










Servicios Personalizados
Revista
Articulo
 Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.12 no.1 Ciudad de México jun. 2002
Article
Primary production in the southern Gulf of Mexico estimated from solar-stimulated natural fluorescence
Producción primaria en el sur del Golfo de México estimada a partir de fluorescencia natural
Raúl Aguirre Gómez
Instituto de Geografía. Universidad Nacional Autónoma de México. Ciudad Universitaria, Circuito Exterior. México, D. F, C. P. 04510. México. e-mail: raguirre@igiris.igeograf.unam.mx
Recibido: 14 de septiembre de 2001
Aceptado: 15 de abril de 2002
Abstract
This paper reports a study on the horizontal and vertical chlorophyll and primary production distributions. Chlorophyll a concentrations were calculated from natural fluorescence during the fall season of 1997 in the southern part of the Gulf of Mexico. It was found a nearly homogeneous distribution of chlorophyll (0.4 to 2.3 mg m−3) and primary production (1.2 to 6.7 mg C m−3 h−1) at oceanic areas, but conspicuously different values at coastal or sediment-laden zones (12.5 - 46.6 mg m−3 for chlorophyll and 34 - 120 mg C m−3 h−1 for production). The results obtained from solar stimulated natural fluorescence are similar to those calculated by other authors using different methods. This method provides reliable results and can be alternatively used.
Keywords: natural fluorescence, primary productivity, southern Gulf of Mexico.
Resumen
En este artículo se reporta un estudio sobre las distribuciones horizontal y vertical de la clorofila y la producción primaria. Las concentraciones de clorofila a fueron calculadas de la fluorescencia natural durante el otoño de 1997, en la región sur del Golfo de México. Se encontraron distribuciones homogéneas de clorofila (0.4 a 2.3 mg m−3) y de producción primaria (1.2 a 6.7 mg C m−3 h−1) en regiones oceánicas, pero diferentes niveles en las zonas costeras o con presencia de sedimentos (12.5 - 46.6 mg m−3 de clorofila y 34 - 120 mg C m−3 h−1 en la producción). Los resultados obtenidos mediante la fluorescencia natural estimulada por el sol son similares a los calculados por otros autores usando diferentes métodos. Los resultados muestran que este método proporciona resultados confiables y puede ser usado en forma alternativa.
Palabras clave: fluorescencia natural, producción primaria, sur del Golfo de México.
Introduction
The southern Gulf of Mexico is an important economical and ecological region due to fisheries and oil exploitation. Petroleum exploitation in this area has received attention with regard to potential impacts on the living resources of the area, hence the importance of a primary production analysis of the region.
Since the 1950's the biology of the Gulf of Mexico has been widely studied (Galstoff, 1954). Studies on the distribution of phytoplankton and primary productivity have been systematically carried out from the beginnings of 1970's (El-Sayed, 1972). Thus, colour gradients across the Loop Current have been related to phytoplankton concentrations with maximum values being separately reported by Ednoff (1974) and Maul (1974). Using satellite-derived information, Mueller-Karger et al. (1991) reported that primary production in the Gulf of Mexico can vary from mesotrophic conditions at the shelf break to oligotrophic conditions at the more oceanic sites. On the other hand, Signoret et al. (1998), using de 14C technique found relative high values of primary production at the Yucatan's continental shelf but lower values at oceanic areas of the southern Gulf of Mexico.
Estimation of primary productivity in the southern Gulf of Mexico utilising solar stimulated natural fluorescence has not been carried out so far, even though this technique has proven to be adequate for this task (e. g. Valdez-Holguín et al., 1995; García-Mendoza and Maske, 1996; Stegmann and Lewis, 1997). Natural fluorescence has been widely investigated as a mean for a rapid assessment of the distribution and biomass of oceanic phytoplankton and for estimating the rates of primary production (Kiefer et al., 1989; Chamberlain et al., 1990).
The aim of this study is to describe the spatial variability of chlorophyll and primary production estimated from solar-stimulated natural fluorescence in the southern Gulf of Mexico. So, it should provide a first insight on the distribution pattern of primary production at regional scale using this technique.
Theoretical background
Natural fluorescence in the ocean can be roughly defined as the solar stimulated emission of chlorophyll a in a narrow band centred at 683 nm. This emission is the resulting effect of light absorption by phytoplankton.
