










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkHidrobiológica
versión impresa ISSN 0188-8897
Hidrobiológica vol.12 no.1 Ciudad de México jun. 2002
Artículo
Taxonomía y distribución de Hildenbrandia Nardo (Hildenbrandiales, Rhodophyta) de agua dulce en la región central de México
Taxonomy and distribution of freshwater Hildebrandia Nardo (Hildenbrandiales, Rhodophyta) in central Mexico
Javier Carmona-Jiménez, Gustavo Montejano y Jorge González-González
Laboratorio de Ficología, Facultad de Ciencias, UNAM. Apdo. Postal 70-620. Ciudad Universitaria, Coyoacán, 04510, México, D.F. Tel: (52) 56 22 48 37. Fax: (52) 56 22 48 28. E-mail: jcj@hp.fciencias.unam.mx
Recibido: 16 de julio de 2001
Aceptado: 10 de diciembre de 2001
Resumen
En este trabajo se presentan los resultados del análisis taxonómico, descripción ambiental y la distribución de 23 poblaciones del género Hildenbrandia recolectadas en 14 localidades de México y una del centro de Europa. Se reconocieron dos grupos, el primero formado por todas las poblaciones mexicanas que se caracterizaron por el diámetro celular pequeño (promedio 4.1µm) y fueron consideradas como una sola especie (H. angolensis). La segunda especie se diferenció por el diámetro celular mayor (promedio 6.4 µm) y correspondió a la población europea (H. rivularis). Se registraron propágulos vegetativos en 10 poblaciones mexicanas. Hildenbrandia angolensis se presentó en ríos y manantiales con temperatura elevada (23-27 "C), neutral a alcalina (pH 7.0-8.0), alto contenido de iones (conductividad específica 700-1520 µS cm−1), velocidad de corriente de moderada a alta, alcalinidad alta (400-554 mg l1) y sitios sombreados con poca profundidad (típicamente <70 cm); estos datos fueron similares a los registros previos. Hildenbrandia angolensis es descrita por primera vez en la cuenca alta del río Balsas y cuenca baja del río Papaloapan, en áreas con clima tropical a subtropical.
Palabras clave: Dstribución, Hildenbrandiales, Hildenbrandia, México, Rhodophyta, ríos, taxonomía.
Abstract
This work includes a taxonomic analysis, habitat descriptions and distribution of 23 populations of the genus Hildenbrandia, collected in 14 sities in Mexico and one in Central Europe. Twenty two populations from Mexico were considered as belonging to a single species (H. angolensis), characterized by having small cell diameter (mean 4.1 µm). The second species (H. rivularis), represented by 1 population from central Europe was characterized by the large cell diameter (mean 6.4 µm). The presence of propagules was observed in 10 Mexican populations. Hildenbrandia angolensis tended to occur in warm (temperature 23-27 °C), neutral to alkaline (pH 7.0-8.0), high ion content (specific conductance (700-1520 µS cm−1), moderate to fast flowing, high alkalinity (400-554 mg l−1) and shallow waters (depth typically <70 cm); these data are essentially similar to previous records. Hildenbrandia angolensis is described for the first time from the higher Balsas and lower Papaloapan basins, both occurring mostly in areas of tropical or subtropical rainforests.
Key words: Distritiution, Hildenbrandiales, Hildenbrandia, Mexico, Rhodophyta, stream, taxonomy.
