










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.10 no.2 Ciudad de México Dez. 2000
Artículos
El Canal de Ballenas, Baja California, como ambiente favorable para el desarrollo del fitoplancton
Eduardo Millán-Núñez1 y Clarice M. Yentsch2
1 Centro de Investigación Científica y de Educación Superior de Ensenada. División de Oceanología. Departamento de Ecología. Apdo. Postal 2732, Ensenada, Baja California, C. P. 22830, México. E-mail: emillan@cicese.mx.
2 Bigelow Laboratory for Ocean Sciences, McKown Point, West Boothbay Harbor, Maine, 04575. U. S. A. E-mail: Yentsch@alpha1.bigelow.org.
Recibido: 10 de marzo de 2000
Aceptado: 4 de septiembre de 2000
Resumen
Se tomaron muestras de fitoplancton en los canales de Ballenas y Salsipuedes en la parte norte-central del Golfo de California. Se identificaron en los canales veintiseis especies de diatomeas en 15 géneros, dominando la especie Thalassioneme frauenfeldii (Grunow) Tempére y Peragallo 1910 ex-Hallegraeff 1986; así como 13 géneros de dinoflagelados y dos de silicoflagelados. La mayoría de estos organismos estuvieron incluidos en las masas de agua denominadas Agua del Golfo de California (AGC) y Agua Subsuperficial Subtropical (ASsSt). Los análisis cualitativos de las diatomeas no presentaron diferencias significativas entre las estaciones del Canal de Ballenas; a diferencia de los dinoflagelados que sí presentaron diferencias entre las estaciones en respuesta a la estabilidad de la columna de agua. La relación inversa entre los nutrientes y abundancia de diatomeas, se debió principalmente a la inestabilidad de la columna de agua, ya que el fitoplancton necesita áreas con menor energía de mezcla para poder acondicionarse a la luz. En conclusión el Canal de Ballenas mostró ser una zona dinámica que provee los nutrientes necesarios para el crecimiento del fitoplancton.
Palabras clave: Canal de Ballenas, fitoplancton, nutrientes, mezcla de la columna de agua.
Abstract
Phytoplankton samples were taken in the Ballenas and Salsipuedes Channels in the northern-central part of the Gulf of California. Twenty-six diatom species in 15 genera were identified in the channels, with dominance of the species Thalassionema frauenfeldii (Grunow) Tempére and Peragallo 1910 ex-Hallegraeff 1986; as well as 13 dinoflagellate genera and two silicoflagellates. Most of these organisms were included in the water masses named Gulf of California Water (GCW) and Subsurface Subtropical Water (SsStW). The quali-quantitative diatom analyses did not show significant differences between stations of the Ballenas Channel; contrary to dinoflagellates which showed differences between the stations in answer to stability of the water column. The inverse relationship between nutrients and diatom abundance, was mainly due to the instability of the water column since phytoplankton needs areas with less mixing energy for light conditioning. In conclusion, Ballenas Channel showed to be a dynamic zone that provides the necessary nutrients for phytoplankton growth.
Key words: Ballenas Channel, phytoplankton, nutrients, water column mixing.
Introducción
A la parte central-norte del Golfo de California se le ha considerado como una zona de alta productividad primaria, comparable a áreas donde ocurren eventos de sugerencias (Zeitzschel, 1969; Álvarez-Borrego y Lara-Lara, 1991; Millán-Núñez et al. 1993; Valdéz-Olguín, 1998). Asimismo, se ha demostrado que en esta área la energía de marea se disipa con gran velocidad. Los valores máximos de disipación de energía se presentan en la zona de las grandes islas (Tiburón y Ángel de la Guarda) hasta >0.3 W m-2, lo que provoca corrientes muy fuertes en mareas vivas (>1.5 m s-1) principalmente en los canales (Badán-Dangón et al., 1991). Al sur de las grandes islas los valores de disipación de energía de marea son despreciables (Argote et al., 1995). La alta energía en la zona de las grandes islas causa una mezcla vertical muy intensa, lo cual a su vez produce las más altas concentraciones superficiales de nutrientes del Golfo (Álvarez- Borrego et al., 1978). Por otro lado, el régimen de vientos del noroeste prevalecen de noviembre a mayo causando sugerencias intensas en la costa oriental (condiciones de invierno); y los vientos del sureste prevalecen el resto del año, causando la entrada al golfo del agua tropical del Pacífico Oriental con características oligotróficas (condición de verano).