The relationship between natural fluorescence Ff (t, z) and the instantaneous rate of photosynthesis Fc (t, z) can be expressed through the following phenomenological equations:
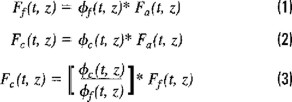
Where Fa (t, z) is the rate of light absorption and ϕf (t, z) are the quantum yield of photosynthesis and fluorescence, respectively. Equation (3) shows that the prediction of the photosynthetic rate from natural fluorescence explicitly depends on the ratio of the quantum yields. So, when this ratio is constant or can be predicted, reliable estimates of Fc(t, z) can be obtained. Unfortunately, this is not always the case and alternative and empirical solutions have been suggested (e. g. Chamberlain et al., 1990).
Kiefer et al., (1989) proposed that Fa(t,z) could be approximated by the following relationship:
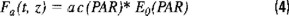
where ac stands for the absorption coefficient for phytoplankton (m−1) and E0 is the scalar irradiance (Eins m−2 s−1). The argument in both magnitudes indicates integration over the spectrum of visible light (400 - 700 nm), which corresponds to the photosynthetically available radiation (PAR).
Hence, the flux of natural fluorescence can be written as:
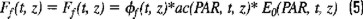
Gordon (1979) defined the product ϕf(t,z)*ac(PAR, t, z) as the natural fluorescence coefficient ψf(t, z). Then, the relationship between chlorophyll a concentration and natural fluorescence can be expressed in terms of ψf, ϕf, and the mean chlorophyll a (Chl) specific absorption coefficient, °ac(PAR,t,z), for the phytoplankton, as follows (Gordon, 1979; Kishino et al., 1984; Topliss, 1985; Chamberlain et al., 1990):

Likewise, daily rates of gross photosynthesis, Fc(z, 24h) can be estimated at depth through ψf, EO(PAR, z, 24 h) and the daily mean value for the quantum yields ratio (ϕc:ϕf), accordingly to the following expression:
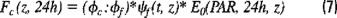
However, in this study, the daily rates of photosynthesis were evaluated during the daylight period instead of 24 hrs.
Study area
The study area is located at the southern region of the Gulf of Mexico, between 18°45' and 22°15' N, and 89°45' and 92°45' W. Two main rivers discharge fresh water into the area: the Coatzacoalcos and the Grijalva-Usumacinta. These rivers are the main sediment sources for the inshore and offshore areas and they represent approximately one-third of all the fluvial discharge in Mexico (Carranza-Edwards et al., 1993). The Terminos lagoon is located in the southern portion of the region. Circulation in the region is cyclonic, driven by an eddy formed at the east, with a westward displacement (Molinari et al., 1978). The annual precipitation reaches 4.5 m and there is an average of 160 cloudy days per year.
The region covers a number of physiographic areas that include Campeche Bank and Campeche Bay, according to Antoine's classification (Antoine, 1972). Figure 1 shows the main features of the region.
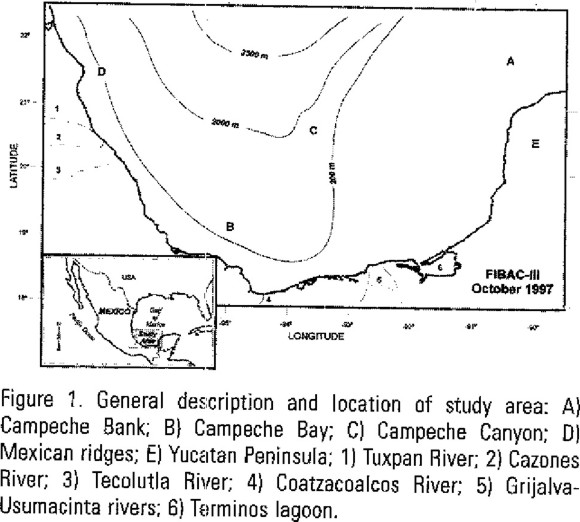
Campeche Bank. It is an extensive and almost flat carbonate region bordered to the west by the Tabasco-Campeche Basin and to the east by the Yucatan straits. This region is influenced by upwellings along the coast of Yucatan peninsula. There is a strong current derived from the Gulf Stream entering through the strait. It is a shallow area with depths ranging between 20 - 200 meters. On the western side of Campeche Bank there is a submarine canyon marking the end of the carbonated area of Campeche Bank and the beginning of the continental shelf. It is known as the Campeche Canyon. Campeche Bay. This region is located in the southern portion of the Gulf of Mexico, limited by parallel 21° N and by the coasts of the states of Veracruz, Tabasco, Campeche and Yucatan to the east, south and west, respectively. It is the seaward extension of the Tabasco-Campeche Basin. The bay, opened toward the Gulf, is bounded on the east by the Campeche Bank, on the south and southwest by the Sierra Madre Oriental. The bordering coastal plain is low, poorly drained and swampy. Circulation within the bay is influenced by the current coming from the Campeche Bank, which is produced by the Yucatan current and Yucatan upwelling (Hernández-Téllez et al., 1993). The rainy season, from May to October (García, 1988), produces the input of fresh water into the bay through the Grijalva-Usumacinta rivers. Fresh water input diminishes the salinity and modifies seawater temperature, creating a coastal front (Monreal-Gómez et al., 1992). It is a deeper oceanic region (2000 - 3500 meters).