Introducción
El género Hildenbrandia es un habitante común de ambientes marinos y dulceacuícolas de zonas templadas o tropicales del mundo (Sheath et al., 1993; Necchi et al., 1999; Sherwood y Sheath, 2000b). Se caracteriza por el desarrollo de un talo costroso de construcción heterótrica, compuesto por una capa de células postradas y una porción de filamentos erectos que forman el pseudoparénquima. La delimitación de especies está basada en la altura del talo, el tamaño celular, la dimensión y tipo de zonación del tetrasporangio. Sin embargo, en las especies dulceacuícolas la reproducción sexual se desconoce y la multiplicación vegetativa se realiza por la formación de propágulos, que son agregados celulares de forma esférica dispuestos en cavidades de la capa polistromática (Starmach, 1969; Seto, 1977, Sherwood y Sheath, 2000b). Para agua dulce se han descrito dos especies. La primera con promedio de largo celular 11.3 µm y diámetro 8.2 µm como H. rivularis (Liebmann) J. Agardh, ampliamente distribuída para Europa (Israelson, 1942; Starmach, 1969, 1977; Sherwood y Sheath, 2000a), Asia (Skuja, 1938; Jao, 1941) y Norteamérica (Wolle, 1887; Flint, 1955) e H. angolensis Welwitsch ex West y West con promedios de largo celular 6.4 µm y diámetro 4.9 µm, descrita para Africa (West y West, 1897), Norteamerica (Sheath et al., 1993; Sherwood y Sheath, 1999) y Sudamérica (Necchi, 1987; Necchi et al., 1999). Existe una tercera especie, H. ramanaginaii Khan referida para la India (Khan, 1974), sin embarso, Sherwood y Sheath (2000b) consideran que la descripción de esta especie no corresponde con las características del género Hildenbrandia y el material tipo no está accesible para su examen. Recientemente Sherwood y Sheath (1999, 2000a) analizaron la secuencia de nucleótidos en fragmentos de los genes rbcL y 18S rRNA y demostraron que las especies de agua dulce son monofiléticas con muy poca diferenciación entre los grupos.
Diversas poblaciones de Hildenbrandia han sido recolectadas en la región central de México. Carmona y Montejano (1993) registraron las condiciones ambientales de H. rivularis en tres manantiales calcáreos de San Luis Potosí. Sheath et al. (1993) describieron las características morfométricas y ambientales de H. angolensis en un sitio no definido de la región central de México. Montejano et al. (2000) incluyeron H. angolensis en la lista de especies presentes en ríos de la región de la Huasteca. No obstante, aún falta el análisis de su taxonomía y distribución. En este trabajo se presentan los resultados en la evaluación de los caracteres taxonómicos, la descripción ambiental y la distribución del género Hildenbrandia en adición a los registros previos de la región central de México.
Material y método
Se estudiaron un total de 23 poblaciones con Hildenbrandia recolectadas en 14 localidades entre los 18° - 22° LN y los 95° - 99° LO) y altitudes de 60 a 1,121 m. (Fig. 1). Con el objeto de comparar la morfología de nuestras poblaciones se incluyó una población de Europa central (río Úslava, cerca de Pilsen, República Checa, coll. F. Lederer, 1.12.1995. FCME3939; temperatura del agua 15°C). En el campo se recolectaron los crecimientos algales visibles, tomando el fragmento de roca con cincel y martillo. Cada crecimiento visible quedó referido a un número de muestra el cual incluyó el registro de los factores ambientales en los que se encontró. Las muestras se preservaron en formalina al 4% y fueron depositadas en el herbario FCME (Holmgren et al.,1990) con las siglas "PA", "PAP" y "BALE" que corresponden a la cuencas de los rías Pánuco, Papaloapan y Balsas respectivamente. Los valores fisicoquímicos del agua se registraron con un analizador electroquímico marca Jenwey 3405 (pH ± 0.02; conductividad ± 0.5 %; temperatura ± 0.5). La insolación la empleamos como una estimación de la cantidad de luz que llega al punto de colecta y es expresada como el porcentaje del horizonte que se observa desde el punto de colecta (Johansson, 1982; Carmona y Montejano, 1993). El registro de la velocidad de corriente se consideró empleando las cinco categorías empleadas por Johansson (1982). La valoración de alcalinidad se cuantificó mediante el método indicador de alcalinidad total (Taras et al., 1971).
