











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El calentamiento global y el cambio climático representan un problema mundial que afecta la seguridad alimentaria y el bienestar de algunos sectores más vulnerables de la población humana. Esta problemática es influenciada por el incremento de gases de efecto invernadero (GEI), como el dióxido de carbono, metano y óxido nitroso, procedentes de actividades antropogénicas (United Nations [UN], 2012). En este sentido, el metano (CH4) es uno de los principales GEI (Intergovernmental Panel on Climate Change [IPCC], 2006), el cual es un subproducto de la fermentación anaeróbica de los carbohidratos, y es producido por los rumiantes, donde las arqueas metanogénicas y los protozoarios ruminales tienen una función importante en la síntesis de este gas. Estos microrganismos utilizan el bióxido de carbono (CO2) y el hidrógeno (H2) que se origina a partir de la síntesis de los ácidos grasos volátiles de cadena corta (AGV), que son producto de la fermentación microbiana de los carbohidratos contenidos en los follajes y granos (Martin, Morgavi & Doreau, 2010; Poppi & McLennan, 1995). Principalmente, las archaeas metanogénicas utilizan el H2 metabólico y el CO2 para formar CH4 y así reducir la acumulación de H2 en el rumen (Benaouda, Gonzáles, Molina & Castelán, 2017). En la actualidad, la ganadería emite el 18% de este gas y la fermentación ruminal contribuye con el 39.1% de las emisiones producidas por las actividades ganaderas (Gerber et al., 2013). Diversos autores mencionan que los rumiantes alimentados con pasturas tropicales que contienen altas concentraciones de fibra (> 70%) y baja calidad (bajo contenido de proteína cruda [PC] y digestibilidad), además de ser escasas en la mayor parte del año, conllevan a una mayor producción de CH4 (Kurihara, Magner, Hunter & McCrabb, 1999; Melesse, Steingass, Schollenberger, Holstein & Rodehutscord, 2017), debido al bajo aporte de proteína, minerales y energía al rumen. La elevada producción de CH4 representa una pérdida de la energía bruta consumida, la cual puede llegar hasta el 12% (Johnson & Johnson, 1995). Por lo tanto, contribuir a mitigar los efectos de CH4 se ha reconocido como un objetivo importante para combatir el cambio climático y, por otro lado, para incrementar los beneficios económicos a los ganaderos al mejorar la eficiencia en la utilización de la energía. En este sentido, el uso de alimentos balanceados mejora la eficiencia productiva y reduce las emisiones de GEI (Benaouda et al., 2017); sin embargo, los alimentos balanceados empleados en la nutrición de rumiantes como estrategia de suplementación durante la época de secas y formulados a base de granos afectan la seguridad alimentaria, lo que provoca que sean comercializados a precios elevados y, por lo tanto, que la rentabilidad de los sistemas de producción sea menor (Castañeda-Serrano, Piñeros-Varón & Vélez-Giraldo, 2017).
Durante la época de secas y lluvias, la vegetación tropical secundaria representa una abundante fuente de biomasa y proteína para los rumiantes (Ventura-Cordero, González-Pech, Torres-Acosta, Sandoval-Castro & Tun-Garrido, 2017), con potencial forrajero y capacidad para mitigar las emisiones de CH4 entérico, por su contenido de taninos condensados y saponinas (Piñeiro-Vázquez et al., 2018). Esta vegetación tropical secundaria (follajes, frutos, flores y vejucos) representa una fuente de PC para la dieta de rumiantes debido a que su contenido proteico y mineral es superior a la de los pastos tropicales. Entre estas especies, las leguminosas han demostrado potencial como estrategia nutricional en el trópico, principalmente en periodos de escases de alimento, puesto que tienen un alto contenido de proteína, el cual varía de 140 g/kg a 240 g/kg de materia seca (MS) (Castañeda-Serrano et al., 2017). Estudios como los de Woodward, Waghorn, Lassey & Laboyrie (2002), Hess et al. (2003) y Animut et al. (2008) han demostrado que las arbóreas que contienen taninos condensados (TC) reducen las emisiones de CH4. En este sentido, es importante evaluar alternativas forrajeras que tengan la capacidad de reducir las emisiones de GEI sin afectar la digestibilidad y que mejoren el comportamiento productivo de los rumiantes en las épocas de escaso alimento. Por lo tanto, el objetivo de este estudio es evaluar el potencial nutritivo, el efecto sobre la fermentación ruminal y producción de metano in vitro de follajes arbóreas y arbustivas con potencial forrajero en la región tropical de México.
