual da como resultado distintos estados tróficos en las diferentes épocas del año. Esto hace que se presente un proceso de eutrofia natural para fosfato y silicato durante todas las épocas del año y de mesotrofía y oligotrofía con respecto a nitrato y nitrito. De acuerdo con el criterio de Smith et al. (1999) y considerando las concentraciones media de clorofila a estimadas durante los tres periodos hidrográficos, Bahía Concepción presentó estados de oligotrofia en 1998 y de mesotrofia en 1997 y 1999. En conclusión: los periodos hidrográficos en Bahía Concepción presentaron una marcada estacionalidad, sin embargo su duración y magnitud mostró variabilidad interanual, debido a: la intensidad de los vientos del NO, la influencia de agua superficial de la parte central del Golfo de California y a eventos de mayor escala como El Niño-La Niña. Bahía Concepción es un ecosistema básicamente oligotrófico la mayor parte del año, excepto durante los períodos de transición hidrográfica en los cuales se presentan estados de mesotrofía los cuales propician incrementos de la biomasa fitoplanctónica. Un estado de eutrofía para fosfato y silicato se registró para las diferentes épocas de los años analizados. Estos estados tróficos son promovidos principalmente por: a) el efecto de los vientos, b) los aportes del agua provenientes del oeste de la parte central del Golfo de California y c) por procesos de desnitrificación que generan incrementos de nitrito y fosfato por abajo de la termoclina, como consecuencia de la acumulación de material orgánico en el fondo de la bahía, produciéndose como resultado eventos de hipoxia y anoxia. Se concluye también que el incremento de la biomasa fitoplanctónica esta regulado por los flujos naturales de nutrientes.
Agradecimientos
Este trabajo fue financiado por el CONACyT mediante los proyectos 007PÑ-1297 y 33684-V; por el CIBNOR mediante los proyectos GEA-3 y GEA-11. Ismael Gárate Lizárraga es Becario COFAA, EDI y CONACyT (Beca 138138); proyecto CGPI 2002-03-73. A Clara Ramírez Jáuregui por su valiosa colaboración en la adquisición del acervo bibliográfico.
Referencias
ALONSO-RODRÍGUEZ, R., F. PÁEZ-OSUNA & R. CORTÉS-ALTAMIRANO. 2000. Trophic conditions and stoichiometric nutrients balance in subtropical waters influenced by municipal sewage effluents in Mazatlán Bay (SE Gulf of California). Marine Pollution Bulletin 40(4): 331-339. [ Links ]
AMMERMAN, J.W. & F. AZAM. 1985. Bacterial 5-nucleotidase in aquatic ecosystems: A novel mechanism of phosphorus regeneration. Science 227: 1338-1340. [ Links ]
BABENERD, B. 1991. Increasing oxygen deficiency in Kiel Bay (Western Baltic): A paradigma of progressing coastal eutrophication. Meeresforsch 33: 121-140. [ Links ]
BOGRAD, J. S., P. M. DIGIACOMO, R. DURAZO, T .L. HAYWARD, K. D. HYRENBACH, R. J. LYNCH, A.W. MANTYLA, F. B. SCHWING, W. J. SYDEMAN, T. BAUMGARTNER, B. LAVANIEGOS & C. S. MOORE. 2000. The state of the California Current, 1999-2000: Forward to a new regime?. California Cooperative Oceanic Fisheries Investigation Report 41: 26-52. [ Links ]