Se estudiaron todos los caracteres taxonómicos previamente considerados en este género (Wolle, 1887; West y West, 1897; Jao, 1941; Skuja, 1938, Israelson, 1942; Starmach, 1952, 1969, 1977; Nichols, 1965; Bourrelly, 1970; Seto, 1977; Necchi, 1987; Sheath et al., 1993; Sherwood y Sheath, 1999, 2000b). Cada población quedó definida por la observación de 3 a 4 zonas de la muestra. Las mediciones fueron hechas en réplicas de 20 y quedaron representadas como un intervalo que contiene la medida máxima, mínima, media aritmética y desviación estándar. El número de réplica fue determinado por la ecuación: 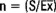 , donde S = desviación estándar,
, donde S = desviación estándar, 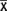 = promedio y E = error estándar predeterminado (en este caso 0.05) (Southwood, 1978). Cada espécimen fue observado, medido y fotografiado con un microscopio Reichert con sistema de iluminación contraste interferencial y sistema de fotomicrografía. El grosor de la capa polistromática, número de células en el filamento, largo y diámetro celular se registraron en el centro del crecimiento en vista transversal.
= promedio y E = error estándar predeterminado (en este caso 0.05) (Southwood, 1978). Cada espécimen fue observado, medido y fotografiado con un microscopio Reichert con sistema de iluminación contraste interferencial y sistema de fotomicrografía. El grosor de la capa polistromática, número de células en el filamento, largo y diámetro celular se registraron en el centro del crecimiento en vista transversal.
Resultados
Análisis morfológico
Se reconocieron dos grupos de poblaciones de Hildenbrandia en la región central de México y República Checa (Tabla 1, Figs 2-7), El primer grupo de todas las poblaciones de la región central de México correspondió a la circunscripción de H. angolensis descrita por Sheath et al. (1993). Estuvo caracterizada por el promedio del largo y diámetro celular relativamente pequeños (5.6 x 4.1 µm) (Tabla 2). Dos características morfológicas importantes fueron registradas en algunas poblaciones mexicanas. La primera fue el patrón de crecimiento donde se observó el desarrollo del talo pseudoparenquimatoso monostromático, con organización filamentosa y numerosas ramificaciones (Fig. 2); en vista superior la forma de las células fue rectangular y el crecimiento del talo fue irregular (Fig. 3) o radial (Fig. 4). Posteriormente, se reconoció el talo pseudoparenquimatoso polistromático, filamentoso, con ramificaciones escasas y 3 a 14 células por filamento; en vista transversal se presentaron filamentos con células cuadradas (Fig. 6) y en vista superior y centro del talo las células se observaron poligonales. En el márgen del talo polistromático se observaron células rectangulares similares a las registradas en el talo monostromático (Fig. 5). La segunda característica fue la presencia de propágulos vegetativos en 10 poblaciones (Tabla 1). Estos se observaron como agregados celulares elipsoidales, de dimensión variable, solitarios o en grupos, que se diferenciaron en cavidades de la capa polistromática. En todas las poblaciones con propágulos se registraron espacios vacíos en la superficie de la costra como resultado de la liberación de las estructuras de reproducción. (Fig. 7).
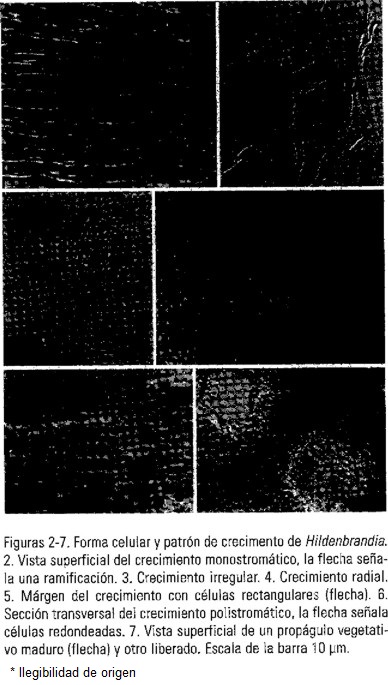
El segundo grupo correspondió a la población europea (Tabla 1) y se determinó como H. rivularis de acuerdo con las características descritas por Sheath et al. (1993) (Tabla 2). Estuvo caracterizada por el promedio del largo y diámetro celular relativamente grande (6.4 x 6.3 µm). Se observaron las capas monostromática y polistromática, las cuales fueron similares a lo descrito por Starmach (1952) y Nichols (1965) para esta especie.
Hildenbrandia angolensis Welwitsch ex West et West. J. Bot. 35:1-7, 1897.