Methods and materials
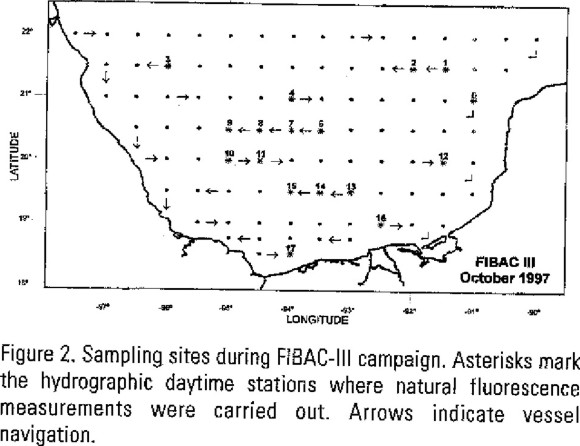
The study was conducted with measurements carried out on board the R/V Justo Sierra (UNAM) from October 3 to 13, 1997. Sampling stations were evenly distributed in a 0.5° × 0.5° latitude/longitude grid (figure 2). Navigation time from one station to other was about 3 hours long, and the sampling time was around 1 hour at each station. Thus, a total of 91 stations were sampled both during daytime and night-time hours during the cruise. For the purposes of this study, only daytime stations are considered since they are suitable for measuring solar stimulated natural fluorescence. However, not all of these stations were sampled because of an instrumental electronic failure at the beginning of the cruise and of the onset of adverse weather conditions around October 14Th. Therefore, the study was carried out at 17 stations distributed along the southern Gulf of Mexico. Five stations were located at the Campeche Bank (1, 2, 5, 12, 16); nine stations were located at the Campeche Bay (3, 4, 6, 7, 8 9, 10, 11, 15); two stations were located close to Campeche Canyon (13, 14); one coastal station (17) was located between the discharging areas of the Coatzacoalcos and Grijalva-Usumacinta rivers.
Mean pigment concentration at the euphotic depth was estimated from the model proposed by Morel and Berthon (1989). For each site, the water column was characterised and the variation of primary production was described. Day length period was of 11.40 h. and was calculated as described by Kirk (1983).
Natural Fluorescente
Vertical profiles of upwelling radiance at the fluorescence wavelength (Lu683), scalar PAR irradiance, water temperature and pressure (depth) were recorded with a Profiling Natural Fluorometer (PNF-300, Biospherical Instruments). It is a small, lightweight instrument attached to a metal frame. The instrument was lowered into the surface from the stern of the ship by hand, avoiding ship shadowing which may introduce errors up to 30% (Gordon, 1985). The PFN-300 was equipped with 200 m of conducting cable that provided real-time profiles on a personal computer onhoard ship. Vertical profiles of the total diffuse attenuation coefficient, kPAR (m−1), were obtained from 20-point smoothed PAR profiles (Chamberlain et al., 1990). Instruments and sensors were calibrated prior the campaign.