Por lo anteriormente expuesto y, previo conocimiento de la fisiología del fitoplancton, se considera que la zona de las grandes islas poseen características ambientales favorables para el desarrollo del fitoplancton (Gilmartin y Revelante, 1978; Millán-Núñez y Gaxiola-Castro, 1989; Millán-Núñez, 1992). Sin embargo, no es suficiente la información acerca del origen de los nutrientes y del comportamiento ecológico del fitoplancton en los canales; por lo que hemos implementado un estudio de las diferentes variables físico-químicas que afectan al fitoplancton en los canales Ballenas y Salsipuedes.
Materiales y métodos
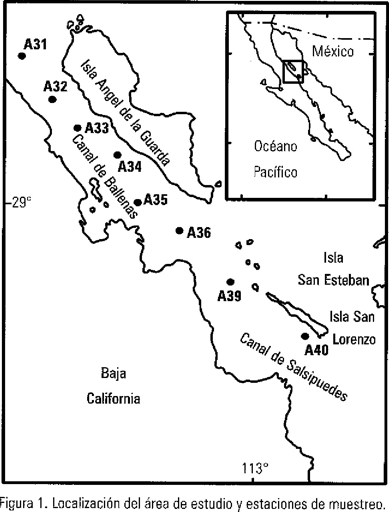
Los canales Ballenas y Salsipuedes se localizan en el Golfo de California entre la Península de Baja California y la Isla Ángel de la Guarda (Fig. 1). Tienen aproximadamente 125 km de largo por 20 km de ancho, con una profundidad máxima que excede los 1600 m, sin embargo, la parte sur del Canal de Salsipuedes tiene una profundidad de... 450 m (Rusnak et al. 1964) lo que limita la circulación profunda entre los canales y la parte sur del Golfo. En este estudio, se tomaron un total de 80 muestras de agua de mar a partir de una línea de ocho estaciones con 10 profundidades (0, 10, 25, 50, 75, 100, 200, 300, 400 y 500 m) durante el 2 y 3 de marzo de 1988 a bordo del B/O USN De'Steiguer.
Las muestras de fitoplancton se tomaron con botellas Niskin de 1.7 litros. Inmediatamente después se pusieron en botellas obscuras de plástico de 250 ml con una solución de formalina al 4% saturado con borato de sodio, posteriormente se vertieron 50 ml en una cámara de sedimentación agregándole tres gotas de Rosa-Bengala. Las especies y la abundancia del fitoplancton fueron analizadas con un microscopio invertido Zeiss con aumento de 160 y 400 (Utermöhl, 1958). Las células que formaron cadenas se contaron en forma individual. La comunidad del fitoplancton se ordenó y se clasificó en cuanto a su abundancia relativa, tratando por separado los grupos de diatomeas y dinoflagelados; para tal efecto se utilizó la ecuación de amplitud del nicho ecológico (Levins, 1978).
Bi = exp [ - ∑ (Pij / Rj) In (Pij / Rj) ]
donde, Bi es la amplitud de nicho; Pij es la proporción de la especie i en la muestra j; y Rj es la suma de sus proporciones en cada punto o tiempo de muestreo.
Los análisis de clorofila a se llevaron a cabo con un fluorómetro Turner Desings utilizando la técnica descrita por Holm-Hansen et al. (1965). Los nutrientes se analizaron en un espectrofotómetro Bausch & Lomb Spectronic 1001 (Strickland y Parsons, 1972). Se utilizó la imagen de temperatura superficial por satélite (NOAA-11, AVHRR: Advance Very High Resolution Radiometer). La resolución espacial del dispositivo fue un arreglo de pixel (picture element) de 1 km × 1 km.
