











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Se considera que casi una tercera parte de los bosques a nivel mundial son primarios, es decir, ecosistemas con especies nativas, los cuales se regeneran de manera natural, en los que la actividad humana es apenas perceptible y donde los procesos ecológicos no sufren perturbaciones destacables (Food and Agriculture Organization of the United Nations [FAO], 2020). En el noreste de México se desarrollan bosques mixtos que en su mayoría son dominados por especies de los géneros Pinus y Quercus. Sin embargo, también se distribuyen especies arbóreas de la región biogeográfica Neártica, que crecen en condiciones climáticas extremas (baja humedad y alta temperatura) y son en algunos casos endémicas para una región (Treviño, 2017). Los bosques conformados por este tipo de especies son considerados relictos, ya que muestran una estructura única, con predominio de árboles maduros y bajo grado de perturbación; conservan una alta biodiversidad; y tienen la posibilidad de brindar servicios ambientales (Cruz et al., 2014).
El bosque donde se desarrolla Picea martinezii T.F. Patterson (una de las 38 especies de este género que existen a nivel mundial) es un ejemplo de bosque maduro, con escasa perturbación. Picea chihuahuana Martínez, P. mexicana Martínez y Picea martinezii son las únicas especies del género que se distribuyen en México (Gernandt & Pérez-de la Rosa, 2014). La madera de Picea es considerada de alto valor comercial en muchas partes del mundo; sin embargo, en México, debido al bajo número de individuos y a la reducción en el tamaño del área de distribución de sus poblaciones, las tres especies se encuentran en peligro de extinción, por lo que no está permitido su aprovechamiento maderable (Diario Oficial de la Federación [DOF], 2010; Thomas & Farjon, 2013).
En el caso particular de Picea martinezii, su área de distribución se restringe a algunas localidades con bosques mixtos de coníferas en el noreste de México, en el estado de Nuevo León, donde frecuentemente coexisten con especies arbóreas de los géneros Abies, Carpinus, Carya, Cornus, Ilex, Pinus, Pseudotsuga, Quercus, Taxus, y Tilia, entre otros (Comisión Nacional para el Conocimiento y Uso de la Biodiversidad-Comisión Nacional de Áreas Naturales Protegidas [Conabio-Conanp], 2009).
Al no ser posible el aprovechamiento forestal de Picea, los ecosistemas forestales donde crece se mantienen con bajo grado de perturbación, por lo que son de gran importancia ecológica debido a que proveen de servicios ecosistémicos a las poblaciones humanas aledañas. Por lo antes mencionado, surge la necesidad de estudiar distintos aspectos ecológicos de las especies de este género para aportar información básica que pueda ser de utilidad en la toma de decisiones sobre el manejo y conservación de estos ecosistemas. Hasta ahora se han realizado varios estudios sobre Picea martinezii y las comunidades forestales donde se distribuye, destacando los trabajos sobre diversidad genética (Ledig et al., 2000a), distribución de sus poblaciones (Ledig et al., 2000b), relación genética entre las especies (Ledig et al., 2004), efectos del cambio climático (Ledig, 2012), indicadores reproductivos (Flores-López et al., 2012) y propuestas de conservación (Mendoza-Maya et al., 2015). Sin embargo, es necesario realizar investigaciones sobre varios aspectos que aún no se han explorado, como la dinámica de sus poblaciones y sus características estructurales (Wehenkel et al., 2022).
La estructura de un ecosistema se relaciona con la forma en que los atributos de los árboles se distribuyen dentro de los mismos (Larson & Funk, 2016) y se puede caracterizar a diferentes niveles de estudio, como el horizontal, el florístico y el dimensional (Aguirre, 2002). Para cuantificar la estructura arbórea, es común utilizar índices que estiman sus patrones de distribución espacial y que están basados en relaciones de vecindad, como el índice de agregación espacial R de Clark & Evans (1954), que sirve para analizar la estructura horizontal y describir la distribución espacial del arbolado (Vandekerkhove et al., 2018); el índice de mezcla de especies, utilizado para analizar la estructura florística; y los índices de diferenciación y dominancia, que se usa para analizar la estructura dimensional (Gadow et al., 2012; Ghalandarayeshi et al., 2017).
El objetivo de este trabajo fue caracterizar y comparar la estructura arbórea a nivel horizontal, florístico y dimensional en cuatro localidades con ecosistemas forestales en donde se presenta Picea martinezii en el noreste de México. Considerando la similitud en las condiciones ambientales que se presentan en las cuatro comunidades estudiadas, se plantea la hipótesis de que la estructura arbórea y el estado de conservación también pueden ser semejantes.
Materiales y métodos
La investigación se realizó en cuatro localidades con Picea martinezii en el estado de Nuevo León, México: El Butano (25° 10.585' N, 100° 7.494' O), Agua de Alardín (24° 2.505' N, 99° 43.882' O), El Bosque (23° 53.414' N, 99° 46.438' O) y La Tinaja (23° 53.376' N, 99° 47.525' O) (Figura 1).