LOC. SINTIPO: Golungo Alto, río cercano a Quibanga, provincia de Sange, Angola, 6.1857, 3435 (BM).
Talo costroso, crecimiento hemisférico o irregular, color guinda. Altura del talo 25.0-156.0 µm; filamentos escasamente ramificados, 3-14 células por filamento. Talo polistromático con largo celular 3.0-11.7 µm (5.6 ± 0.8 µm), diámetro celular 2.3-7.4 µm (4.1 et al. 0.4 µm). Propágulos vegetativos 13.0-79.5 µm de largo a 27.0-103.2 µm de diámetro.
Material examinado: Tamaulipas, Poza Azul, J. Carmona, 27.5.1993 (FCME PA3442); San Luis Potosí, Nacimiento el Salto, G. Montejano, 15.3.1984 (FCME PA2173); G. Montejano, 25.3.1987 (FCME PA2601); Ciudad Valles, Micos, J. Carmona, 17.02.1992 (FCME PA3320); Tamasopo, Puente de Dios, G. Montejano, 8.11.1987 (FCME PA2868); J. Carmona, 7.11.1990 (FCME PA3230); Tamasopo, río Tamasopo, G. Montejano, 27.5.1987 (FCME PA2741); Aquismón, Santa Anita, G. Montejano, 7.5.85 (FCME PA2460); G. Montejano, 26.3.1987 (FCME PA2623); Tanchachín, G. Montejano, 7.09.1981 (FCME PA780); G. Montejano, 3.4.1982 (FCME PA1010); Ciudad Valles, Puente de Dios II, J. Carmona, 9.5.85 (FCME PA2547); Cd. Valles, Choy, J. Carmona, 13.4.1988 (FCME PA2922); J. Carmoma, 9.9.1989 (FCME PA3168); J. Carmona, 1.5.1991 (FCME PA3259); Coy, G. Montejano, 24.3.1987 (FCME PA2585); Aquismón, La Garita, J. Carmona, 6.7.1995 (FCME PA3576); J. Carmona, 25.11.1995 (FCME PA3620); Tambaque, G. Montejano, 20.2.1992 (FCME PA3357); J. Carmona, 25.11.1995, (FCME PA3621); Veracruz, Poza Reina, J. Carmona, 12.3.1993 (FCME PAPs/n); Morelos, Las Estacas, G. Montejano, 27.1.1996 (FCME BALE3023).
Ecologia y distribución
Las localidades mexicanas estudiadas se presentaron en 14 corrientes (Tabla 3). 8 manantiales y 6 ríos de primer orden, con clima cálido a semicálido, Las poblaciones de H. angolensis se registraron en condiciones ecológicas similares: temperatura del agua elevada de 23°C a 27°C, pH de 7.0 a 8.0, conductividad alta de 750 a 1,500 µS cm−1, alcalinidad total de 370 a 554 mg l−1, intensidad luminosa muy baja a media y velocidad de corriente de 30 a 130 cm s−1. El substrato más frecuente fue roca calcárea y ocasionalmente sobre roca ígnea y vidrio. Comúnmente creció junto a otras algas como Hyella cf. fontana, Pleurocapsa minor, Thorea hispida y esporádicamente con Blennothrix ganeshii, A. huastecana, fases 'Chantransia' y Sirodotia huillensis. Las 22 poblaciones mexicanas crecieron a lo largo de todo el año y se observaron como habitantes perennes de manantiales con cavernas a ríos con alta cobertura de la vegetación. No se registró la abundancia del crecimiento, sin embargo, en las localidades Nacimiento el Salto, Puente de Dios y Choy se observó que la costra guinda cubrió 4 m2 de la superficie de las rocas.