Se utilizaron los datos del CTD Neil Brown MK III para realizar un diagrama T-S y detectar en los canales Ballenas y Salsipuedes las diferentes masas de agua; se compararon los resultados obtenidos con el banco de datos de 1939-1994 reportados por Lavín et al. (1997). La estabilidad de la columna de agua (Neuman y Pierson, 1966) se obtuvo a partir de los 50 m de profundidad, para evitar las anomalías que produce la capa superficial del mar por el forzamiento atmosférico y la radiación solar. Para determinar el comportamiento de las variables entre estaciones, así como la correlación entre variables, utilizamos el programa STATISTICA versión 6.0.
Resultados y discusiones
El diagrama T-S de los canales de Ballenas y Salsipuedes (Fig. 2) presentó dos masas de agua bien definidas, con un intervalo en la densidad del agua de 26.2-26.5 de σt. El Agua Subsuperficial Subtropical (ASsSt) que se localizó por debajo de los 350 m con salinidad menor a 34.9 ups (unidades prácticas de salinidad) y 11.5 °C, y el Agua del Golfo de California (AGC) que se caracteriza como una agua relativamente tibia y de alta salinidad (>34.90 ups), son masas de agua previamente descritas por Torres-Orozco (1993), Griffiths (1968) y Stevenson (1970). En este estudio no se encontraron diferencias físico-oceanográficas entre las estaciones de la línea de muestreo (A31 a la A39). Sin embargo, la estación A40 mostró el mismo promedio superficial de temperatura, pero con diferencias menores de 0.025 ups (Tabla 1). Esta diferencia de salinidad en superficie, se debió posiblemente a la influencia del Agua Superficial Ecuatorial (ASE) que se transportó hacia el norte del Canal Salsipuedes por medio de procesos advectivos locales. Por debajo de los 200 m de profundidad, la estación A40 mostró condiciones similares al resto de las estaciones.

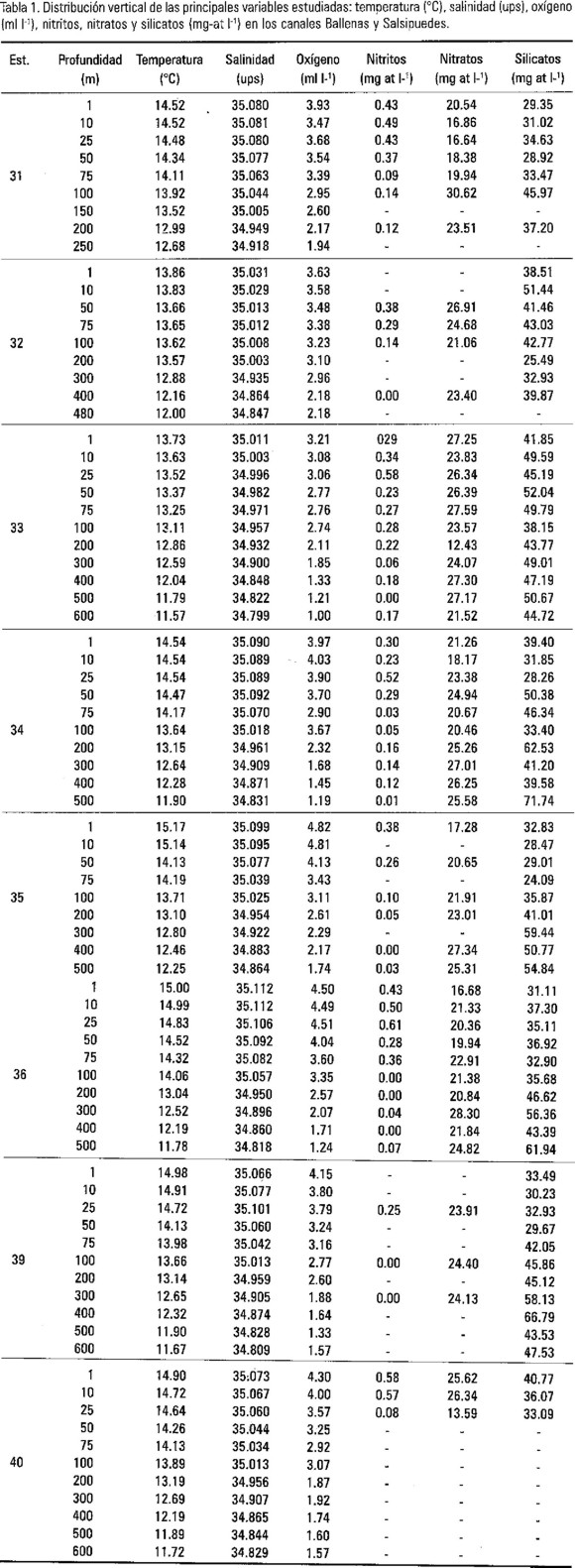
Asimismo, se puede observar para superficie, que el agua con bajas temperaturas proceden del Canal de Ballenas (Fig. 3, estación A32 y A33) y frente a la Isla San Esteban; posiblemente ocasionadas por el efecto de mezcla por marea. De igual manera, la imagen de temperatura superficial por satélite (no se presenta) permite detectar un desplazamiento de agua fría hacia el sur de las grandes islas, lo que provoca un frente con el agua cálida proveniente del sur del Golfo de California.

