











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El género Cucurbita incluye 27 subespecies y se ubica entre los primeros diez principales cultivos de hortalizas en el mundo (Balkaya et al., 2010). La especie pepo es la más cultivada en el mundo y con origen en América (Formisano et al., 2012). Como subespecie, y comercialmente, son altamente reconocidos los tipos cultivados Zucchini, Pumpkins y squashes (Montero‐Pau et al., 2018). En la familia cucurbitácea se sabe que la reproducción sexual es controlada por diferentes factores genéticos, ambientales y fitohormonales (Manzano et al., 2011). En el caso específico de Cucurbita pepo, es un cultivo monoico con dos o tres fases de desarrollo sexual; en la primera, solo se producen flores masculinas, y en la segunda se alterna el desarrollo de flores masculinas y femeninas, mientras que la fase final consiste solo en flores femeninas (Manzano et al., 2013). Dado que los órganos sexuales femeninos y masculinos están separados en flores unisexuales, el desarrollo de los frutos presenta una gran dependencia de polinizadores, proceso que se limita a las seis horas en que las flores de ambos sexos permanecen abiertas durante la mañana, por lo cual depende en gran medida de las condiciones ambientales que afectan la cantidad de flores femeninas y masculinas por planta, al igual que la actividad de los polinizadores (Loy, 2012; Martínez et al., 2014).
La polinización en Cucurbita pepo depende en gran medida de la actividad de insectos polinizadores, especialmente de las abejas del género Melipona (Ali et al., 2014); sin embargo, las poblaciones de estos polinizadores se han reducido drásticamente debido a la pérdida de hábitats, envenenamiento por plaguicidas, virus y ácaros parásitos; por ejemplo, el número de colmenas en los Estados Unidos se ha reducido de casi 6 millones en la década de 1940 a 2.3 millones en 2008 (Levy, 2011). Como resultado, se ha reportado una tendencia de menor tasa de crecimiento en el rendimiento de cultivos que dependen de polinizadores en comparación con aquellos con baja o ninguna dependencia (Garibaldi et al., 2009).
Adicionalmente, el aborto floral es un fenómeno biológico en la reproducción y evolución de las plantas, fenómeno observado en Cucurbita texana (Krupnick et al., 2000). El aborto floral en plantas como C. texana se considera que es resultado del estrés abiótico (Zhang et al., 2013). La alteración en las condiciones climáticas impondrá nuevas selecciones en el fenotipo y características que confieren tolerancia al estrés (Anderson et al., 2012). El estrés abiótico durante la reproducción tiene efectos adversos en el rendimiento de los cultivos; el aborto de semillas y frutos potencialmente viables ha llamado la atención de los ecologistas de plantas en las últimas dos décadas (Samarah et al., 2009). La polinización o la fertilización ineficaz, producida por temperaturas bajas o altas o humedad inadecuada, causa una disminución en la formación de los frutos, resultando en una disminución de la producción (Nepi et al., 2010). Algunas plantas presentan la capacidad de producir frutos sin la necesidad de fertilización por el gameto masculino, es decir, sin polinización, esta característica es denominada como partenocarpia (Joldersma & Liu, 2018).
La partenocarpia es un rasgo importante para mejorar el rendimiento, la calidad y el procesamiento de los frutos hortícolas, particularmente de las especies que son cultivadas en invernadero y son de polinización cruzada (Dhatt & Kaur, 2016). Incluso la partenocarpia tiene el potencial de generar frutos sin semillas, lo cual es una característica altamente deseable (Knapp et al., 2017). La partenocarpia ocurre de manera natural en algunas especies y se ha aprovechado intensivamente, tal es el caso de Musa spp. estéril triploide y las especies auto incompatibles del género Citrus spp., aunque los mecanismos intrínsecos de partenocarpia siguen sin ser comprendidos (Dash & Rai, 2016; Mesejo et al., 2016).