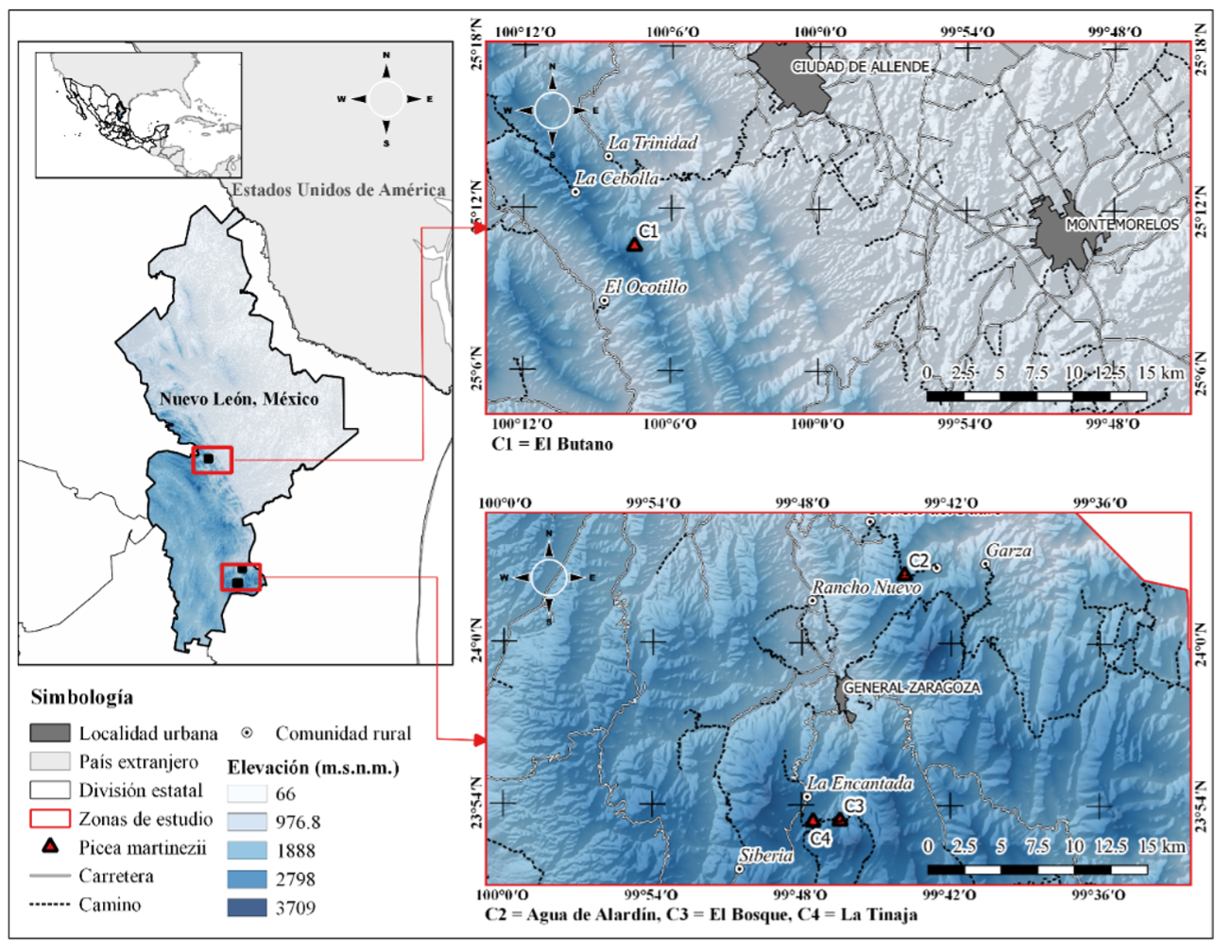
En las cuatro localidades se presentan condiciones ambientales similares: se distribuyen en cañadas, con exposición norte, en la Sierra Madre Oriental (SMO), en un gradiente altitudinal que va de 2180 m a 2600 m (registrado en los sitios establecidos para este estudio). El clima que predomina es tipo C (w), que corresponde a templado subhúmedo (Instituto Nacional de Estadística y Geografía [INEGI], 2008), con precipitación media anual de 700 mm - 800 mm (INEGI, 2006) y temperatura media anual entre 12 °C y 16 °C (INEGI, 2007). El suelo tipo leptosol predomina en todas las comunidades (INEGI, 2013). Las condiciones ambientales mencionadas favorecen el establecimiento y desarrollo de especies arbóreas típicas del bosque mesófilo de montaña de los géneros Abies, Carya, Cornus, Ilex, Picea, Pinus, Quercus, Taxus, entre otras (Rzedowski, 2006), incluyendo a Picea martinezii, y otras coníferas consideradas en alguna categoría de riesgo como Abies vejarii Martínez, Taxus globosa Schltdl. y Pseudotsuga menziesii (Mirb.) Franco (Treviño, 2017). Entre las localidades analizadas, sólo El Butano se encuentra dentro de un área natural protegida.