BUSTILLOS-GUZMÁN, J., H. CLAUSTRE & J. C. MARTY. 1995. Specific phytoplankton signatures and their relationship to hydrographic conditions in the coastal North-western Mediterranean Sea. Marine Ecology Progress Series 124: 247-258. [ Links ]
BUSTILLOS-GUZMÁN, J., D. J. LÓPEZ-CORTÉS, F. HERNÁNDEZ & I. MURILLO. 2000. Pigment signatures associated with an anoxic coastal zone: Bahia Concepcion, Gulf of California. Journal of Experimental Marine Biology and Ecology 249: 77-88. [ Links ]
CASAS, B., M. VARELA & A. BODE. 1999. Seasonal succession of phytoplankton species on the coast of A Coruña (Galicia, northwest Spain). Boletin del Instituto Español de Oceanografía 15(1-4): 413-429. [ Links ]
CASTRO, R., A. S. MASCARENHAS, R. DURAZO & C. A. COLLINS. 2000. Variación estacional de la temperatura y salinidad en la entrada del Golfo de California, México. Ciencias Marinas 26(4): 561-583. [ Links ]
CLINE, J. D. & F. A. RICHARDS. 1972. Oxygen deficient conditions and nitrate reduction in the Eastern Tropical North Pacific Ocean. Limnology and Oceanography 17(6): 885-900. [ Links ]
CHAVEZ, F. P., P. G. STRUTTON, G. E. FRIEDERICH, R. A. FEELY, G. C. FELDMAN, D. G. FOLEY & M. J. PHADEN. 1999. Biological and chemical response of the Equatorial Pacific Ocean to the 1997-1998 El Niño. Science 286: 2126-2131. [ Links ]
DE LA LANZA-ESPINO, G. & M. A. RODRIGUEZ-MEDINA. 1993. Nutrient exchange between Subtropical lagoons and the marine environment. Estuaries 16 (2): 273-279. [ Links ]
DE LA LANZA-ESPINO, G. & F. FLORES-VERDUGO. 1998. Nutrient fluxes in sediment (NH4+ and PO3-3) in N.W. coastal lagoon Mexico associated with an agroindustrial basin. Water, Air, and Soil Pollution 107: 105-120. [ Links ]
DELGADILLO-HINOJOSA, F., G. GAXIOLA-CASTRO, J. A. SEGOVIA-ZAVALA, A. MUÑOZ-BARBOSA & M. V. OROZCO-BORBÓN. 1997. The effect of vertical mixing on Primary Production in Bay of the Gulf of California. Estuarine, Coastal and Shelf Science 45: 135-148. [ Links ]
FIGUEIRAS, F. G., F. X. NIELL & C. MOURIÑO. 1986. Nutrientes y oxígeno en la ría de Pontevedra (NO de España). Investigaciones Pesqueras 50(1): 97-115. [ Links ]
GÁRATE-LIZÁRRAGA, I, M. L. HERNÁNDEZ-OROZCO, C. BAND-SCHMIDT & G. SERRANO-CASILLAS. 2001a. Red tides along the coasts of Baja California Sur, Mexico (1984 to 2001). Oceánides 16(2): 127-134. [ Links ]
GÁRATE LIZÁRRAGA, I., D. J. LÓPEZ CORTÉS, J. J. BUSTILLOS GUZMÁN, F. E. HERNÁNDEZ-SANDOVAL & I. MURILLO-MURILLO. 2001b. El Niño 1997-1998, impacto en la biomasa de dinoflagelados en bahía Concepción, Golfo de California. pp 153-162. In: E. ESCOBAR BRIONES, M, BONILLA, A, BADÁN, M. CABALLERO & A. WINCKELL (Eds.). Los efectos del fenómeno El Niño en México 1997-1998. Consejo Nacional de Ciencia y Tecnología, México. [ Links ]
GAXIOLA-CASTRO, G., J. GARCÍA-CORDOVA, J. E. VALDÉZ-HOLGUÍN & M. BOTELLO-RUVALCABA. 1995. Spatial distribution of chlorophyll a and primary productivity in relation to winter physical structure in the Gulf of California. Continental Shelf Research 15(9): 1043-1059. [ Links ]
GAXIOLA-CASTRO, G., S. ÁLVAREZ-BORREGO, M. F. LAVÍN, A. ZIRINO & S. NÁJERA-MARTÍNEZ. 1999. Spatial variability of the photosynthetic parameters and biomass of the Gulf of California. Journal of Plankton Research 21(2): 231-245. [ Links ]