La mayor inestabilidad de la columna de agua, se localizó principalmente en las estaciones A32 y A33 con valores de energía muy similares entre sí (Figs. 4a y 4b, Tabla 1). La inestabilidad se asocia a cambios verticales de la columna de agua, lo que complica los procesos físicos oceanográficos, tal y como lo sugirió Badán-Dangón et al. (1985) y Lozano-Oaxaca (1990) de que el forzamiento de la marea (> 4 m de amplitud) y los procesos atmosféricos son los que originan la mezcla y circulación del agua en el Golfo de California.
En los canales de ballenas y Salsipuedes se detectaron un total de 26 especies de diatomeas en 15 géneros, así como 13 géneros de dinoflagelados y dos silicoflagelados (Tabla 2). En este estudio, mostramos un mayor interés en el grupo taxonómico de las diatomeas, porque históricamente el Canal de Ballenas se ha caracterizado por ser una zona muy dinámica (Badán-Dangón et al., 1991; Lozano-Oaxaca, 1990; Simpson et al. 1994); por lo tanto, no favorable a la sucesión fitoplanctónica (Margalef, 1958). Las primeras cinco especies de diatomeas de la Tabla 2 mostraron una mayor persistencia a las condiciones hidrográficas de los canales Ballenas y Salsipuedes que el resto de la comunidad. La diatomea dominante fue Thalassionema frauenfeldii, especie que se caracteriza por aclimatarse en aguas frías menores de 14.5 (C (Round, 1967; Hallegraeff, 1986); estas temperaturas bajas se encuentran comunmente en las aguas subsuperficiales del Canal de Ballenas. Asimismo, Gárate-Lizarraga et al. (1990) reportaron para otoño al sur de las grandes islas al mismo organismo como especie dominante y con distribución cosmopolita.
En general, la distribución vertical de las diatomeas y dinoflagelados mostraron una relación inversa con relación a la profundidad (Figs. 5 y 6). Sin embargo, a los 50 m de profundidad en la estación A35 se detectó un incremento en la abundancia de diatomeas de... 18 × 103 células l-1, valores similares a los reportados en otoño por Gárate-Lizarraga et al. (1990) y en verano por López-Vergara (1993); y menores en un orden de magnitud que los reportados para primavera por Álvarez-Borrego y Gaxiola-Castro (1987), y Cortés-Lara, et al. (1999). La abundancia de células de diatomeas reportadas en primavera al sur de las Islas San Esteban y San Lorenzo por Millán-Núñez y Gaxiola-Castro (1989) fueron más altos que los reportados en este estudio.