Los datos fueron obtenidos en cuatro unidades de muestreo permanentes de 2500 m2 (50 m X 50 m) establecidas en el año 2016 (una unidad por localidad). Según el método propuesto por Corral-Rivas et al. (2009), cada unidad se dividió en cuatro cuadros de 625 m2 (25 m x 25 m), y en cada uno de ellos se realizó el inventario de todos los árboles con diámetro a 1.30 m (Dn) mayor de 7.5 cm. Se registraron los datos del género, especie, Dn, altura total y distancia (metros) y azimut (grados), con respecto al centro de la unidad de muestreo.
Para analizar la estructura arbórea a nivel horizontal, florístico y dimensional se emplearon los índices que se indican en la Tabla 1. Los valores se calcularon de manera general (tomando en cuenta todas las especies) y por grupo (considerando los géneros más representativos). Con excepción del índice de Clark & Evans (1954), los índices empleados se basan en el método de los cinco árboles, que consideran un árbol de referencia y sus cuatro vecinos más cercanos (Gadow et al., 2012).
Tabla 1 Índices empleados para caracterizar la estructura arbórea.
| Índice | Símbolos y definiciones | Clasificación | |
| Agregación
(Clark & Evans, 1954) |
N | Número de medidas de distancia tomadas |
R entre 0.00-0.49 (Agrupación)
R entre 0.50-1.49 (Aleatoriedad) R entre 1.50-2.149 (Uniformidad) |
| r | Cada distancia tomada | ||
| ρ | Número de individuos | ||
| Σr | Suma de las medidas de distancia | ||
|
|
Distancia media | ||
|
|
Distancia media esperada | ||
| Mezcla de especies
(Gadow et al., 2012) |
n | Número de árboles vecinos considerados (cuatro en este caso) |
Mi = 0.00 (Mezcla nula)
Mi = 0.25 (Mezcla baja) Mi = 0.50 (Mezcla intermedia) Mi = 0.75 (Mezcla alta) Mi = 1.00 (Mezcla muy alta) |
| vj | Toma uno de dos posibles valores: en caso de que la especie j sea diferente a la especie de referencia i, el valor asignado será de 1, y tomará un valor de 0 en caso contrario. | ||
| Diferenciación dimensional
(Gadow et al., 2012) |
n | Número de árboles vecinos considerados (cuatro en este caso) |
Ti = 0.00 (Diferenciación nula)
Ti = 0.25 (Diferenciación baja) Ti = 0.50 (Diferenciación intermedia) Ti = 0.75 (Diferenciación alta) Ti = 1.00 (Diferenciación muy alta) |
| min(di,dj) | Diámetro menor del árbol de referencia i o el del árbol vecino j | ||
| max(di,dj) | Diámetro máximo del árbol de referencia i o el del árbol vecino j | ||
| Dominancia dimensional
(Aguirre et al., 2003) |
n | Número de árboles vecinos considerados (cuatro en este caso) |
Ui = 0.00 (Dominancia nula)
Ui = 0.25 (Dominancia baja) Ui = 0.50 (Dominancia intermedia) Ui = 0.75 (Dominancia alta) Ui = 1.00 (Dominancia muy alta) |
| vj | Toma uno de dos posibles valores: en caso de que la especie j sea diferente a la especie de referencia i, el valor asignado será de 1, y tomará un valor de 0 en caso contrario. | ||
Fuente: Elaboración propia.
El índice R propuesto por Clark & Evans (1954), empleado para caracterizar la estructura horizontal, permite conocer la manera y el grado de distribución espacial de los individuos. El índice de mezcla (Mi) empleado para caracterizar la estructura florística, define la proporción de árboles vecinos que pertenecen a especies diferentes, con respecto a un árbol de referencia i (Rubio-Camacho et al., 2017).
Los índices de diferenciación (Ti) y dominancia (Ui), empleados para caracterizar la estructura dimensional, describen la diferenciación por dimensiones, definida como una medida de la proximidad espacial existente entre árboles de diferentes medidas dentro de un rodal. Para su determinación se puede considerar cualquier variable que represente el tamaño de un árbol (García-García et al., 2022; Silva-González et al., 2022); para efectos de esta investigación se empleó el Dn.
Para calcular los índices y realizar los análisis estadísticos correspondientes, se utilizó el software libre R® versión 3.3.3 (R Core Team, 2017), junto con el script TreeDiversityIndices.R (Pommerening et al., 2011) y la caja de herramientas llamada SPATSTAT® versión 1.54-0 (Baddeley et al., 2017). Los análisis estadísticos se realizaron según lo establecido por Wackerly et al. (2008) y Dytham (2011). Primero, se efectuaron pruebas para verificar si los datos obtenidos a partir de los índices cumplían con los supuestos de normalidad y homocedasticidad, donde se encontró que sólo los índices 𝑅 y 𝑇𝑖 cumplían con estos supuestos. En ambos casos se empleó un análisis de varianza (Anova) para probar diferencias entre comunidades, el resto fueron sometidos a una prueba de Kruskal-Wallis. Finalmente, se realizaron pruebas de comparación múltiple de Tukey para identificar diferencias estadísticas por pares entre comunidades.