Discusión
La valoración y análisis de 23 poblaciones permitió delimitar dos especies: Hildenbrandia angolensis de la región central de México e H. rivularis de la República Checa. En cuanto a los caracteres estudiados, la mayoría fue variable incluso dentro de la misma población, tal como: la altura del talo, número de células en el filamento, forma celular y dimensiones del propágulo vegetativo, por lo que no se consideraron caracteres con valor taxonómico. Por el contrario, el promedio del diámetro celular de los filamentos erectos fue relativamente constate y se pudo reconocer diferencia entre las poblaciones mexicanas y la de Europa central. El promedio del diámetro celular, propuesto por Sheath et al. (1993), puede presentar traslapes considerables si se consideran los tres tipos de forma celular. La formación y liberación de propágulos vegetativos sugiere que son de tipo endógeno tal como sugiere Sherwood y Sheath (2000b) y diferente a lo descrito por Seto (1977). La reproducción se registró en todos los manantiales sin alguna tendencia estacional. Esto puede estar explicado por la poca diferencia ambiental a lo largo del año, tal como se describe en las poblaciones registradas por Sherwood y Sheath (2000b).
En cuanto a ecología se refiere, las localidades mexicanas fueron muy homogéneas en temperatura, iluminación, substrato y conductividad, muy distinta a la localidad europea donde hay una diferencia en la temperatura de 8°C. Esto parece apoyar que existen dos especies con requerimientos ecológicos diferentes: H. rivularis que se presentó con temperatura baja e H. angolensis que se desarrolló óptimamente con temperatura mayor a 23 "C. Nuestros sitios de recolecta coinciden en general con las características ambientales de localidades en otros países donde se ha descrito H. angolensis, tales como: Angola (West y West, 1897), Estados Unidos de Norteamérica, Belice, Martinica, Saint Vincent, Costa Rica, República Dominicana, Jamaica, Granada, Dominica, Puerto Rico, Isla Guadeloupe (Sheath et al., 1993), Brasil (Necchi, 1987; Necchi et al., 1999) y sitios de regiones tropicales de otros países en donde se ha registrado H. rivularis, aunque tiene que confirmarse su identidad taxonómica, tales como: Indonesia (Skuja, 1938), Malasia (Kumano, 1978), Irak (Hinton y Maulood, 1980), Isla Guadeloupe (Bourrelly y Manguin, 1952), Puerto Rico (Wille, 1915) y Cuba (Starmach, 1986-1987). La distribución de H. angolensis en la región central de México fue heterogénea. La distribución más amplia se presentó en la cuenca baja del río Pánuco en los límites de la planicie costera del Golfo de México y la Sierra Madre Oriental, que es una región cárstica con factores ambientales muy homogéneos. Por primera ocasión se registraron poblaciones de Hildenbrandia en la cuenca alta del río Balsas y cuenca baja del río Papaloapan, también se presentaron en condiciones de alta conductividad relacionadas con el afloramiento de roca calcárea. Por el contrario, no se ha registrado en la cuenca alta del valle de México, seguramente por diferencias en las condiciones ecológicas (Tamayo, 1981; Ferrusquía-Villafranca, 1993; Ramírez et al., 2001).
El esquema filogenético de Sherwood y Sheath (1999) basado en el análisis genético de los fragmentos rbcL y 18S rRNA mostró que las especies de agua dulce y marinas en Norteamérica son parafiléticas. La base del esquema son poblaciones marinas, que por invasión múltiple colonizaron ambientes de agua dulce. La cercanía de la Sierra Madre Oriental, la planicie costera del golfo y el golfo de México, así como la historia geológica de la región (Ferrusquía-Villafranca, 1993), sugieren que las poblaciones de agua dulce en las cuencas bajas de los ríos Pánuco y Papaloapan representan formas de invasión por alguno de los mecanismos propuestos por Skuja (1938) y Sherwood y Sheath (1999): como colonización y permanencia durante largo tiempo en estuarios; o por poblaciones que quedaron aisladas en sitios que se elevaron por la deriva continental, o bien, por el transporte de aves o insectos. Este último mecanismo parece explicar la presencia de Hildenbrandia en el estado de Morelos, ya que es un manantial rodeado de coordilleras.
Las poblaciones de H. angolensis en la región central de México resultaron habitantes frecuentes de zonas con clima tropical y ambientes cársticos. Este trabajo representa el primer estudio detallado sobre su taxonomía y distribución en la región central de México, sin embargo, aún es necesario realizar estudios para evaluar e interpretar la ecología y su relación filogenética con poblaciones marinas.