Primary Productivity
Estimates of primary productivity and biomass were obtained from natural fluorescence measurements of phyto-plankon. It is assumed that all of the natural fluorescence (radiance centred at 683 nm) measured with the profiler is due to chlorophyll a. In order to estimate productivity using this method, the ratio of natural fluorescence is related to the photosynthetic ratio by means of an empirical equation proposed by Chamberlain et al., (1990), which is related to equation (3). They found that the ratio of the quantum yields, (ϕc:ϕf), rather than being constant or predictable, decreases with increasing light intensity. This empirical equation can be expressed as a function of time and depth (t, z), as follows:
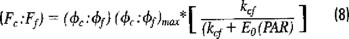
where (ϕc:ϕf)max is an empirical constant representing the maximum value of the ratio of quantum yields and kcf is another empirical constant which stands for the value of irradiance when the ratio is equal to half its maximum value. Quantum yields ratios were calculated from equation (3) by plotting Fc vs. Ff. The slope of the curve is the quantum yields ratio (ϕc:ϕf). Thus, the quantum yield ratio and therefore the ratio of carbon assimilation to fluorescence emission is a function of irradiance. In Chamberlain et al.'s model (1990), the "best-fit" value for the ratio between the quantum efficiencies of photosynthesis and fluorescence was 2.3 assimilated carbon atoms per photon emitted, and an irradiance value of 133 mE m−2 s−1. Hence, solving for the photosynthesis term Fc(t, z), the following equation is obtained:
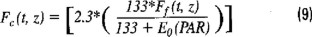
Thus, this equation relates photosynthetic rate to natural fluorescence and irradiance.
Results
Natural fluorescence
The relationship derived by Kiefer et al. (1989) was used to calculate the volume natural fluorescence emission (mol quanta m−3s−1) from the Lu683 signal (mol quanta m−2s−1str−1) according to the following equation:
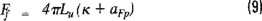
where κ is the diffuse attenuation coefficient for downward irradiance, aFp is the attenuation coefficient for the fluorescence emission on the path of the sensor, and 4π is a geometric constant to convert the radiance to volume emission and indicates the assumption of isotropy.
Distribution Patterns of chlorophyll a and primary productivity.
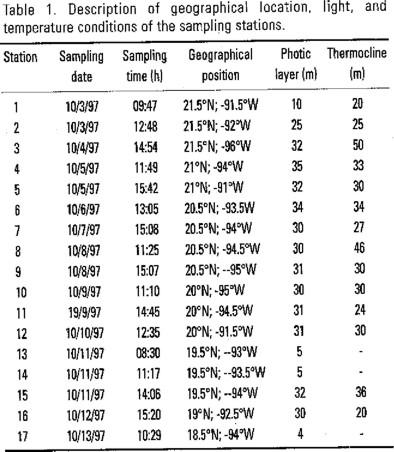
The stations sampled in this cruise virtually cover the southern part of the Gulf of Mexico over a wide geographic range. Table 1 shows representative spatial and vertical structure of temperature, light conditions, chlorophyll a and primary production.
Spatial variability of Chlorophyll a and productivity rates.
The distribution of chlorophyll a in surface waters, as measured by fluorescence, shows variations due to geographical characteristics. Relatively high chlorophyll a concentrations ranging from 12.5 to 46.6 mg m−3 were found at the coastal location (17), at areas under Yucatan's upwelling influence (1) and close to Campeche Canyon (13, 14). The standing crop values over the remaining stations in the region studied were, in general, low. Chlorophyll a values range in these stations from 0.6 to 2.3 mg m−3, with a mean value of 1.1 (σ = 0.51) mg m−3.
The mean total primary productivity at oceanic locations was 2.5 (σ= 1.6) mg C m−3 h−1 and varied from 1.2 to 6.7 mg C m−3 h−1. Primary productivity estimates at stations influenced by topographical features ranged from 34 to 120 mg C m−3 h−1. The highest rates were found at stations near Campeche Canyon.
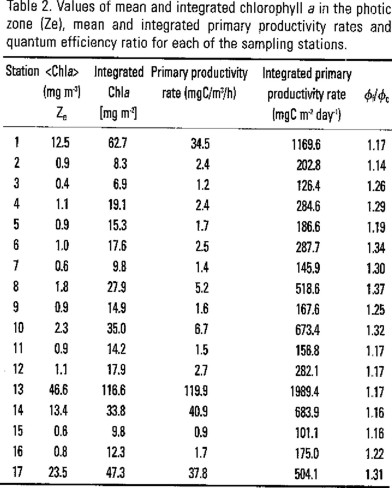
Integrated values of chlorophyll a and productivity rates.
Integrated euphotic zone chlorophyll a contents showed moderately high values (> 9.5 and < 20 mg m−2) in most of the oceanic sampling areas located at Campeche Bay, except for coastal regions including station 1 within Yucatan's upwelling influence and stations 13 and 14 located at Campeche Canyon that reached values between 33.8 and 116.6 mg m−2. Stations located at Campeche Bank also showed moderately high values, which seems to be characteristic of the zone (Sackett, 1972). The mean of depth-integrated chlorophyll a at oceanic areas was of 19.3 (σ = 11.6) mg m−2, with no appreciable distribution pattern.
