










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkHidrobiológica
versão impressa ISSN 0188-8897
Hidrobiológica vol.10 no.1 Ciudad de México Jul. 2000
Notas
Abundancia y riqueza específica de aves playeras en la playa "El Conchalito", Baja California Sur, durante 1993 y 1995
Roberto Carmona1 y Cynthia Carmona 1,2
1 Universidad Autónoma de Baja California Sur. Depto de Biología Marina. Ciencias del Mar. A. P. 19-B. La Paz, B. C. S. 23080. México.
2 Centro Interdisciplinario de Ciencias Marinas. A. P. 592. La Paz, B. C. S. 23000. México.
Recibido: 7 de febrero de 1999
Aceptado: 22 de septiembre de 1999
Resumen
Entre 1993 y 1995 se realizó la construcción de dos unidades habitacionales en las adyacencias de la playa "El Conchalito", Ensenada de La Paz, Baja California Sur. Fueron comparadas la abundancia y la riqueza específica de aves playeras a partir de censos llevados a cabo en esta playa, de febrero de 1993 a enero de 1994 y de febrero de 1995 a enero de 1996. Para realizar la comparación se utilizaron pruebas de diferencia de proporciones (α=0.05), mismas que evidenciaron diferencias significativas. La abundancias de las especies más numerosas se comparó utilizando la diferencia porcentual promedio mensual, mediante diferencias de proporciones. Las pruebas mostraron una disminución significativa de las especies más abundantes en la playa (Limosa fedoa, Catoptrophorus semipalmatus, Limnodromus spp. y Charadrius wilsonia), las que no mostraron diferencias significativas fueron aquellas con utilización del área menor (Calidris mauri y Numenius americanus). Es probable que estas disminuciones se relacionen con el disturbio ocasionado por el desmonte y la construcción de la unidad habitacional antes mencionada.
Palabras clave: Aves playeras, abundancia, riqueza específica, Bahía de La Paz.
Abstract
Between 1993 and 1995, the construction of two housing units were built nearby "El Conchalito" beach, Ensenada de La Paz, Baja California Sur. Abundance and richness of shorebirds were censused from February, 1993 to January, 1994 and from February, 1995 to January, 1996, and compared with test of proportion (α=0.05). The results showed that the reductions in abundance and richness were significant. Species abundances were compared using the monthly average percentual difference, using a proportion test. Those species which presented signifficant reductions (Limosa fedoa, Catoptrophorus semipalmatus, Limnodromus spp. and Charadrius wilsonia) were the most abundant in the beach, while those which had no significant differences were those that were scarce (Calidris mauri and Numenius americanus). It is probable that population reductions are related with the disturbance caused by the buildong of a housing facility.
Key words: Shorebirds, abundance, specific richness, La Paz Bay.
Uno de los grupos predominantes entre las aves acuáticas migratorias es el de los playeros. En la ensenada de La Paz, Baja California Sur, México, se conforma por cuatro familias del orden Charadriiformes: Scolopaciae, Charadriidae, Haematopidae y Recurvirostridae (Carmona, 1995). Muchas de las especies de este grupo experimentan fuertes demandas energéticas asociadas con vuelos migratorios de gran alcance; razón por la que las poblaciones de aves playeras de sistemas costeros parecen asociarse a sitios con recursos alimenticios abundantes y predecibles (Skagen y Knopf, 1993).
Las aves migratorias se benefician por su habilidad para explotar recursos cíclicos en lugares a los que no se les puede dar uso continuo. Sin embargo, ésta misma habilidad puede ocasionar la dependencia de una secuencia específica de sitios eslabonados a lo largo de su ruta (Wolf, 1981; Erwin et al., 1986). Se mencionan cuatro aspectos básicos de las aves migratorias que tienen que ver con su conservación: i) las frágiles características de sus ciclos de vida, ii) su elevada concentración en pequeños sitios de migración e invernación, iii) la precisión de sus requerimientos espaciales y energéticos en la migración y iv) su competencia con el hombre (Myers et al., 1987). Los primeros tres aspectos son intrínsecos de las aves, sin embargo, en lo que respecta a su competencia con el hombre, el crecimiento poblacional humano ha traido como consecuencia que se hayan degradado muchos de los sitios necesarios en la ruta de las aves migratorias (Myers, 1983; Senner y Howe, 1984; Escofet et al., 1993).