Agradecimientos
Agradecemos a Michele Gold Morgan y al Dr. Filip Lederer por haber proporcionado el material del río Balsas y de la República Checa respectivamente. A Eberto Novelo y Orlando Necchi Jr. por la revisión y valiosos comentarios al manuscrito. Proyecto financiado por proyecto PAPIIT-DGAPA (IN2297) de JCJ.
Literatura citada
BOURRELLY, P., 1970. Les algues d'eau douce. Initiation a la Systématique Les algues bleues et rouges. T. III. Boubée et Cie. París. 512 p. [ Links ]
BOURRELLY, P. y F. MANGUIN, 1952. Algues d'Eau Douce de la Guadeloupe. Société d'Edition d'Enseignement Supéieur, Paris. 281 p. [ Links ]
CARMONA, J. J. y G. MONTEJANO, 1993. Caracterización algal en manantiales de la Huasteca Potosina, México. Boletín de la Sociedad Botánica de México 53: 21-41. [ Links ]
FERRUSQUÍA-VlLAFRANCA, I., 1993. Geology of Mexico: A Synopsis. pp. 3-107. In: RAMAMORTHY, T. P., R. BYE, A. LOT y J. FA (Eds). Biological Diversity of Mexico, Origins and Distribution. Oxford University Press. New York. [ Links ]
FLINT, L. H., 1955. Hildenbrandia in America. Phytomorphology 5: 185-189. [ Links ]
HINTON, G. C. y B.K. MAULOOD, 1980. Freshwater red algae, a new addition to the Iraqi flora. Nova Hedwigia 33: 487-497. [ Links ]
HOLMGREN P. K., N. H. HOLMGREN y L. C. BARNETT, 1990. Index Herbariorum. Part I. The Herbaria of the World. 8th edition. New York Botanical Garden, New York. 693 p. [ Links ]
INSTITUTO NACIONAL DE ESTADÍSTICA GEOGRAFÍA E INFORMÁTICA, 1985. Síntesis Geográfica de San Luis Potosí. Secretaría de Programación y Presupuesto. México. 186 p. [ Links ]
INSTITUTO NACIONAL DE ESTADÍSTICA GEOGRAFÍA E INFORMÁTICA, 1987. Síntesis Geográfica del estado de Puebla. Secretaría de Programación y Presupuesto. México. 56 p. [ Links ]
ISRAELSON, G., 1942. The freshwater Floridae of Sweden. Studies on their taxomomy, ecology and distribution. Symbolae Botanicae Up-saliensis 6 (1): 1-135. [ Links ]
JAP, C., 1941. Studies on the freshwater algae of China VII. A preliminary account of the Chinese Freshwater Rhodophyceae. Sinensia 12 (1-6): 245-290. [ Links ]
JOHANSSON, C., 1982. Attached algal vegetation in running waters of Jamtland, Sweden. Acta Phytogeographica Suecica 71:1-80. [ Links ]
KHAN, M., 1974. On a fresh water Hildenbrandia Nardo, from India. Hydrobiologia 44 (2-3): 237-240. [ Links ]
KUMANO, S., 1978. Notes on freshwater red elgae from West Malasya. Botanical Magazine, Tokyo 91: 97-107. [ Links ]
MONTEJANO, Z. G., J. J. CARMONA y U. E. CANTORAL, 2000. Algal communities from calcareous springs and streams in La Huasteca: a Synthesis. pp. 139-145. In: MUNAWAR, M., S. LAWRENCE, I. F. MUNAWAR y D. MALLEY (Eds). Aquatic Ecosystems of Mexico: Status and Scope. SPB Academic Publishing. Amsterdam. [ Links ]
NECCHI, O. JR., 1987. Estudos sobre as Rhodophyta de águas continentais do Brasil. VI. Ocorrência de Hildenbrandia rivularis (Liebmann) J. Agardh nos estados do Rio Janeiro e São Paulo. Nerítica, Pontal do Sul 2 (Suppl.): 1147-1152. [ Links ]
NECCHI O. JR., C. C. Z. BRANCO y L. H. Z. BRANCO, 1999. Distribution of Rhodophyta in streams from São Paulo State, southeastern Brazil. Archiv fü Hydrobiologie 147: 79-89. [ Links ]