En el caso de Cucurbita pepo, la partenocarpia es un fenómeno reproductivo presente, y en el caso específico del tipo zucchini, su uso es limitado por la obtención de bajo rendimiento (Pomares-Viciana et al., 2017). Por ejemplo, en los resultados de Ribeiro et al. (2020) las plantas de polinización natural produjeron entre 300 g y 340 g de biomasa de frutos, mientras que los que presentaba partenocarpia natural no generaron frutos. Por lo tanto, para el aprovechamiento y estudio de la partenocarpia en Cucurbita pepo tipo zucchini es necesaria la identificación de genotipos con dicha característica reproductiva, y que puedan ser utilizados para la formación de nuevas variedades. La importancia económica y agroalimentaria para esta hortaliza es alta porque presenta consumos per cápita de hasta 11.5 g dia-1 (Aryal & Reinhold, 2011). El objetivo del presente estudio fue identificar genotipos partenocárpicos de Cucurbita pepo tipo Grey Zucchini.
Materiales y Métodos
Se cruzaron seis líneas parentales de calabaza Grey Zucchini y se obtuvieron las generaciones S1 y S2 correspondientes. Los cruzamientos y el cultivo de las líneas siempre se realizaron en condiciones controladas para evitar la recombinación genética. Del material genético generado y perteneciente a la empresa Evergreen HLT®, se seleccionaron 50 genotipos para su evaluación en el presente estudio, cada uno con cinco repeticiones, teniendo una población total de 250 plantas. El experimento se estableció en un invernadero ubicado en la Estación Joaquín, con coordenadas 20° 27' 12.7" N y 101° 28' 44.5" O, en el municipio de Abasolo, Guanajuato. Las semillas se sembraron en bolsas de 5.68 L calibre 600 mm, utilizando como sustrato suelo agrícola previamente homogenizado de manera mecánica mediante una mezcladora para cemento. Se utilizó fertirriego con una dosis de 200-150-150 más micronutrientes distribuida en dos aplicaciones semanales, con un cambio en las proporciones de los macronutrientes según la etapa vegetativa y/o reproductiva de las plantas. Durante el cultivo, las temperaturas máxima y mínima promedio fueron de 32.9 °C y 12.1 °C, respectivamente.
Respecto al manejo reproductivo de los genotipos, las flores masculinas se retiraron para evitar que las flores femeninas fueran polinizadas. Antes de la apertura de las flores femeninas, estas se cubrieron con bolsas de papel para evitar la introducción de insectos polinizadores y polen externo. Las bolsas se retiraron hasta el comienzo de la formación de frutos y se identificaron los genotipos partenocárpicos de acuerdo con su capacidad para generar frutos sin polinización. Como variables cuantitativas en los genotipos seleccionados, se determinó el peso de los frutos (PF, g) y con un vernier electrónico su longitud (LF, cm), midiendo desde la unión con el pedúnculo floral hasta el extremo apical. Se cuantificó el número de cortes (NC) como el promedio de frutos cortados por planta durante cuatro semanas después de la maduración del primer fruto. Después del corte de cada fruto, estos se lavaron con una solución de detergente catiónico comercial y sales de amonio cuaternarias 1% y se mantuvieron a 22 °C; posteriormente, como vida de anaquel (VA), se contabilizó el número de días en que los frutos perdieron la dureza comercialmente aceptable. Adicionalmente, los frutos se clasificaron por su morfología acostillada o cilíndrica (MF) realizando comparación visual con frutos comerciales. Durante la cosecha manual, se clasificó el desprendimiento de pedúnculo (DP) como viable y no viable. Los genotipos no viables presentan daños físicos y desprendimientos en la unión de pedúnculo con el fruto, esta comparación visual permitió la clasificación de los genotipos. Se determinó la susceptibilidad a la cenicilla (SC) (Erysiphe cichoracearum) y al mildiu (SM) (Pseudonospora cubensis), según se desarrollaron dichas enfermedades en el follaje. Las plantas se clasificaron por la presencia y/o ausencia de las enfermedades in situ y por la identificación de los cuerpos fructíferos por microscopía. Para las variables cuantitativas se realizaron análisis de varianza en un diseño completamente al azar con cinco repeticiones, así como pruebas de separación de medias de Tukey (p > 0.05). Las variables cualitativas se analizaron en un diseño completamente al azar con cinco repeticiones mediante la prueba de Kruskal-Wallis. Igualmente, se generó un dendrograma de similitud mediante el método Euclidiano, incluyendo todas las variables y los genotipos partenocárpicos. Todos los análisis estadísticos se realizaron con el software Minitab® 16.2.3.