Resultados
Se registraron 19 especies arbóreas representadas por 12 géneros, las cuales fueron consideradas para analizar y comparar la estructura general del bosque. De los géneros, los que presentaron mayor densidad y dimensiones y que se encontraron en todas las comunidades fueron Abies, Picea, Quercus y Taxus (Tabla 2), por lo que estos fueron empleados en la comparación por grupos para analizar su comportamiento estructural particular.
Tabla 2 Características dasométricas de los individuos arbóreos registrados.
| No. | Especie | Densidad (N ha-1) | Dn (cm) | Altura (m) | |||||||||
| But | Alar | Bos | Tin | But | Alar | Bos | Tin | But | Alar | Bos | Tin | ||
| 1 | Picea martinezii T.F.Patt. | 72 | 36 | 52 | 128 | 21.5 | 38.8 | 13.5 | 19.9 | 14.7 | 19.8 | 11.7 | 16.0 |
| 2 | Abies vejarii Martínez | - | 84 | 224 | 64 | - | 22.2 | 22.3 | 25.7 | - | 12.3 | 16.9 | 23.3 |
| 3 | Abies durangensis var.Coahuilensis (I.M.Johnst.) Martínez | 216 | - | - | - | 20.6 | - | - | - | 16.2 | - | - | - |
| 4 | Quercus affinis Scheidw. | - | 36 | 124 | - | - | 34.6 | 20.8 | - | - | 16.2 | 13.0 | - |
| 5 | Quercus glaucoides M.Martens & Galeotti | 16 | - | - | - | 12.1 | - | - | - | 11.8 | - | - | - |
| 6 | Quercus greggii (A.DC.) Trel. | 8 | - | - | - | 10.7 | - | - | - | 6.7 | - | - | - |
| 7 | Quercus mexicana Bonpl. | - | 4 | - | - | - | 13.3 | - | - | - | 8.5 | - | - |
| 8 | Quercus sartorii Liebm. | 64 | 40 | - | - | 22.2 | 34.2 | - | - | 13.0 | 13.7 | - | - |
| 9 | Quercus laurina M.Martens & Galeotti | - | - | 100 | 68 | - | - | 23.2 | 37.6 | - | - | 13.0 | 17.5 |
| 10 | Quercus sideroxyla Bonpl. | - | - | 36 | - | - | - | 38.6 | - | - | - | 13.6 | - |
| 11 | Taxus globosa Schltdl. | 124 | 92 | 56 | 48 | 19.2 | 16.1 | 14.4 | 19.0 | 6.7 | 7.1 | 6.2 | 11.6 |
| 12 | Ilex rubra S.Watson | 128 | - | - | - | 23.2 | - | - | 14.6 | - | - | - | |
| 13 | Carpinus caroliniana Walter | 44 | 12 | 8 | - | 14.1 | 22.2 | 20.1 | - | 9.0 | 14.0 | 12.9 | - |
| 14 | Sambucus nigra Lour. | 4 | - | - | - | 17.1 | - | - | - | 8.4 | - | - | - |
| 15 | Cornus florida L. | - | 76 | - | - | - | 22.6 | - | - | - | 13.1 | - | - |
| 16 | Tilia americana L. | - | 44 | - | - | - | 17.5 | - | - | - | 5.2 | - | - |
| 17 | Arbutus xalapensis Kunth | - | - | 4 | - | - | - | 13.0 | - | - | - | 10.1 | - |
| 18 | Pinus ayacahuite Ehrenb. ex Schltdl. | - | - | 4 | 44 | - | - | 17.6 | 28.6 | - | - | 14.5 | 21.0 |
| 19 | Pseudotsuga menziesii (Mirb.) Franco | - | - | 8 | 36 | - | - | 18.3 | 17.9 | - | - | 16.7 | 17.8 |
Nota. But = El Butano; Alar = Agua de Alardín; Bos = El Bosque; Tin = La Tinaja; Dn = Diámetro normal a la altura de 1.3 m.
Fuente: Elaboración propia.
En el análisis general (Tabla 3), se encontró que a nivel horizontal la distribución espacial de las especies arbóreas fue de tipo aleatoria, debido a que los valores de R fueron cercanos a 1 y no presentaron diferencias estadísticas significativas (p > 0.05). Esto implica que en las cuatro comunidades las especies arbóreas no tienden a agruparse ni a presentar un arreglo uniforme. A nivel florístico se presentó un alto grado de mezcla en la mayoría de los casos (Mi = 0.75), y sólo hubo diferencias significativas (p < 0.05) en la comunidad de El Bosque (Mi = 0.50), la cual fue clasificada como intermedia. En la estructura dimensional, los valores de los índices de diferenciación y dominancia se clasificaron como intermedios debido a que los valores de Ti y Ui fueron cercanos o iguales a 0.50, sin presentar diferencias estadísticas significativas entre comunidades (p > 0.05).