Daily-integrated primary productivity rates ranged from 101 to 1990 mg C m−2 day−1. Lowest rate values were found at oceanic areas with a mean value of 192.4 (σ = 65.4) mg C m−2 day−1, excluding stations 8 and 10 which presented relatively high productivity rates of 518.6 and 673.4 mg C m−2 day−1. Stations influenced by topographical features had the highest productivity rates ranging from 504.1 to 1989.4 mg C m−2 day−1.
Vertical Distributions.
Based on differences due to geographical location alone, variations in hydrographic and biological characteristics between stations were expected. Such differences are clearly found in the temperature vertical profiles. Thus water temperatures at Campeche Bank were rather homogeneous in the upper 20 to 30 m, whilst those at Campeche Bank showed a wider thermocline range from 27 to 50 m. Shallow stations located at the coast and near Campeche Canyon showed no thermocline. Figure 3 exemplifies these general trends.
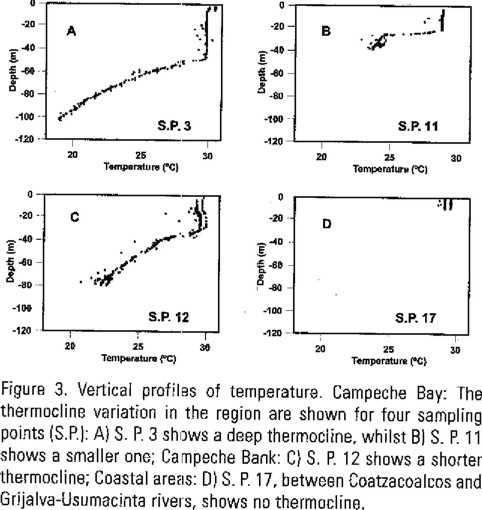
Photic zones in the study area were found to be between a 25 to 35 m range for nearly all stations. Four stations were out of this range: a) Station 1, located at Campeche Bank, with a photic zone of 10 m, presumably influenced by the Yucatan upwelling current; b) stations 13, 14, located near Campeche Canyon, with 5 m photic zone, mainly affected by sediment transport occurring at the interface and c) station 17, located near the Tabasco-Campeche coast, with a 4 m photic zone, optically affected by the discharges of Coatzacoalcos river and, to a lesser extent, by Grijalva-Usumacinta rivers.
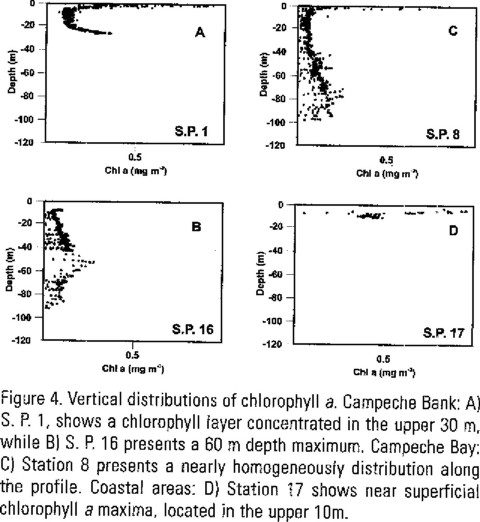
Vertical distributions of chlorophyll a showed different patterns. Stations 3, 4, 5, 8, 12, and 15 were nearly homogeneously mixed with depth. Stations 1, 2, 6, 7, 9, 10, 11, and 16 were characterised by a chlorophyll rich layer concentrated in the upper 20 - 40 m, excluding station 16 that presented a 60 m depth maximum. Stations 13, 14, and 17 showed near superficial chlorophyll a maxima, located in the upper 10 m. Figure 4 shows examples of each one of these distribution patterns.