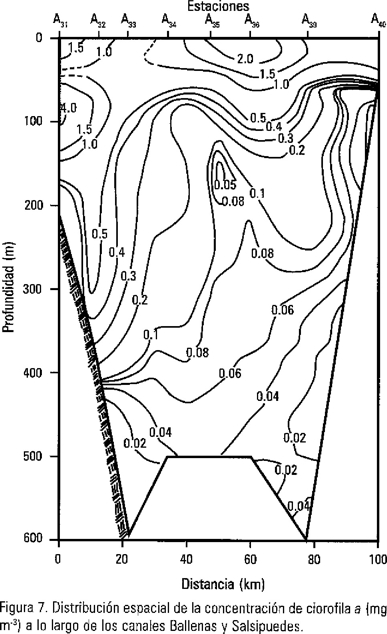
La concentración de clorofila a en este estudio, mostró valores altos en superficie y, en algunos casos, máximos a 75 y 100 m hasta de 4.83 mg m-3 en la estación A31 (Fig. 7). Lo que ocasiona un acondicionamiento del fitoplancton a bajas irradiancias, que disminuye su capacidad fotosintética, y una fotosíntesis nula cuando las células se encuentran a profundidades grandes. Sin embargo, en la mayoría de las estaciones del Canal de Ballenas se observaron bajas densidades de las células; esto posiblemente se debió a los procesos advectivos y turbulentos que tienden a homogeneizar la columna de agua en el canal (Fig. 4), lo que provoca una disminución en la penetración de la luz y como consecuencia un incremento en la diversidad pigmentaria por unidad de organismo (Fallowski, 1984; y Millán-Núñez et al., 1998). La disminución espacio-temporal de la irradiancia descendente registrada por Millán-Núñez (1999) para la zona de las grandes islas, han mostrado la gran variabilidad del fondo de la zona eufótica (1% de la irradiancia que incide en la superficie) de 30 m, 25 m, y 48 m para abril de 1993, octubre de 1994, y agosto de 1995 respectivamente; por lo que se considera que una gran parte de los pigmentos fotosintéticos de la columna de agua en este estudio, escasamente podrían estar fotosintetizado.
La concentración de fosfatos, silicatos, nitritos y nitratos (Fig. 8, Tabla 1) mostraron una relación inversa en relación al grupo de diatomeas, esta relación posiblemente se debió a que la columna de agua presentó inestabilidad; por lo que, la alta concentración de nutrientes que se detectó en el Canal de Ballenas no pudo ser asimilado por el fitoplancton, debido a la escasa disponibilidad de luz para llevar a cabo la fotosíntesis.

El análisis de varianza (one-way ANOVA), mostró diferencias significativas entre estaciones para los grupos de diatomeas y dinoflagelados (F=2.67, p<.017; y F=2.90, p<.011, respectivamente); por tal motivo, se aplicó la prueba de asociación de Tukey HSD (Honest Significant Difference) para determinar en dónde se presentaron las diferencias (Figs. 9a y 9b). Los resultados mostraron para el grupo de las diatomeas, que las diferencias se localizaron solamente en la estación A32; ya que si se elimina esta estación, estadísticamente no se presentarían diferencias significativas en las estaciones. No así, el grupo de los dinoflagelados que mostraron diferencias significativas entre las estaciones A32 y A35, observándose una mayor estabilidad de la columna de agua de la estación A35 (Fig. 4d); lo que representó condiciones favorables para el desarrollo de los dinoflagelados.

Con la información anteriormente expuesta, se concluye que el Canal de Ballenas es una zona de alta inestabilidad en la columna de agua, y que además provee los nutrientes necesarios que requiere el fitoplancton para crecer. Sin embargo, el constante movimiento de la columna de agua limita la aclimatación de las células de fitoplancton para una mejor fotosíntesis.