Tabla 3 Comparación de índices entre comunidades (análisis general).
| Índice | But | Alar | Bos | Tin | p-valor |
| R (M) | 1.05 a | 0.88 a | 0.87 a | 0.81 a | 0.318 |
| Mi (Mdn) | 0.75 a | 0.75 a | 0.50 b | 0.75 a | 0.000 |
| Ti (M) | 0.37 a | 0.42 a | 0.41 a | 0.36 a | 0.169 |
| Ui (Mdn) | 0.50 a | 0.50 a | 0.50 a | 0.50 a | 0.862 |
Nota. But = El Butano; Alar = Agua de Alardín; Bos = El Bosque; Tin = La Tinaja; M = valores promedio comparados con Anova; Mdn = valores de medianas comparados con Kruskal-Wallis. Valores en fila con diferente literal indican diferencias según prueba post hoc Tukey (p < 0.05).
Fuente: Elaboración propia.
En el análisis por géneros representativos (Abies, Picea, Quercus y Taxus), el índice R, al igual que el análisis general, indicó valores cercanos a 1 y no hubo diferencias estadísticamente significativas (p > 0.05) entre comunidades (Tabla 4), lo que muestra que en las cuatro comunidades los géneros estudiados se distribuyen de forma aleatoria, tal como se ilustra en la Figura 2. La mezcla de especies indicó valores de Mi > 0.50 en la mayoría de los casos, con diferencias estadísticas en Abies, Picea y Quercus (p < 0.05) sólo en una comunidad (Tabla 4). Esto indica que los cuatro géneros tienden a coexistir en un grado que va de intermedio a alto y muy alto, siendo Picea el más común (Figura 2).
Tabla 4 Comparación de índices entre comunidades (análisis por grupo).
| Índice | Género | But | Alar | Bos | Tin | p-valor |
| R (M) | Picea | 1.18 a | 0.85 a | 1.33 a | 1.06 a | 0.686 |
| Abies | 1.24 a | 0.89 a | 0.90 a | 1.31 a | 0.604 | |
| Taxus | 1.35 a | 1.24 a | 1.17 a | 1.61 a | 0.752 | |
| Quercus | 1.08 a | 0.98 a | 0.94 a | 1.20 a | 0.806 | |
| Mi (Mdn) | Picea | 1.00 a | 1.00 a | 0.87 a | 0.50 b | 0.000 |
| Abies | 0.75 ab | 0.37 a | 0.50 a | 1.00 b | 0.000 | |
| Taxus | 0.75 a | 0.75 a | 1.00 a | 0.62 a | 0.119 | |
| Quercus | 0.75 a | 0.75 a | 0.25 b | 1.00 a | 0.000 | |
| Ti (M) | Picea | 0.44 ab | 0.52 a | 0.45 ab | 0.31 b | 0.030 |
| Abies | 0.33 a | 0.34 a | 0.40 a | 0.31 a | 0.138 | |
| Taxus | 0.38 a | 0.35 a | 0.37 a | 0.50 a | 0.323 | |
| Quercus | 0.43 a | 0.49 a | 0.30 a | 0.40 a | 0.485 | |
| Ui (Mdn) | Picea | 0.37 a | 0.75 a | 0.25 a | 0.25 a | 0.145 |
| Abies | 0.75 a | 0.50 a | 0.50 a | 0.62 a | 0.701 | |
| Taxus | 0.50 a | 0.25 a | 0.25 a | 0.50 a | 0.219 | |
| Quercus | 0.50 a | 0.75 a | 0.50 a | 1.00 a | 0.069 |
Nota. But = El Butano; Alar = Agua de Alardín; Bos = El Bosque; Tin = La Tinaja; M = valores promedio comparados con Anova; Mdn = valores de medianas comparados mediante Kruskal-Wallis. Valores en fila con diferente literal indican diferencias según la prueba post hoc de Tukey (p < 0.05).
Fuente: Elaboración propia.