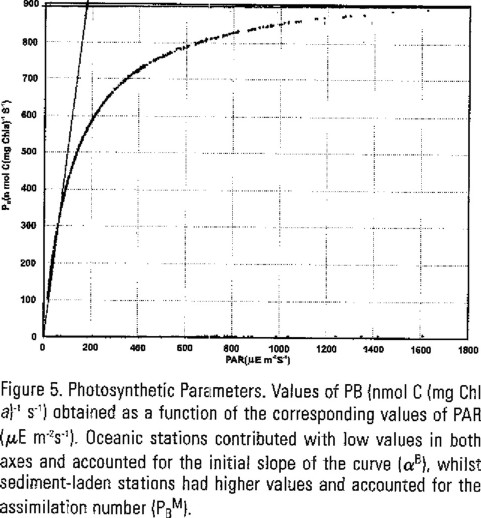
At the 17 stations the phytoplanktonic biomass was similar as reflected by the chlorophyll a concentration, which ranged from 0.4 to 2.3 mg m−3, within the oceanic stations and from 12.5 to 46.6 mg m−3 at the sediment-laden stations. To compare both cases, we normalised the production parameters to biomass, PB, which is the ratio of carbon fixed to the concentration of chlorophyll a, expressed as nmol C (mg Chl a)−1s−1. The values of PB obtained as a function of the corresponding values of PAR (µE m−2s−1) are shown in figure 5. The curve obtained is related to conventional P versus I curves, but it differs from them since irradiance is not constant at each depth. In this curve, which includes all of the stations, oceanic stations' values were lower and accounted for the linear interval of the curve (αB), whilst those for the sediment-laden stations laid within the assimilation number PB. The values found for these parameters are: αB = 4.5 nmol C (mg Chl a)−1 s−1 µEins m2 and PB = 900 nmol C (mg Chl a)−1 s−1. The nearly "perfect" shape of the curve suggests either that primary productivity estimates obtained from the PNF-300 instrument follow a pre-established model such as those proposed by Plan et al., (1980) or Geider and Osborne (1992), or it is rather a consequence of equation (9), since PB depends solely on the measured fluorescence. It must be noted that photoinhibition did not occur at these values of PAR.
Evaluating the relationship between primary production and natural fluorescence, a mean value for the ratio of quantum yields (ϕc/σf) of 1.2 (σ= 0.07) was found, suggesting that around 12 photons fluoresced per 10 carbon atoms fixed. This result is close to the value of 1.5 reported by Chamberlain et al., (1990). Values of quantum yield ratios of each station are included in Table 2.
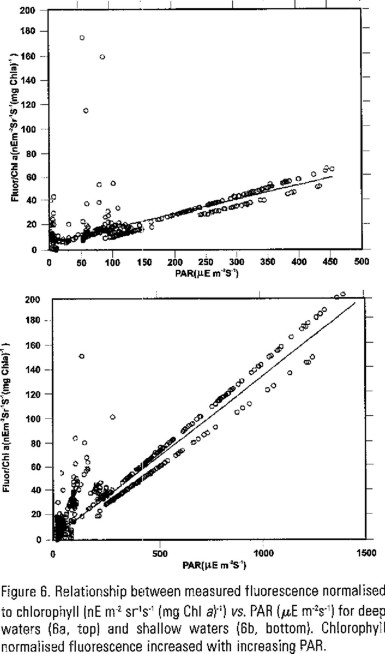
The relationship between measured fluorescence normalised to chlorophyll vs. PAR is shown in figure 6, which exemplifies deep (figure 6a) and shallow waters (figure 6b). Fluorescence, at the stations exemplified here, increased with increasing PAR and did not decline at higher irradiance intensities.
Discussion
In this paper we presented results from a cruise to the southern Gulf of Mexico in October 1997 where both biological and optical measurements were carried out. Cruise stations covered a number of geological and geographic environments, which was reflected in the temperature, chlorophyll a distributions and primary productivity rates. Thus, two distinct regions were detected, firstly, a region comprehending shallow coastal areas, upwelling and zones influenced by submarine topography, and secondly, deeper areas with oceanic characteristics. These regions can be clearly appreciated in satellite imagery.
In this study we assumed that primary productivity estimations made with natural fluorescence are consistent with in situ data measured with traditional techniques such as 14C after encouraging results reported by other researchers (Chamberlain et al., 1990; Valdez-Holguín et al., 1995; García-Mendoza and Maske, 1996; Stegmann and Lewis, 1997) and, therefore, it can be used on its own, as a reliable method for this task.
In the coastal, and topographically influenced regions, several physical processes, such as mixing, upwelling, or advection, are responsible for the generation of fronts (Monreal-Gómez and Salas de León, 1990). These fronts are nutrient-rich environments; therefore the productivity rates here were higher. However, these magnitudes must be considered cautiously due to the amount of terrigenous suspended sediment that may be present along with phytoplankton as reported by Carranza-Edwards et al. (1993). Due to the morphology of the basin, the different currents, upwelling and river discharge, the water chemistry (nutrients) and physical environment (light availability, vertical mixing, depth of the photic zone) the system is quite complex. Therefore, this is very likely to have an impact on the calculation of quantum efficiencies for fluorescence.