GILMARTIN, M. & N. REVELANTE. 1978. The Phytoplankton characteristics of the Barrier Island Lagoons of the Gulf of California. Estuarine and Coastal Marine Science 7: 29-47. [ Links ]
GOERING, J. J. & J. D. CLINE. 1970. A note on denitrificaction in seawater. Limnology and Oceanography 15(2): 306-309. [ Links ]
GÓMEZ-GUTIÉRREZ, J., R. PALOMARES-GARCÍA, R. DE SILVA-DÁVILA, M. A. CARBALLIDO-CARRANZA & A. MARTÍNEZ-LÓPEZ. 1999. Copepod daily egg production and growth rates in Bahía Magdalena, México. Journal Plankton Research 21(12): 2227-2244. [ Links ]
HERNÁNDEZ-BECERRIL, D. 1983. Estructura del fitoplancton en la parte central y norte del Golfo de California (junio, 1982). Tesis de Maestría (Ecología marina), Centro de Investigación Científica y Educación Superior de Ensenada, México. 111 p. [ Links ]
HOLLIGAN, P. M., W. M. BALCH & C. M. YENTSCH. 1984. The significance of subsurface chlorophyll, nitrite and ammonium maxima in relation to nitrogen for phytoplankton growth in stratified waters of the Gulf of Maine. Journal of Marine Research 42: 1051-1073. [ Links ]
IGNATIADES, L., M. KARYDIS & P. VOUNATSOU. 1992. A possible method for evaluating oligotrophy and eutrophication based on nutrient concentration scales. Marine Pollution Bulletin 24: 238-243. [ Links ]
JUSTIC, D., N. N. RABALAIS & R. E. TURNER. 1995. Stoichiometric nutrient balance and origin of coastal eutrophication. Marine Pollution Bulletin 30(1): 41-46. [ Links ]
KARYDIS, M., L. IGNATIADES & N. MOSHOPOULOU. 1983. An index associated with nutrient eutrophication in the marine environment. Estuarine, Coastal and Shelf Science 16: 339-344. [ Links ]
KIØRBOE, T. 1993. Turbulence, Phytoplankton cell size, and the structure of pelagic food Webs. Advances in Marine Biology 29: 1-72. [ Links ]
LARA-LARA, J. R. & J. E. VALDÉZ-HOLGUÍN. 1988. Biomasa y productividad primaria del Golfo de California por fracciones de tamaños durante primavera de 1984. Ciencias Marinas 14(1): 1-14. [ Links ]
LAVÍN, M. F., E. PALACIOS-HERNÁNDEZ & C. CABRERA. 2003. Sea surface temperature anomalies in the Gulf of California. Geofísica Internacional 42(3): 363-375. [ Links ]
LECHUGA-DEVÉZE, C. H., M. L. MORQUECHO-ESCAMILLA, A. REYES-SALINAS & J. R. HERNÁNDEZ-ALFONSO. 2000. Environmental natural disturbance at Bahía Concepción Gulf of California. pp. 245-255. In: M. MUNAWAR, S. G. LAWRENCES, I. F. MUNAWAR & D. F. MALLEY (Eds.). Aquatic Ecosystems of México: Status and Scope Ecovision World Monographie Series. [ Links ]
LECHUGA-DEVÉZE, C. H., A. REYES-SALINAS & M. L. MORQUECHO-ESCAMILLA. 2001. Anoxia in coastal bay: case study of a seasonal event. Biologia Tropical 49(2): 525-534. [ Links ]
LÓPEZ-CORTÉS, D. J., J. BUSTILLOS-GUZMÁN, I. GÁRATE-LIZÁRRAGA, F. HERNÁNDEZ-SANDOVAL & I. MURILLO. 2003. Phytoplankton biomasses and hydrographic conditions during El Niño 1997-1998 in Bahía Concepción, Gulf of California. Geofísica Internacional 42(3): 495-504. [ Links ]
MARTÍNEZ-LÓPEZ, A. & I. GÁRATE-LIZÁRRAGA. 1997. Variación diurna de la materia orgánica particulada en una laguna costera del Golfo de California. Revista de Biología Tropical 45(3):1310-1317. [ Links ]
MCPHADEN, M. 1999. Genesis and evolution of the 1997-98 El Niño. Science 28: 950-954. [ Links ]