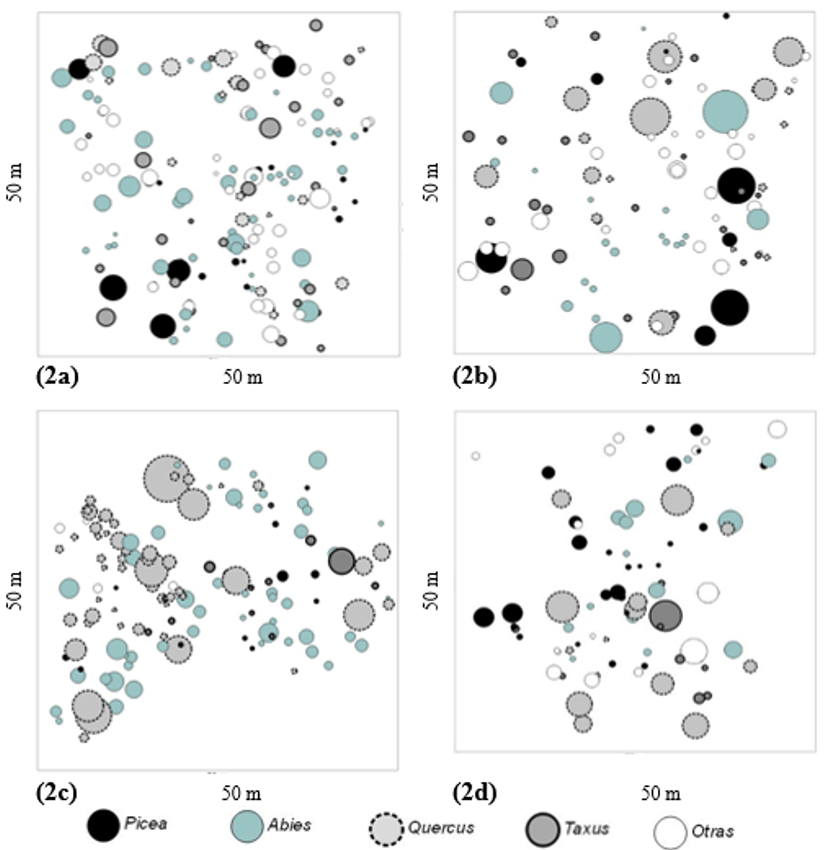
Fuente: Elaboración propia.
Figura 2 Distribución espacial y dimensional de todos los géneros arbóreos registrados en las cuatro comunidades estudiadas. (2a) El Butano, (2b) Agua de Alardín, (2c) El Bosque, (2d) La Tinaja. Los círculos representan los árboles de acuerdo con su respectivo diámetro normal a la altura de 1.3 m.
El índice de diferenciación dimensional (Ti) presentó valores cercanos a 0.50 en los cuatro géneros, con diferencias estadísticas significativas sólo para Picea entre las comunidades Agua de Alardín y La Tinaja. Sin embargo, sus valores se clasifican en el mismo grado de diferenciación (Tabla 4), con lo que se asume que por lo general la mitad de los árboles que los rodean (dos individuos) son de diferente tamaño (Figura 2).
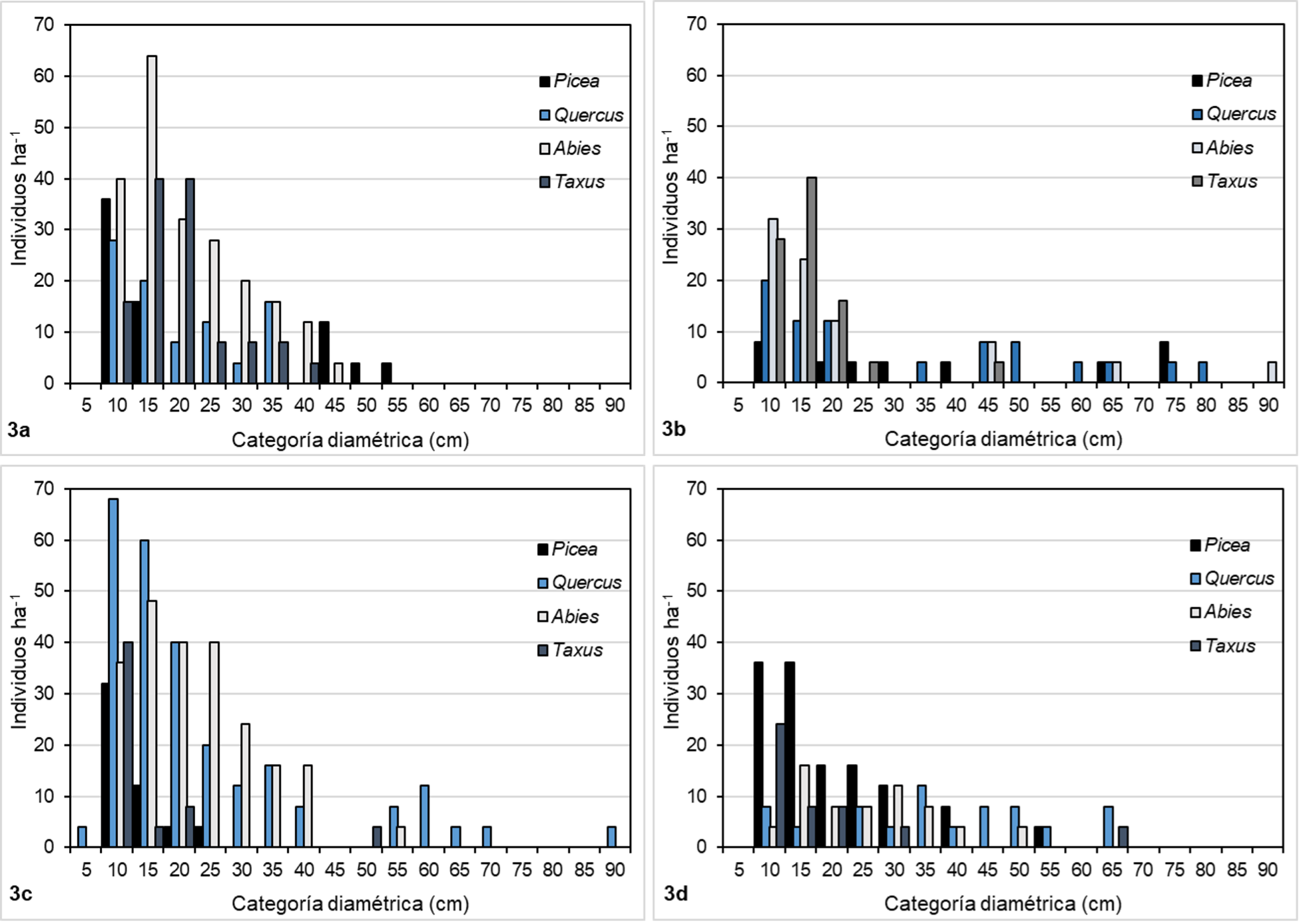
Por su parte, el índice de dominancia dimensional en diámetro (Ui) mostró que, de los cuatro géneros representativos, Picea y Taxus fueron los menos dominantes debido a que sus valores de Ui se clasificaron como bajos, intermedios y altos, mientras que Abies y Quercus fueron los más dominantes, ya que sus valores de Ui se clasificaron entre intermedios, altos y muy altos, sin presentar diferencias importantes entre comunidades (Tabla 4). Este patrón se puede apreciar mejor en la Figura 3, donde se observa que Abies y Quercus generalmente están bien representados en categorías diamétricas de tamaño considerable (de 25 cm a 40 cm), lo que aumenta su dominancia; mientras que Picea y Taxus están mejor representados en las primeras categorías, principalmente (5 cm a 20 cm), lo que los convierte en los menos dominantes.
Discusión
La distribución espacial de la estructura horizontal mostró que Picea martinezii y otras especies arbóreas que coexisten tienden a distribuirse de manera aleatoria. Este tipo de distribución es el resultado de la respuesta de los árboles a los procesos naturales que se llevan a cabo dentro de las comunidades, tales como la dispersión y el establecimiento de los individuos bajo condiciones lumínicas específicas (Götzenberger et al., 2012; Yan et al., 2018), que en el presente estudio ocurrieron en las especies más representativas: Abies spp., Picea martinezii, Quercus spp. y Taxus globosa. Aunado a lo anterior, se ha demostrado que los patrones de distribución espacial tienden a ser de forma agrupada cuando los individuos son jóvenes (plántulas o regeneración) y se convierten en aleatorios cuando las plantas llegan a edades intermedias y adultas, debido a que en estas etapas generalmente se desarrollan en los claros que se forman al azar bajo el dosel (Omelko et al., 2018).