Para saber cuáles de estos sitios son de importancia crítica para aves migratorias y residentes, se requiere del conocimiento de datos básicos sobre su variación numérica y específica a lo largo de menos de un año, continuando con información sobre su variación interanual (Page et al., 1979).
Existe controversia respecto a una probable disminución en las poblaciones de aves playeras en las últimas décadas (Paulson, 1993; Page & Gill, 1994; Rappole & McDonald, 1994); otros sugieren que existe poca evidencia de disminuciones en las abundancias recientes (Paulson, 1993). Por lo anterior, los estudios sobre variación interanual de abundancias se hacen impredecibles.
Se sabe que, en algunos casos las rutas migratorias de primavera y de otoño son diferentes, probablemente con la finalidad de tomar ventaja de distintas oportunidades de alimentación (Paulson, 1993; Carmona y Danemann, 1998). Éste podría ser el caso de la ensenada de La Paz, ya que la zona es utilizada por las aves playeras básicamente durante la migración de otoño (Carmona, 1995; Fernández et al., 1998; Carmona, 1997). En los márgenes de la ensenada se ubica la capital del Estado, e inmersa en la zona urbana se localiza en la playa "El Conchalito". Esta playa reviste importancia para las aves playeras, dado que es una zona de paso, invernación y alimentación de diferentes especies (Carmona, 1995; Carmona, 1997). Entre 1993 y 1995 la periferia de la zona sufrió importantes modificaciones debido a su urbanización, pues se construyeron dos unidades habitacionales adyacentes, las cuales albergan a poco más de 200 familias. Las construcciones han ocasionado una nueva presión para la fauna nativa, debido básicamente a su uso como área de recreación, a la acumulación de basura, y a la presencia de fauna doméstica.
En el trabajo presente se compara la abundancia y la riqueza específica de aves playeras en la playa "El Conchalito", durante 1993 (antes de la construcción) y 1995 (después de la misma).
El clima de la región es semidesértico seco y caluroso, con temperaturas mínimas de 2 a 8°C entre diciembre y febrero, y máximas de 40 a 43°C entre julio y agosto. La evaporación promedio anual es de 215 mm, sobrepasando a la precipitación, que es de apenas 200 mm anuales, con régimen de lluvia en verano (CETENAL, 1970).
La playa El Conchalito (Fig. 1), inmersa en la región urbana de la ciudad de La Paz, está ubicada en la parte sudoriental de la ensenada de La Paz (24° 08' 21" N; 110° 20' 55" W). Presenta una zona de inundación que va de los 60 a los 250 m, y tiene una longitud de aproximadamente 2 km (Carmona, 1995). El sustrato de la playa varía desde arenoso compacto con numerosos fragmentos de concha en su porción más norteña, hasta lodoso en su parte Sur. La playa se encuentra bordeada por el manglar "El Conchalito", constituído por tres especies de mangle: rojo (Rhizophora mangle), negro (Avicennia germinans) y blanco (Laguncularia racemosa) (Becerril, 1994).

Para determinar la abundancia de aves en esta playa, se realizaron censos quincenales de febrero de 1993 a enero de 1994, y semanales, de febrero de 1995 a enero de 1996. Se utilizaron binoculares (7 y 10 X) y telescopio (30-65 X), se contabilizó con los primeros, mientras que el telescopio se utilizó para corroborar identificaciones. Los conteos se realizaron caminando paralelamente a la línea de la costa, procurando perturbar lo menos posible a los grupos de aves que se encontraban alimentándose, la técnica empleada fue la de barrido (Palacios et al., 1991). Los censos se hicieron en niveles de bajamar de marea muerta, ya que las aves tiene espacio de alimentación disponible y a este nivel de marea se pueden llevar a cabo censos semanal y quincenalmente.
Para obtener las abundancias mensuales a contrastar (1993 vs. 1995) y dada la diferencia en el esfuerzo realizado, los datos se estandarizaron dividiendo la sumatoria mensual entre el numero de censos llevados a cabo en el mes correspondiente; en función a estos promedios se realizaron las comparaciones entre los dos años. La riqueza fue estimada directamente, es decir se registró el total de especies observadas por mes y a lo largo de todo el estudio. Para realizar pruebas de proporciones, se determinó la disminución porcentual promedio para cada mes, tanto de abundancia como de riqueza (Daniel, 1995).