The mean of depth-integrated chlorophyll a stock in the area was lower than those reported in other Mexican waters such as the Gulf of California (Valdez-Holguin et al., 1995) or the Gulf of Tehuantepec (Robles-Jarero and Lara-Lara, 1993). The main reason accounting for this difference is that these two regions are well known for their high rates of primary productivity. However, the value of the mean is similar to that reported by El-Sayed (1972) for the Gulf of Mexico, who found mean values for primary production and chlorophyll a concentrations, over a seven year period (1964-1971), of 0.35 mg C m−3 hr−1 (6.63 mg C m−2 hr−1) and 0.20 mg m−3 (12.42 mg m−2), respectively. The values reported in this paper are also in the range found by Signoret et al., (1998) who reported relatively high values of primary production (0.84 - 2.11 g C m−2 day−1) and of integrated chlorophyll a (13.28 - 36.85 mg m−2) near the Yucatan's continental shelf, whilst lower values were found at the southern and western oceanic zones of the Gulf of Mexico.
The integrated primary productivity estimated from solar-stimulated natural fluorescence shows variations that are possibly associated, on one hand, to the time of the day stations were sampled and, consequently, to in situ irradiance variations (Kirk, 1983) and, on the other hand, to the extrapolation of productivity for the day length.
Natural fluorescence measurements can be used as an alternative and reliable approach for estimating primary production, although it will not do any harm to eventually combine in situ primary productivity data, measured with traditional techniques, such as 14C, and natural fluorescence profiles. This combination will support the results and can be important for calibration purposes.
Acknowledgements
I wish to thank Dr. T. Platt and Dr. S. Sathyendranath for commenting on the manuscript. I thank also Dr. D. Salas and Dr. A. Monreal for their kind invitation to participate in FIBAC-III cruise. Thanks are also given to Dr. M. Signoret for allowing the use of the PNF-300 instrument. I am grateful to the Government of Canada, through the International Council for Canadian Studies (ICCS), for the financial support during my stay in Canada at the Bedford Institute of Oceanography (BIO), Biological Oceanography Division. Finally, I wish to acknowledge the crew of the R/V Justo Sierra and the staff of the BIO for all the help provided.
References
ANTOINE, J. W., 1972. Structure of the Gulf of Mexico. pp. 1-34. En: R. REZAK & V. J. HENRY (Eds.). Contributions on the Geological and Geophysical Oceanography of the Gulf of Mexico, Texas A & M University Oceanographic Studies 3. [ Links ]
CARRANZA-EDWARDS, A., ROSALES-HOZ, L. and A. MONREAL-GÓMEZ, 1993. Suspended sediments in the southeastern Gulf of Mexico. Marine Geology 12: 257-269. [ Links ]
CHAMBERLAIN, W. S., BOOTH, C. R., KIEFER, D. A., MORROW, J. H. and R. C. MURPHY, 1990. Evidence for a simple relationship between natural fluorescence. photosynthesis and chlorophyll in the sea. Deep-Sea Research 37: 951-973 [ Links ]
EDNOFF, M. E., 1974. Surface phytoplankton communities and their relationship to the loop current in the Gulf of Mexico. M. S. Thesis, Florida State University, Tallahassee. 187 p. [ Links ]
EL - SAYEO, S. Z., 1972. Primary productivity and standing crop of phytoplankton, pp. 8-13. En: V. C. BUSHNELL (ed.). Chemistry primary productivity, and benthic marine algae of the Gulf of Mexico, Serial atlas of the marine environment, folio 22. American Geogrephical Society 8. [ Links ]
GALSTOFF, P. S., 1954. Gulf of Mexico - its origin, waters, and marine life. Fisheries Bulletin 89, Fish and Wildlife Service, 604 p. [ Links ]
GARCÍA, E., 1988. Modificaciones al sistema de clasificación climática de Köeppen. Ed. Ofsett Larios. 264 p. [ Links ]