Agradecimientos
A Ken Ritcher de NOSC (Naval Ocean System Center, de San Diego, CA.) por invitarnos a participar en el crucero SOMAP II. De igual manera a Emilio Palacios por la elaboración de datos sobre estabilidad de la columna de agua. A Jósé M. Domínguez y Fco. Javier Ponce por los dibujos aquí presentados. Asimismo, a dos revisores anónimos por sus sugerencias y comentarios.
Literatura citada
Álvarez-Borrego, S. y G. Gaxiola-Castro, 1987. Photosynthetic parameters of northern Gulf of California phytoplankton. Continental Shelf Research 277: 1-11. [ Links ]
Álvarez-Borrego, S. y J. R. Lara-Lara, 1991. The phisical environment and primary productivity of the Gulf of California, pp. 555-567. En: J. P. Dauphin y B. R. T. Simoneit (Comps.). The Gulf and Peninsular Provinces of the Californias. American Association Petrology Geology Memoir 47. 834 p. [ Links ]
Álvarez-Borrego, S., J. A. Rivera, G. Gaxiola-Castro, M. J. Acosta-Ruíz y R. Schwartzlose, 1978. Nutrientes en el Golfo de California. Ciencias Marinas 5: 53-71. [ Links ]
Argote, M. L., A. Amador, M. F. Lavín y J. R. Hunter, 1995. Tidal dissipation and stratification in the Gulf of California. Journal of Geophysical Research 100: 16103-16118. [ Links ]
Badán-Dangón, A., C. E. Dorman, M. A. Merrifield y C. D. Winant, 1991. The lower atmosphere over the Gulf of California. Journal of Geophysical Research 96: 16877-16896. [ Links ]
Badán-Dangón, A., C. J. Koblisky y T. Baumgarter, 1985. Spring and summer in the Gulf of California: observations of surface thermal patterns. Oceanologica Acta 8: 13-22. [ Links ]
Cortés-Lara, M. del C., S. Álvarez-Borrego y A. D. Giles-Guzmán, 1999. Efecto de la mezcla vertical sobre la distribución de nutrientes y fitoplancton en dos regiones del Golfo de California, en verano. Revista Sociedad Mexicana de Historia Natural 49: 193-206. [ Links ]
Falkowski, P. G., 1984. Physiological responses of phytoplankton to nature light regimes. Journal Plankton Research 6: 295-302. [ Links ]
Gárate-Lizárraga, I., D. A. Siqueiros-Beltrones y C. H. Lechuga-Deveze, 1990. Estructura de las asociaciones microfitoplanctónicas de la región central del Golfo de California en otoño de 1986. Ciencias Marinas 16(3): 131-153. [ Links ]
Gilmartin, M. y N. Revelante, 1978. The phytoplankton characteristics of barrier Island Lagoons of the Gulf of California. Estuarine Coastal Marine Science 7: 29-47. [ Links ]
Griffiths, R. C., 1968. Physical, chemical and biological oceanography of the entrance to the Gulf of California, Spring of 1960. Special Scientific Report. U. S. Fish & Wildlife Service 573, 47 p. [ Links ]
Hallegraeff, G. M., 1986. Taxonomy and morphology of the marine plankton diatoms Thalassionema and Thalassiothrix. Diatom Research 1(1): 57-80. [ Links ]
Holm-Hansen, D., C. J. Lorenzen, R. V. Holmes y J. D. J. Strickland, 1965. Fluorometric determination of chlorophyll. Journal Cons. International pour L'exploration de la mer 30: 3-15. [ Links ]
Lavín, M. F., E. Beier y A. Badán, 1997. Estructura hidrográfica y circulación del Golfo de California: Escalas estacional e interanual. Contribuciones a la Oceanografía Física en México. Monografía No. 3. Unión Geofísica Mexicana, pp. 141-171. [ Links ]
Levins, R., 1978. Evolution in changing environments. Princeton University Press Princeton, pp. 39-65. [ Links ]