Para determinar cuales especies fueron las representativas durante las dos temporadas, se analizó la abundancia porcentual acumulada, para así (ordenadas descendentemente), detectar los puntos donde se acumuló aproximadamente el 80% de la abundancia. Las especies así incluidas fueron comparadas para ambos años. Las comparaciones se realizaron determinando el porcentaje de disminución mensual promedio entre 1993 y 1995 (α=0.05; Daniel, 1995). La forma de contraste empleada no permitió que se consideraran meses en los que no se observaron aves, por lo que el número de muestra no fue homogéneo.
Se comparó la abundancia de aves playeras para 1993 y 1995 (tabla 1; figura 2), notándose una clara disminución para el segundo año, misma que fue corroborada estadísticamente (t=4.53, g.l.=11, p<0.01). Existen algunas diferencias temporales, de tal forma que las menores disminuciones se observaron en el verano, mientras que las más notables ocurrieron durante las épocas de movimiento de las aves (primavera y otoño), de éstas fue más marcada la diferencia de otoño.


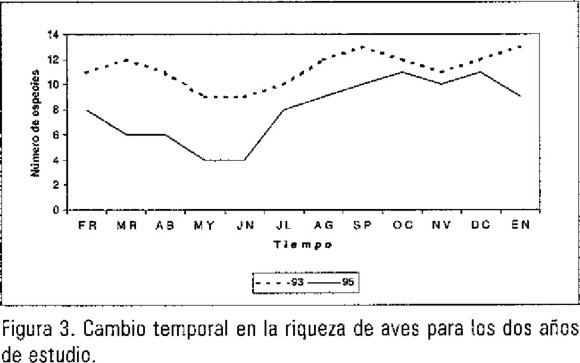
En lo que respecta a la riqueza específica (tabla 1; figura 3), se observó una disminución menos drástica que para la abundancia (t=2.29, g.l.=11, p=0.02). En contraste con la abundancia, la riqueza mostró mayores diferencias en el verano, seguida de la primavera y las mayores similitudes se observaron durante el otoño y el invierno. En 1993 se observaron 20 especies, mientras que para 1995 este número disminuyó a 16 (más una nueva), lo que indica una disminución de 20% (tabla 1); sin embargo, las cuatro especies involucradas apenas representaron el 6.1% de la abundancia total en 1993 (tabla 1).
Las especies incluidas en el 80% de abundancia acumulativa porcentual para 1995 fueron: Limosa fedoa, Limnodromus spp., Calidris mauri, Catoptrophurus semipalmatus, Charadrius wilsonia y Numenius americanus; estas mismas representan el 78.5% para 1993, razón por la que se consideró que su comparación permite un análisis adecuado de la comunidad.
Es de resaltar que Limosa fedoa es en la que se observó mayor diferencia, seguida de tres especies en las que también hubo cambios significativos (Catopthophorus semipalmatus, Limnodromus spp. y Charadrius wilsonia), en estos tres casos la diferencia mensual promedio se ubicó entre el 30 y el 40%. Por último, hubo dos especies en las que, pese a disminuir sus abundancias, no se detectaron cambios significativos (Numenius americanus y Calidris mauri).
Durante 1993 la playa se utilizó preferentemente durante la migración de otoño; patrón observado en otros puntos de la península (López, 1994; Carmona, 1995; Fernández et al., 1998; Carmona & Danemann, 1998). Durante 1995 las abundancias observadas en primavera y otoño fueron similares, por lo que el aparente efecto causado por las construcciones fue más notorio en otoño.
Al analizar la abundancia temporal de las especies más abundantes, se observó que las que presentaron la mayor disminución poblacional fueron, en orden descendente: Limosa fedoa (especie invernante, con fracciones poblacionales veraneantes), Catoptrophus semipalmatus (especie que hace uso del área como zona de paso e invernación, con individuos veraneantes), Limnodromus spp. (género que hace uso del área como zona de paso) y la única especie residente Charadrius wilsonia (existen grupos de paso); y las dos especies que no mostraron cambios en su abundancia fueron Calidris mauri y Numenius americanus (ambas especies utilizan el área como zona de paso e invernación). Existió un patrón general, en el cual cuanto más abundante fue una especie mayor fue su disminución, mismo que se refleja en los resultados de la prueba de hipótesis y en los niveles de significancia (tabla 2).