NICHOLS, H. W., 1965. Culture and development of Hildenbrandia rivularis Denmark and North America. American Journal of Botany 52(1): 9-15. [ Links ]
RAMÍREZ, V. M., M. Y. BELTRÁN, G. M. BOJORGE, J. J. CARMONA, U. E. CANTORAL y F. VALADEZ, 2001. Flora algal del río La Magdalena, México. Boletín de la Sociedad Botánica de México 68: 51-73. [ Links ]
SECRETARÍA DE PROGRAMACIÓN Y PRESUPUESTO, 1981. Síntesis Geográfica del Estado de Morelos. México. 110 p. [ Links ]
SETO, R., 1977. On the vegetative propagation of a freshwater red alga, Hildenbrandia rivularis (Liebm.) J. Ag. Bulletin of the Japanese Society of Phycology 25: 129-136. [ Links ]
SHEATH, R. G., D. KACZMARCZYK y K. M. COLE, 1993. Distribution and systematics of freshwater Hildenbrandia (Rhodophyta, Hildenbrandiales) in North America. European Journal of Phycology 28: 115-121. [ Links ]
SHERWOOD A. R. y R. G. SHEATH, 1999. Biogeography and systematics of Hildenbrandia (Rhodophyta Hildenbrandiales) in North America: inferences from morphometrics and rbcL and 18S rRNA gene sequence analyses. European Journal of Phycology 34: 523-532. [ Links ]
SHERWOOD A. R. y R. G. SHEATH, 2000a. Biogeography and systematics of Hildenbrandia (Rhodophyta, Hildenbrandiales) in Europe: inferences from morphometrics and rbcL and 18S rRNA gene sequence analyses. European Journal of Phycology 35:143-152. [ Links ]
SHERWOOD A. R. y R. G. SHEATH, 2000b. Microscopic analysis and seasonality of gemma production in the freshwater red alga Hildenbrandia angolensis (Hildenbrandiales, Rhodophyta). Phycological Research 48: 241-249. [ Links ]
SKUJA, H., 1938. Die Süsswasserrhodophyceen der Deutschen Limnologischen Sunda-Expedition, Archiv für Hydrobiologie, Suppl-Bd. 15: 603-637. [ Links ]
SOUTHWOOD, T. R., 1978. Ecological Methods with particular reference to the study of Insect Populations. Chapman and Hall. New York. 524 p. [ Links ]
STARMACH, K., 1952. The reproduction of the freshwater Rhodophyceae Hildenbrandia rivularis (Liebm.) J. Ag. Acta Societatis Botanicorum Poloniae 21(3): 445-474. [ Links ]
STARMACH, K., 1969. Hildenbrandia rivularis i glony towarzyszan w potoku Cedronka KoloWejherowa (województwo Gdansk). Fragmenta Floristica et Geobotanica 15(3): 387-398. [ Links ]
STARMACH, K., 1977. Flora Slodkowodna Polski. Tomo 4: Phaeophyceae - Rhodophyceae. Polska Akademia Nauk. Warszawa-Kraków. 445 p. [ Links ]
STARMACH, K., 1986-1987. Some taxa of freshwater red algae (Rhodophyta) from Cuba. Fragmenta Floristica et Geobotanica 31-32 (3-4): 474-494. [ Links ]
TAMAYO, J. L., 1981. Geografía Moderna de México. 9a. Ed. Edit. México, D.F. Trillas. 400 p. [ Links ]
TARAS, M. J., A. E. GREENBERG, R. HOAK y M. C. RRAND, 1971. Standar Methods. American Public. Health Association. 13a. Ed. 74 p. [ Links ]
WEST, W. y G. S. WEST, 1897. Welwitsch's African freshwater algae. Journal of Botany 35: 1-7. [ Links ]
WILLE, N., 1915. Report on an expedition to Puerto Rico for collecting fresh-water algae. Journal of the New York Botanical Garden 16: 132-146. [ Links ]
WOLLE, F., 1887. Freshwater algae of the United States. The Comenius Press. Bethlehem. Pennsylvania. 61 p. [ Links ]