Resultados y Discusión
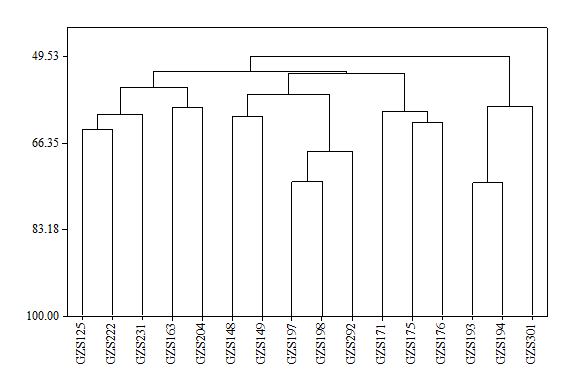
A partir de la población de 250 plantas con 50 genotipos, el 32% corresponde a los genotipos que fueron capaces de producir frutos sin polinización y por lo cual son considerados partenocárpicos: GZS125, GZS148, GZS149, GZS163, GZS171, GZS175, GZS176, GZS193, GZS194, GZS197, GZS198, GZS204, GZS222, GZS231, GZS292 y GZS301. La partenocarpia es un carácter heredable con modelos de control genéticos que varían desde un solo gen hasta múltiples loci de caracteres cuantitativos; en el caso de Curcubita pepo, el carácter es controlado por la expresión de genes relacionados con el metabolismo de los carbohidratos y la coordinación de fitohormonas durante la formación de los frutos (Pomares-Viciana et al., 2019). Se identificaron diferencias significativas (p < 0.01) en el PF entre los genotipos partenocárpicos, siendo el genotipo GZS301 el que presentó el mayor valor (160.3 g); sin embargo, no tendría un alto potencial productivo porque no presentó un alto número de cortes (Tabla 1). En Cucurbita spp. el número de frutos por planta es un carácter que se selecciona indirectamente por su correlación con el rendimiento (Aruah et al., 2012). Se identificaron diferencias altamente significativas (p < 0.01) en el NC, siendo los genotipos GZS171, GZS175, GZS197, GZS198 y GZS204 los que presentaron el mayor valor y, al mismo tiempo, bajos resultados de PF, en particular los cuatro primeros (121.7 g, 125.2 g, 113.6 g y 103.9 g, respectivamente). Un alto número de frutos por planta indica un mayor número de flores femeninas (Akter et al., 2013). Los resultados indican que los genotipos GZS171, GZS175, GZS197 y GZS198 presentaron el mayor número de flores femeninas partenocárpicas; aunque esto significaría que dichos genotipos tendrían un mayor número de frutos, estos no fueron los genotipos que presentaron los rendimientos más altos. En cambio, el genotipo GZS194, además de ser partenocárpico, presentó la mejor relación PF NC-1, por lo que potencialmente se podrían hacer 3.2 cortes por semana con frutos con pesos de 148.5 g. La partenocarpia en la familia Cucurbitaceae siempre ha formado parte de los componentes del rendimiento debido a que ha sido seleccionada indirectamente, ya que en los programas de mejoramiento convencionales no se han diferenciado si los frutos son obtenidos por partenocarpia, como en el caso de Cucumis sativus (Lietzow et al., 2016). Se identificaron diferencias altamente significativas (p < 0.01) en la LF de los genotipos partenocárpicos. De acuerdo con la norma NMX-FF-020-1982 (Secretaría de Comercio y Fomento Industrial, 1982), la categoría de más alta calidad Mexico Extra para calabacitas italianas considera un rango de longitud entre 8.5 cm y 13.0 cm, por lo que los frutos de los genotipos GZS125, GZS175, GZS197 y GZS231 se encontrarían en esta categoría. Existen diferencias altamente significativas (p < 0.01) en la VA, el genotipo GZS193 mantuvo la dureza de los frutos por ocho días, siendo el mayor valor de VA. Los genotipos GZS148, GZS171, GZS197 y GZS22 mantuvieron la dureza de los frutos por siete días (Tabla 1). La vida de anaquel es importante en la calabaza por ser altamente perecedera y particularmente sujeta a desecación y lesiones (Paris, 2016). La calidad de los frutos es afectada después de su cosecha por el manejo, factores ambientales y genéticos (Arah et al., 2015); respecto a estos últimos, los mecanismos genéticos de este carácter aún no son claros en Cucurbita spp., las evidencias en otras especies de importancia económica como el tomate indican un control dependiente de la interacción entre varios genes inducido por la regulación de factores de transcripción involucrados en diversos procesos del desarrollo de las plantas con dominios proteicos como los NAC, que corresponden a factores de transcripción específicos (Hu et al., 2010; Casals et al., 2012). Los genotipos partenocárpicos GZS193, GZS148, GZS171, GZS197 y GZS22 