Una revisión de la literatura indica que L. fedoa era más numerosa en el siglo pasado, sin embargo, no se tienen datos que apoyen una declinación poblacional en los últimos años, al menos para la parte occidental de norteamérica (Paulson, 1993; Page & Gill, 1994). Howe et al. (1989) encontraron que Limnodromus griseus mostró decrementos estadísticamente significativos entre 1972 y 1983. N. americanus después de ser abundante en el siglo pasado, llegó a tener niveles poblacionales peligrosamente bajos; sin embargo, se cree que las poblaciones ya se han recuperado, sólo se sugiere una declinación poblacional para la porción Este de su área de distribución, y un incremento para la porción Oeste (Page & Gill, 1994). De las demás especies no se tienen reportados cambios poblacionales.
La disminución de la riqueza específica, involucró a cuatro especies, cuya abundancia conjunta fue menor al 7%. Es decir, probablemente no se detectaron debido a sus bajas densidades.
La diferencia en las abundancias tiene dos posibles explicaciones: (1) que en 1993 se haya presentado una abundancia anómalamente alta, o bien (2) a una efectiva disminución de las aves. En el segundo caso, la disminución podría deberse a un descenso poblacional general, o a un cambio en la utilización de los humedales regionales. Esta disminución podría relacionarse con el disturbio ocasionado por el desarrollo urbano, pues está comprobado el efecto negativo de la presencia humana para distintas especies de aves acuáticas (Klein et al., 1995). Considerando la bibliografía, es poco probable que 1993 haya sido anómalamente alto en la zona, pues con regularidad se ha efectuado un vuelo en el mes de enero a lo largo de la costa pacífica de la península, de estos datos es claro que 1993 no altera el patrón, al menos para cinco lagunas costeras de la región (Page y Palacios, 1993; Page et al., 1997), por lo que es factible suponer que en 1993, la Ensenada de La Paz no presentó abundancias particularmente altas. Dadas las escasas posibilidades de un descenso poblacional global, es más probable suponer un cambio en la utilización de la playa, el cual a su vez se puede relacionar con el disturbio del lugar, causado por el desmonte y la edificación de la unidad habitacional a escasos 200 metros de la playa, con lo que podría disminuir la calidad del hábitat. Lo anterior está apoyado por el hecho de que las especies que presentaron una disminución significativa, fueron aquellas que se encuentran más comúnmente en la playa. Pfister et al., (1992), encontraron disminuciones de aves playeras entre 1972 y 1989 a causa del disturbio humano, y reportan efectos diferenciales, dependiendo de la especie.
En suma, entre 1993 y 1995 existió una disminución significativa tanto en la abundancia como en la riqueza de aves playeras en la Playa "El Conchalito", esta disminución coincidió con la edificación de una unidad habitacional aledaña a la playa, por lo que es probable que al menos parte de esta reducción se relacione con una disminución en la calidad del hábitat, ocasionada por el desarrollo urbano.
Agradecemos a las personas tanto de la UABCS como del CICIMAR que nos acompañaron a los censos. A Georgina Brabata y Andrea Cuéllar por su apoyo en la edición del manuscrito. A Juan Guzmán por su apoyo logístico.