Materiales y métodos
Localización
El trabajo se realizó en el laboratorio del Instituto Tecnológico de Conkal, Yucatán, ubicada al este del municipio de Conkal (21°, 05’, N; 89°, 32’, O, con una altitud de 9 msnm).
Muestreo de arbóreas forrajeras
Se recolectaron hojas frescas de Gliricidia sepium, Guazuma ulmifolia, Leucaena leucocephala, Morus alba, Tithonia diversifolia, Brosimum alicastrum, Cynodon nlemfluensis, Megathyrsus maximus, Piscidia piscipula y Hibiscus rosa-sinensis. Posteriormente, el material fue secado en una estufa de aire forzado a 55 °C hasta obtener un peso constante y fueron molidas a un tamaño de partícula de 1 mm en un molino (Azteca, Monterrey, Nuevo León, México), previo al análisis químico.
La recolección de follaje se realizó en los meses de junio y agosto de 2018, en la región central de Conkal, Yucatán, México. Esta región tiene un clima cálido subhúmedo con precipitaciones regulares en el verano (mayo-agosto), una temperatura promedio anual de 26 °C, precipitaciones de 984.4 mm y fluctuaciones de humedad relativa del 66% al 89% (García, 1981). Los suelos son calcáreos, poco profundos y con altos porcentajes de rocas llamados litosol (tzek'el en lengua maya) (Duch, 1988). La vegetación es de baja densidad y los arbustos alcanzan entre 4 m y 7 m de altura. El área selvática de la región se caracteriza por una selva baja caducifolia (Gutiérrez-Báez & Zamora-Crescencio, 2012).
Análisis químico
Las muestras recolectadas del follaje fueron molidas a través de un tamiz de 1 mm para su análisis, según la Association of Official Analytical Chemists(AOAC) (1990). El contenido de materia seca (MS) del forraje se determinó en una estufa de aire forzado a 57 °C durante 48 h (peso constante). Las determinaciones de nitrógeno (PC = N × 6.25) se llevaron a cabo con un LECO® CN2000 series 3740 (LECO® Corp., St. Joseph, MI) (AOAC, método número 992.15). El contenido de materia orgánica (MO) de las muestras se determinó por combustión en una mufla a 550 °C (AOAC, método número 923.03), y la concentración de fibra detergente neutro (FDN) y fibra detergente acida (FDA) se determinó según lo sugerido por Van Soest, Robertson & Lewis (1991). La concentración de TC se determinó según lo sugerido por Terril, Rowan, Douglas & Barry (1992). El contenido de saponinas (Sa) se determinó según lo sugiere Makkar (2006).
Animales y muestreo de líquido de rumen
Se utilizaron cuatro ovinos de pelo con un peso vivo promedio (PV) de 31 kg ± 1 kg, con una edad de siete meses como donadores de líquido ruminal. Los animales fueron alimentados dos veces al día por un periodo de dos meses con una dieta basada en 60% de forraje y 40% de concentrado y un contenido de proteína cruda de 16% kg/MS; adicionalmente, se proporcionó sal mineral y agua a libre acceso. El líquido ruminal fue tomado antes de la alimentación, el cual se extrajo mediante sonda esofágica (Ramos-Morales et al., 2014), posteriormente se colocó en un termo hermético a 39 °C en condiciones anaeróbicas y se transportó hasta el laboratorio para luego ser filtrado con gasas dobles.
Producción de gas in vitro
La producción de gas se determinó mediante el procedimiento sugerido por Menke & Stengass (1988) y modificado por Makkar, Blümmel & Becker (1995). Se colocó 1 g de materia seca (MS) de cada forraje molido a un tamaño de partícula de 1 mm en frascos de vidrio color ámbar de 120 mL. Después, se adicionaron 90 mL de inóculo ruminal mezclado en una relación 1:9, con una solución mineral reducida según el método propuesto por Menke & Stengass (1988). Conforme se agregó el inóculo, el flujo continuo de CO2 fue constante con la finalidad de mantener las condiciones de anaerobiosis. El flujo de CO2 se retiró y se colocó un tapón de goma y un arillo de aluminio para sellar los frascos a presión. El aire del interior de cada frasco se extrajo hasta igualar la presión interior a cero e inmediatamente después fueron colocados en un baño maría a una temperatura constante de 39 °C. Los datos fueron corregidos con base a los blancos incubados bajo las mismas condiciones, con la diferencia de que no contenían sustrato de las arbóreas o arbustivas.