Es posible que el patrón mencionado en el párrafo previo se esté presentando en las comunidades forestales estudiadas, pues se asume que los árboles se encuentran en edades intermedias y adultas. Como no se registraron individuos < 7.5 cm de diámetro, considerados como parte de la regeneración, la distribución espacial de las especies es aleatoria. Se considera que este tipo de distribución es característica de las comunidades forestales estudiadas y, además, se sugiere que en dichas comunidades no se han presentado disturbios antropogénicos fuertes recientemente (por ejemplo, aprovechamientos maderables intensivos), con ello se deduce que este tipo de distribución es característica de bosques de clima templado, donde habitan otras especies de Picea en México, así como en otros bosques mixtos de coníferas sin aprovechamiento forestal (Qindi et al., 2012; Wehenkel et al., 2015).
En el análisis general de la estructura florística, el alto grado de mezcla que presentaron las especies arbóreas en la mayoría de las comunidades, con excepción de El Bosque, sugieren que existe gran acumulación de casos donde tres de los cuatro árboles que rodean al de referencia son de especies diferentes. Esto se debe a que una gran proporción de especies presentan baja densidad y solo en una minoría la densidad es alta, ya que, según Juárez-Sánchez et al. (2014), existe relación entre la densidad de individuos y el grado de mezcla. Es decir, a medida que la densidad de un alto número de especies disminuye, el índice de mezcla aumenta. En el caso particular de la comunidad El Bosque, los resultados sugieren que la mitad de las especies presenta baja densidad y la otra mitad presenta densidad alta, por esta razón el grado de mezcla se clasificó como intermedio. Esta relación fue encontrada también en el análisis por grupo, donde los géneros Picea y Taxus en gran parte de las comunidades presentaron el mayor grado de mezcla, lo que coincide con el bajo número de árboles que se presentan.