López Vergara, A. T., 1993. Caracterización de los principales factores físico-químicos que afectan la distribución del fitoplancton en la parte central norte del Golfo de California. Tesis de Licenciatura, Facultad de Ciencias Marinas, UABC. México, 66 p. [ Links ]
Lozano-Oaxaca, A. R., 1990. Variación estacional de la estratificación en el Canal de Ballenas. Tesis de Licenciatura. Facultad de Ciencias Marinas, UABC. México, 45 p. [ Links ]
Margalef, R., 1958. Temporal succesion and spatial heterogeneity in phytoplankton. En: Buzzati-Traverso (De), Perspectives in marine biology. University of California Press, Berkeley, 621 p. [ Links ]
Millán-Núñez, E., 1992. Spatial distribution of the phytoplankton in the northern Gulf of California. Ciencias Marinas 18(2): 101-117. [ Links ]
Millán-Núñez, E., 1999. Variabilidad del coeficiente específico de absorción de luz por fitoplancton en el Golfo de California. Tesis de Doctorado en Ciencias. CICESE. México, 131 p. [ Links ]
Millán-Núñez, E. y G. Gaxiola-Castro, 1989. Spatial variability of phytoplankton in the Gulf of California during the El Niño 1983, Nova Hedwigia 49: 113-119. [ Links ]
Millán-Núñez, E., J. R. Lara-Lara y J. S. Cleveland, 1998. Variations in specific absoptioncoefficient and total phytoplankton in the Gulf of California. CalCOFI Report 39: 157-167. [ Links ]
Millán-Núñez, R., R. Cajal-Medrano, E. Santamaría del Angel y E. Millán-Núñez, 1993. Productividad primaria y clorofila a en la parte central del Golfo de California (otoño 1987). Ciencias Marinas 19(1): 29-40. [ Links ]
Neuman, G. y W. Pierson, 1966. Principles of physical oceanography. Prentice-Hall, inc., Engelwood Cliffs, N. J. 545 p. [ Links ]
Round, F. E., 1967. The phytoplankton of the Gulf of California. Part. Its compositions, distributions and contributions to the sediments. Journal Exp. Marine Biology Ecology 1: 76-97. [ Links ]
Rusnak, G. A., R. L Fisher y F. P. Shepard, 1964. Bathymetry and faults of Gulf of California, pp. 59-75. En: Tj. H. Van Andel y G. G. Shor (Eds). Marine Geology of the Gulf of California. American Association of Petoleum Geologist. [ Links ]
Simpson, J. H., A. J. Souza y M. F. Lavín, 1994. Tidal mixing in the Gulf of California, pp. 169-182. En: K. J. Beven, P. C. Chatwin y J. H. Millbank (Eds.). Mixing and transport the Environment. John Wiley, New York, 467 p. [ Links ]
Stevenson, M. R., 1970. On the physical and biological oceanography near the entrance of the Gulf of California, October 1966-August 1967. Inter-American Tropical Tuna Commission Bulletin 14(3): 389-504. [ Links ]
Strickland, J. D. y T. R. Parsons, 1972. A practical handbook at seawater analysis. 2nd ed. Bulletin Fishieries Research Board, Canada, 310 p. [ Links ]
Torres-Orozco, E., 1993. Análisis volumétrico de las masas de agua del Golfo de California. Tesis de Maestría en Ciencias, CICESE. México, 80 p. [ Links ]
Utermöhl, H. 1958. Zur vervollkommnung der quantitativen phytoplankton methodik. Mitt. Int. Verein Theor. Angew. Limnology 9: 1-38. [ Links ]
Valdéz-Olguín, J. E., 1998. Variabilidad de los parámetros fotosintéticos del fitoplancton en función de la climatología de la zona eufótica. Tesis de Doctorado en Ciencias, CICESE. México 170 p. [ Links ]
Zeitschel, B., 1969. Primary productivity in the Gulf of California. Marine Biology 3: 201-207. [ Links ]