Literatura citada
Becerril, F., 1994. Reparto de los recursos temporal, espacial y trófico, por parte de los ardéidos anidantes en el manglar El Conchalito, Baja California Sur, México, durante las temporadas reproductivas de 1992 y 1993. Tesis de licenciatura. U. A. B. C. S. La Paz, B. C. S., México. 73 p. [ Links ]
Carmona, R., 1995. Distribución temporal de aves acuáticas en la playa El Conchalito, Ensenada de La Paz, B. C. S., México. Invest. Marinas, CICIMAR 10 (1-2): 1-22. [ Links ]
Carmona, R. y G. Danemann, 1998. Distribución espaciotemporal de aves en la salina de Guerrero Negro, Baja California Sur, México. Ciencias Marinas 24: 389-408. [ Links ]
Carmona, C., 1997. Relación entre la abundancia de aves playeras y organismos bentónicos en la playa "El Conchalito", Ensenada de La Paz, B. C. S., durante un ciclo anual. Tesis de Maestría en Ciencias. CICIMAR-IPN. La Paz, B. C. S., México. 69 p. [ Links ]
CETENAL, 1970. Carta Climática, La Paz 12R-VIII. Dirección de planeación. La Paz, B. C. S. México. [ Links ]
Daniel, W., 1995. Bioestadística: Base para el análisis de las ciencias de la salud. Limusa. México. 667 p. [ Links ]
Erwin, R. M., M. Coulter y H. Cogswll, 1996. The use of natural vs man-modified wetlands by shorebirds and waterbirds. Colonial Waterbirds 9 (2): 137-138. [ Links ]
Escofet, A., I. Espejel, J. L. Ferman, L. Gómez-Morin Fuente y G. Torres-Moye, 1993. El manejo de fragmentos en la zona costera. 182-193 pp. En: Salazar-Vallejo, S. y N. E. González (eds.) Biodiversidad Marina y Costera de México. CONABIO y CIORO, México. 865 p. [ Links ]
Fernández, G., R. Carmona y H. De la Cueva, 1998. Abundance and seasonal variation of western sandpipers (Calidris mauri) in Baja California Sur, México. Southwestern Naturalist 43 (1): 57-61. [ Links ]
Howe, M. A., P. H. Geissler y B. A. Harrington, 1989. Population tredns of North American Shorebirds based on the International Shorebird Survey. Biology Conservation 49: 185-199. [ Links ]
Klein, M. L., S. R. Humphrey y H. Franklin, 1995. Effects of ecotourism on distribution of waterbirds in a wildlife refuge. Conservation Biology 9: 1454-1465. [ Links ]
López, E., 1994. Infauna de dos playas arenosas adyacentes a Punta Cabras (B. C., México) y su relación con el "Corredor migratorio del Pacífico". Tesis de Licenciatura. UABC. México. 107 pp. [ Links ]
Myers, J. P., 1983 Conservation of migrating shorebirds: Staging areas, geographic bottlenecks, and regional movements. American Birds 37: 23-25. [ Links ]
Myers, J. P., R. I. G. Morrison, P. Z. Antas, B. A. Harrington, T. E. Lovejoy, M. Sallaberry, S. E. Senner y A. Tarak, 1987. Conservation strategy for migratory species. American Scientist 75: 19-26. [ Links ]
Page, G., L. Stenzel y C. Wolfe, 1979. Aspects of the occurrence of shorebirds on a central California estuary. 15-32 pp. en: Pitelxa, F. (ed.) Shorebirds in marine environments. Studies in Avian Biology 2. Cooper Omithological Society. E. U. A. 261p. [ Links ]
Page, G. W. y R. E. Gill, 1994. Shorebirds in western North America: late 1800s to late 1900s. Studies in Avian Biology 15: 285-309. [ Links ]
Page, G. W. y E. Palacios, 1993. Winter shorebird numbers in wet-lands along the coast of Baja California. PRBO and CICESE. Report 16 p. [ Links ]
Page, G. W., E. Palacios, L. Alfaro, S. González, L. E. Stenzel y M. Jungers, 1997. Numbers of wintering shorebirds in coastal wetlands of Baja California, México. J. Field Ornithol. 68: 562-574. [ Links ]
Palacios, E., A. Escofet y D. H. Loya-Salinas, 1991. El Estero de Punta Banda, B. C., México como eslabón del "Corredor del Pacífico": abundancia de aves playeras. Ciencias Marinas 17: 109-130. [ Links ]
Paulson, D., 1993. Shorebirds of the Pacific Northwest. University of Washington Press. E. U. A. 406 p. [ Links ]
Pfister, Ch., B. A. Harrington y M. Lavine, 1992. The impact of human disturbance on shorebirds at a migration staging area. Biological Conservation 60(2): 115-126. [ Links ]
Rappole, J. H. y M. V. Mc Donald, 1994. Cause and effect in population declines of migratiory birds. Auk. 111 (3): 652-660. [ Links ]
Senner, S. E. y M. A. Howe, 1984. Conservation of neartic shorebirds. Behavior of Marine Organisms 5: 379-421. [ Links ]
Skagen, S. K. y F. L. Knopf, 1993. Toward conservation of midcontinental shorebirds migrations. Conservation Biology 7 (3): 533-541. [ Links ]
Wolf, W. J., 1981. Management of estuaries in the Netherlands with respect to bird populations. Estuaries 4 (3): 265-266. [ Links ]