GARCÍA-MENDOZA, E. and H. MASKE, 1996. The relationship of solar-stimulated natural fluorescence and primary productivity in Mexican Pacific waters. Limnology and Oceanography 41(18): 1697-1710. [ Links ]
GEIDER, R. J. and B. A. OSBORNE, 1992. Algal photosynthesis. Chapman and Hall. 256 p. [ Links ]
GORDON, H., 1979. Diffuse reflectance of the ocean: the theory of its augmentation by chlorophyll a fluorescence at 685 nm. Applied Optics 18: 1161-1166. [ Links ]
GORDON, H. R., 1985. Ship perturbation of irradiance measurernents at sea, 1: Monte Carlo simulations. Applied Optics 24(23): 4172-4182. [ Links ]
HERNÁNDEZ-TÉLLEZ, J., J. ALDECO and D. A. SALAS DE LEÓN, 1993. Cooling and Heating due to latent and sensible heat over the Yucatan continental shelf. Atmósfera 6(4): 223-233. [ Links ]
KIEFER, D. A., W. S. CHAMBERLAIN and C. R. BOOTH, 1989. Natural Fluorescence of chlorophyll a: Relationship to photosynthesis and chlorophyll concentration in the western South Pacific gyre. Limnology and Oceanography 34(5): 868-881. [ Links ]
KIRK, J. T. O., 1983. Light and photosynthesis in aquatic ecosystems. Cambridge University Press. 401 p. [ Links ]
KISHINO, M., S. SUGIHARA and N. OKAMI, 1984. Influence of fluorescence of chlorophyll a on undewater upward irradiance spectrum. La Mer 22: 224-232. [ Links ]
MAUL, G. A., 1974. Relationship between ERTS radiances and gradients across oceanic fronts, pp. 1279. En: Third Earth Res. Tech. Satellite Symp. V. 1, NASA. [ Links ]
MOLINARI, R. L., J. F. FESTA and D. W. BEHRINGER, 1978. The circulation in the Gulf of Mexico derived from estimated dynamic height fields. Journal of Physical Oceanography 8: 987-996. [ Links ]
MONREAL-GÓMEZ, M. A. and D. A. SALAS DE LEÓN, 1990. Simulación de la circulación de la Bahía de Campeche. Geofísica Internacional 29(2): 101-111. [ Links ]
MONREAL-GOMEZ, M. A., D. A. SALAS DE LEÓN, A. R. PADILLA-PILOTZE and M. A. ALATORRE-MENDIETA, 1992. Hydrography and estimation of density currents in the southern part of the Bay of Campeche, Mexico. Ciencias Marinas 18(4): 115-133. [ Links ]
MOREL, A. and J. F. BERTHON, 1989. Surface pigments, algal biomass profiles, and potential production of the euphotic layer: Relationships reinvestigated in view of remote-sensing applications. Limnology and Ocaanography 34(8): 1545-1562. [ Links ]
MUELLER-KARGER, F. E., J. J. WALSH, R. H. EVANS and M. B. MEYERS, 1991. On the seasonal phytoplankton concentration and sea surface temperature cycles of the Gulf of Mexico as determined by satellites. Journal of Geophysical Research 96(C7): 12645-12665. [ Links ]
PLATT, T., C. L. GALLEGOS and W. G. HARRISON, 1980. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. Journal of Marine Research 38: 687-701. [ Links ]
ROBLES - JARERO, E. G. and J. R. LARA - LARA, 1993. Phytoplankton biomass and primary productivity by size classes in the Gulf of Tehuantepec, México. Journal of Plankton Research 15(12): 1341-1358. [ Links ]
SACKETT, W. M., 1972. Chemistry of the Gulf of Mexico, pp. 1-2. En: V. C. BUSHNELL (ed.), Chemistry, primary productivity, and benthic marine algae of the Gulf of Mexico, Serial atlas of the marine environment, folio 22. American Geographical Society 8. [ Links ]
SIGNORET, M., C. BULIT y R. PÉREZ, 1998. Patrones de distribución de clorofila a y producción primaria en aguas del Golfo de México y del Mar Caribe. Hidrobiológica 8(2): 81-88. [ Links ]
STEGMANN, P. M. and M. R. LEWIS, 1997. Shipboard measurements of phytoplankton production and solar-stimulated fluorescence rates in the Northwest Atlantic. Continental Shelf Research 17:743-760. [ Links ]
TOPLISS, B. J., 1985. Opticel measurements in the Sargasso Sea: solar stimulated chlorophyll fluorescence. Oceanologica Acta 8:263-270. [ Links ]
VALDEZ-HOLGUÍN, J. E., G. GAXIOLA-CASTRO and R. CERVANTES-DUARTE, 1995. Primary productivity in the Gulf of California, calculated from the relationship between superficial irradiance and chlorophyll in the euphotic zone. Ciencias Marinas 21(3):311-329. [ Links ]