podrían constituir un excelente modelo de estudio para elucidar el control genético de la vida de anaquel en Cucurbita pepo, por sus altos valores presentados en la VA respecto a los demás genotipos. Se identificaron diferencias altamente significativas (p < 0.01) en la morfología de los frutos entre los genotipos partenocárpicos, de los cuales GZS148, GZS149, GZS171, GZS193, GZS194, GZS197, GZS198 y GZS292 tienen morfología acostillada. De los genotipos partenocárpicos, el 50% presentó frutos con morfología acostillada, por lo que las evidencias no sugieren que esta sea un carácter dominante asociado a la partenocarpia. La morfología acostillada en Cucurbita pepo podría tener su origen en algún ancestro en Oaxaca, en el sur de México (Gong et al., 2012), lo cual sugiere que alguna de las líneas progenitoras pudo tener dicha característica. Cabe mencionar que en el Bajío la morfología más aceptada es la acostillada, por lo que esta es una variable importante para considerar para el mercado local. Se identificaron diferencias altamente significativas (p < 0.01) entre los genotipos en el desprendimiento del pedúnculo (DP) durante la cosecha manual de los frutos. En este contexto, la partenocarpia es altamente controlada por diferentes fitohormonas, y la aplicación exógena de melatonina en Pyrus communis puede inducir dicha característica reproductiva y de manera indirecta modifica las características del pedúnculo, favoreciendo la cosecha manual de los frutos (Liu et al., 2018). En este experimento, los resultados en los genotipos partenocárpicos GZS125, GZS171, GZS175, GZS176, GZS204 y GZS222 indican que la cosecha manual fue viable porque presentaron un pedúnculo con una menor área de unión con el fruto. En la susceptibilidad a cenilla (SC) y a mildiu (SM) se identificaron diferencias altamente significativas (p < 0.01), donde los genotipos GZS148 y GZS149 presentaron resistencia a ambos patógenos, puesto que no se presentó o identificó la presencia de estas enfermedades en las plantas (Tabla 2). Aunque en el presente estudio se identificaron genotipos partenocárpicos, ninguno presentó todas las características evaluadas: alto peso y longitud de fruto, alto número de días de vida de anaquel, morfología del fruto acostilladas, desprendimiento manual del pedúnculo y resistencia a la cenicilla y el mildiu, que son de importancia agronómica y económica. El genotipo GZS194 presentó el mayor potencial productivo por su relación entre el peso de los frutos y los cortes por semana; además, mostró resistencia a la cenicilla y sus frutos tuvieron morfología acostillada. El genotipo GZS301 registró el mayor peso del fruto y resistencia a la cenicilla. El genotipo GZS193 fue uno de los que presentó mayor valor de VA, resistencia a la cenicilla y sus frutos presentaron morfología acostillada. El genotipo GZS148 presentó una larga vida de anaquel, sus frutos presentaron morfología acostillada, además de resistencia al mildiu y a la cenicilla. GZS171 presentó una larga vida de anaquel, el mayor número de cortes por semana, morfología acostillada y su cosecha manual fue viable. De acuerdo con Würschum (2012), la selección de líneas parentales como los genotipos mencionados y la selección de progenie superiores permiten la acumulación de alelos favorables. De acuerdo con el dendrograma, se identifican cuatro grupos con similitudes: a) 54.43, b) 56.39, c) 59.33 y d) 58.35, respectivamente (Figura 1), entre los cuales el último contiene a los genotipos GZS193, GZS194 y GZS301, lo cual sugiere que a dicha base genética se podrían mantener y adicionar los genes de los genotipos GZS148 y GZS171. La pirimidación de genes mediante el cruzamiento de genotipos con características agronómicas y de importancia económica, como la partenocarpia en el caso del presente estudio, es considerada una estrategia de mejoramiento genético eficiente y duradera (Souza et al., 2014), teniendo en cuenta que los genotipos partenocárpicos ya han sido identificados.