PALMER, D., D. J. FREDERICKS, C. SMITH & D. T. HEGGIE. 2000. Nutrients from sediments. AGSO Research. Newsletter 33: 1-4. [ Links ]
PETERSON, W. T. & D. C. BELLANTONI. 1987. Relationships between water-column stratification, phytoplankton cell size and copepod fecundity in Long Island Sound and off Central Chile. pp. 411-421 In: A. I. L. PAYNE, J. A. GULLAND & K. H. BRINK (Eds). The Benguela and Comparable Ecosystems. [ Links ] South African Journal Marine Science. [ Links ]
PINGREE, R. D., P. M. HOLLIGAN, G. T. MARDELL & R. N. HEAD. 1976. The influence of physical stability on spring, summer and autumn phytoplankton blooms in the Celtic Sea. Journal of the Marine Biological Association of the United Kingdom 56: 845-873. [ Links ]
PINGREE, R .D., L. MADDOCK & E. I. BUTLER. 1977. The influence of Biological activity and physical stability in determining the chemical distributions of inorganic Phosphate, Silicate and Nitrate. Journal of the Marine Biological Association of the United Kingdom 57: 1065-1073. [ Links ]
REVELANTE, N. & M. GILMARTIN. 1990. Vertical water column resource partitioning by a ciliated protozoan population under stratified conditions in the northern Adriatic. Journal of Plankton Research 12(1): 89-107. [ Links ]
ROBINSON, G. A. 1970. Continuous plankton records: variation in the seasonal cycle of phytoplankton in the North Atlantic. Bulletin of Marine Ecology 6: 333-345. [ Links ]
SCHAREK, R., V. SMETACEK, E. FAHRBACH, L. I. GORDON, G. ROHARDT & S. MOORE. 1994. The transition from winter to early spring in the eastern Weddell Sea, Antartica: Plankton biomass and composition in relation to hydrography and nutrients. Deep-Sea Research 41(8): 1231-1250. [ Links ]
SIMPSON, J. H., D. G. HUGHES & N. C. MORRIS. 1977. The relation of seasonal stratification to tidal mixing on the continental shelf. pp. 327-340. In: A. MARTIN (Ed.). A Voyage of Discovery, Deep-Sea Research (suppl. [ Links ]).
SIMPSON,J. H. & T. P. RIPPETH. 1993. The Clyde Sea: a model of the seasonal cycle of stratification and mixing. Estuarine, Coastal and Shelf Science 37:129-144. [ Links ]
SMITH, V. H., G. D. TILMAN & J. C. NEKOLA. 1999. Eutrophication: impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environmental Pollution 100: 179-196. [ Links ]
STAL, L.J., S. B. BEHRENS, M. VILLBRANDT, S. VAN BERGEIJK & F. KRUYNING. 1996. The biogeochemistry of two eutrophic marine lagoons and its effect on microphytobenthic communities. Hydrobiologia 329: 185-198. [ Links ]
STRICKLAND, J. D. H. & T. R. PARSONS. 1972. A practical handbook of seawater analysis. Fisheries Research Board of Canada Bulletin 310 pp. [ Links ]
THUNELL, R., E. TAPPA, C. PRIDE & E. KINCAID. 1999. Sea-surface temperature anomalies associated with the 1997-1998 El Niño recorded in the oxygen isotope composition of planktonic Foraminifera. Geology 843-846. [ Links ]
VALDÉZ-HOLGUÍN,J. E. & J. R. LARA-LARA. 1987. Productividad primaria en el Golfo de California, efectos del evento El Niño 1982-1983. Ciencias Marinas 13(2): 34-50. [ Links ]
VINER, A. B. 1985. Thermal stability and phytoplankton distribution. Hydrobiologia 125: 47-69. [ Links ]