En el caso particular de Picea martinezii, la baja densidad se atribuye a los problemas genéticos que enfrenta (Flores-López et al., 2012; Ledig et al., 2000a). A pesar de esta situación, se considera que la estructura florística de las comunidades donde habita la especie es estable e incluso puede ser mejor a la que presentan otras comunidades forestales de clima templado en México, donde los grados de mezcla tienden a ser generalmente intermedios (Juárez-Sánchez et al., 2014; Rubio-Camacho et al., 2017). Es probable que un alto grado de mezcla, como el encontrado en la mayoría de las comunidades estudiadas, favorezca la competencia entre individuos de diferentes especies, lo cual se ha demostrado tiende a ser menos fuerte en comunidades forestales donde habitan especies del género Picea y otras especies arbóreas de clima templado-frío (Cavard et al., 2011; Erfanifard et al., 2019; Torimaru et al., 2013). Por lo tanto, el establecimiento y crecimiento de las especies arbóreas que coexisten en estas comunidades es más propicio.
Las condiciones favorables de competencia entre árboles de diferente especie se ven reflejadas en los resultados obtenidos de la estructura dimensional, donde la diferenciación intermedia observada en el análisis general, y por grupo, sugiere que en todas las comunidades las especies arbóreas presentan distribución equilibrada en el tamaño de sus diámetros. De acuerdo con Ghalandarayeshi et al. (2017), el grado de diferenciación está relacionado con las dimensiones de los árboles y tiende a incrementar a medida que aumentan de tamaño, como resultado de la competencia, las condiciones del micrositio y la composición genética. De todo esto se deduce que la diferenciación intermedia que presentan las cuatro comunidades, en términos de dimensiones diamétricas, indica un estado de competencia equilibrado que obedece a condiciones de micrositio y composición genética similares. En adición, la dominancia intermedia encontrada también en el análisis general recalca el estado de competencia que presentan las comunidades, el cual, aunque no es tan elevado, propicia la dominancia en diámetro de algunas especies sobre otras.
Según Lutz et al. (2012), la dominancia está determinada por los géneros más representativos de cada comunidad. En este caso, el análisis por grupos indicó que, de los cuatro géneros más representativos presentes en las cuatro comunidades, Abies y Quercus y resultaron ser los más dominantes en diámetro, mientras que Picea y Taxus resultaron tener menor dominancia. En los ecosistemas templados del noreste de México se ha demostrado que Abies y Quercus, cuando se encuentran asociados, tienden a ser dominantes y se distinguen de los demás por presentar mayor dimensión (González et al., 2018). Para los individuos de Taxus globosa, el valor de dominancia menor es característico de este tipo de vegetación, debido a que suelen presentar dimensiones menores y casi siempre se encuentran en los estratos inferiores y medios de dosel en bosques montanos (Thomas, 2013). Es frecuente que los individuos de Picea martinezii sean dominantes, considerando las dimensiones que pueden llegar a alcanzar (Conabio-Conanp, 2009). Sin embargo, presentan baja densidad, lo cual, como ya se había mencionado, se atribuye a problemas genéticos que enfrenta la especie. Lo anterior está ocasionando la producción de semillas inviables (óvulos abortados, rudimentarios y semillas vacías), han estado mermando la incorporación de nuevos individuos de esta especie a pesar de poseer múltiples ciclos reproductivos y ser de larga vida, por lo que ha soportado estas presiones, permitiendo que sobrevivan durante muchos años (Flores-López et al., 2012; Ledig et al., 2000a).
Se considera que la reducción en la incorporación de nuevos individuos de Picea martinezii ha estado generando condiciones favorables para el establecimiento de especies como las del género Quercus, las cuales se ha demostrado que se establecen mejor en ecosistemas forestales que presentan algún tipo de disturbio (e.g., incendios forestales) (Brewer, 2015). Incluso, como en este caso, la reducción en la incorporación de individuos de Picea martinezii causa problemas genéticos, lo que en el largo plazo podría representar un reto más para el establecimiento de las plántulas de esta especie e implicar cambios estructurales, principalmente en la estructura florística, como es el caso de la comunidad El Bosque, donde se considera que ya se están presentando debido a que los grados de mezcla tienden a ser menores a los que presentan las demás comunidades.
Conclusiones
La estructura horizontal y dimensional que presentan las comunidades estudiadas es similar, a excepción de la localidad El Bosque. En estas comunidades, las especies arbóreas se distribuyen espacialmente de manera aleatoria, con altos grados de mezcla que dan origen a un estado de competencia entre individuos de diferentes especies, lo que es favorable para su establecimiento y crecimiento.
Las condiciones estructurales estables y favorables que presentan estas comunidades remarcan la importancia ambiental de seguir conservándolas, donde a pesar de que no se pueden realizar aprovechamientos forestales, estas representan un gran potencial para la provisión de servicios ambientales y la conservación de biodiversidad.
Con el análisis de la estructura de las comunidades estudiadas, se considera factible el recuperar áreas con características ecológicas similares cercanas a las estudiadas mediante trabajos de restauración utilizando germoplasma obtenido en estos sitios, tratando de replicar las condiciones estructurales de las especies arbóreas, conservando la diversidad florística y promoviendo de esta manera el reclutamiento de especies arbustivas y arbóreas.
Conflicto de interés
Los autores declaran no tener conflicto de intereses.