Tabla 1 Variables cuantitativas evaluadas en los genotipos partenocárpicos.
| Genotipos | PF** | LF** | NC** | VA** |
| GZS125 | 69.2 k | 12.3 bc | 4.0 bc | 5.8 bc |
| GZS148 | 92.4 hi | 13.3 abc | 3.0 bc | 7.0 ab |
| GZS149 | 82.6 j | 13.4 abc | 2.6 c | 4.8 c |
| GZS163 | 142.8 c | 13.6 abc | 4.2 b | 5.0 c |
| GZS171 | 121.7 e | 13.2 abc | 7.0 a | 7.0 ab |
| GZS175 | 125.2 e | 12.4 bc | 7.0 a | 6.0 bc |
| GZS176 | 74.6 k | 13.3 abc | 6.8 a | 5.0 c |
| GZS193 | 134.8 d | 14.7 ab | 3.0 bc | 8.0 a |
| GZS194 | 148.5 b | 15.4 a | 3.2 bc | 6.8 ab |
| GZS197 | 113.6 f | 13.0 abc | 7.0 a | 7.0 ab |
| GZS198 | 103.9 g | 14.0 abc | 7.0 a | 6.0 bc |
| GZS204 | 142.3 c | 14.0 abc | 6.6 a | 5.0 c |
| GZS222 | 95.5 h | 13.6 abc | 3.0 bc | 7.0 ab |
| GZS231 | 87.1 i | 12.0 c | 3.0 bc | 5.0 c |
| GZS292 | 73.9 k | 14.2 ab | 4.0 bc | 6.0 bc |
| GZS301 | 160.3 a | 14.4 ab | 2.8 bc | 6.0 bc |
| DMSH | 2.06 | 1.10 | 0.68 | 0.66 |
Peso de los frutos (PF, g); longitud de los frutos (LF, cm); número de cortes (NC); vida de anaquel (VA, días). Análisis de varianza: diferencias significativas p < 0.05 (*); diferencias altamente significativas p < 0.01 (**). Prueba de Tukey (p < 0.05): valores con la misma letra dentro de columnas de promedios son estadísticamente iguales, diferencia mínima significativa honesta (DMSH).
Fuente: Elaboración propia.
Tabla 2 Variables cualitativas evaluadas en los genotipos partenocárpicos.
| Genotipos | MF** | DP** | SC*** | SM** |
| GZS125 | C | V | S | R |
| GZS148 | A | NV | R | R |
| GZS149 | A | NV | R | R |
| GZS163 | C | NV | S | R |
| GZS171 | A | V | S | S |
| GZS175 | C | V | S | S |
| GZS176 | C | V | S | S |
| GZS193 | A | NV | R | S |
| GZS194 | A | NV | R | S |
| GZS197 | A | NV | R | S |
| GZS198 | A | NV | R | S |
| GZS204 | C | V | S | R |
| GZS222 | C | V | S | R |
| GZS231 | C | NV | S | R |
| GZS292 | A | NV | R | S |
| GZS301 | C | NV | R | S |
Morfología de los frutos (MF, acostillada [A] o cilíndrica [C]); desprendimiento del pedúnculo (DP, viable [V] o no viable [NV]); susceptibilidad a la cenicilla (SC, susceptible [S] o resistente [R]); susceptibilidad al mildiu (SM, susceptible [S] o resistente [R]). Kruskal-Wallis: diferencias significativas p < 0.05 (*), diferencias altamente significativas p < 0.01 (**).
Fuente: Elaboración propia.
Conclusiones
De los 50 genotipos evaluados, se identificaron 16 partenocárpicos. Los genotipos partenocárpicos GZS193, GZS194, GZS301, GZS148 y GZS171 presentaron la mayoría de las características, que le confieren a la especie su valor agronómico y económico. El mayor peso y longitud de fruto fueron 148.5 g y 14.7 cm; ocho días fue el mayor valor de vida de anaquel, se presentaron frutos con morfología acostillada y viables para su cosecha manual, así como genotipos resistentes a la cenicilla y el mildiu. Los genotipos identificados se podrían usar como progenitores para mantener la partenocarpia y acumular todas las características en un solo genotipo mediante un siguiente ensayo de cruzamientos dirigidos.