La presión de gas generada por la fermentación del sustrato se midió con un manómetro con escala de 0 kg/cm-2 a 1 kg/cm-2 con aguja hipodérmica. En cada medición se extrajo el gas hasta bajar la presión interior nuevamente a 0. Las lecturas se realizaron a las 2 h, 4 h, 6 h, 8 h, 12 h, 16 h, 20 h, 24 h, 30 h, 36 h, 42 h, 48 h, 60 h y 72 h de incubación y se transformó en volumen de gas utilizando la siguiente ecuación de regresión logística: V = (P + 0.019) (0.024)−1 (Elmasry et al., 2016), donde V = volumen de gas producido y P = presión que se genera en cada botella.
Cinética de fermentación de gases in vitro y fracciones fermentables.
El volumen máximo (Vm, mL g−1), la tasa de fermentación (S, h−1) y la fase lag (L, h) de la producción de gas se estimaron en el PROC GLM del statistical analysis software (SAS, 2008) utilizando el siguiente modelo logístico: V = Vm/1+ e(2 - 4 S(t - L) (Pell & Schofield, 1993), donde V = volumen de gas en el momento (t), Vm = volumen máximo de gas, S = velocidad de fermentación específica (similar a la tasa de degradación) y L = tiempo de colonización de microorganismos. También se obtuvieron volúmenes acumulados de gas para los intervalos de tiempo de 0 h a 8 h (Vf0-8), de 8 h a 24 h (Vf8-24) y de 24 h a 72 h (Vf24-72) de incubación. Estos volúmenes se utilizaron para estimar las fracciones (mg/g-1MS incubada) de fermentación rápida (FR), fermentación media (FM) y fermentación lenta (FL), según las ecuaciones de regresión lineal propuestas por Miranda, Vázquez, Améndola, Sandoval & González (2015): FR = Vf0-8/0.427; FM = Vf8-24/0.615; y FL = Vf24-72/0.345. La suma de las tres fracciones es la fracción fermentable total (FT).
Degradabilidad in vitro de materia seca y materia orgánica.
Después del periodo de incubación de los sustratos experimentales (72 h), los residuos de cada sustrato se recolectaron filtrándolos a través de bolsitas de dacrón (10 cm x 4 cm). La degradabilidad in vitro de la MS (DEGMS) se estimó secando el material residual a 55 °C hasta lograr peso constante. La degradabilidad in vitro de la MO (DEGMO) se estimó por combustión en una mufla a 550 °C (AOAC 1980, método número 923.03). Posteriormente, el DEGMS (%) se calculó basándose en la MS inicial y residual (Monforte-Briceño, Sandoval-Castro, Ramírez-Avilés & Capetillo, 2005) y la DEGMO (%) se calculó de manera similar basándose en la MO inicial y residual (Miranda et al., 2015).
Producción de metano
La producción total de gas durante 24 h de fermentación se midió con una jeringa de vidrio (Becton Dickinson BD Luer) a las 6 h, 12 h, 18 h y 24 h de incubación. Todas las muestras fueron depositadas en botellas de vidrio de 25 mL, cerradas herméticamente con tapones de goma y anillos de aluminio. En cada botella se depositaron 25 mL de una solución salina saturada (1 L de agua por 1 kg de cloruro de sodio [NaCl]) en una proporción de 1:1 para formar una trampa y mantener las propiedades del gas. Todas las muestras fueron analizadas para determinar la concentración de CH4 a través de un cromatografo de gases marca Aligent Technology 7890B, modelo G3440B, equipado con un detector de ionización de llama (FID) y una columna (HP-5) de 30 m × 0.32 mm × 0.25 µm. Se inyectaron 5 mL de gas con una jeringa de 60 mL a una temperatura de 60 °C y una velocidad de acarreo de 80 mL min-1 usando helio como gas transportador (Bhatta, Saravanan, Baruah & Prasad, 2015; Kajikawa, Tajima, Mitsumori & Takenaka, 2007).
Diseño experimental y análisis estadístico
Se analizaron las variables de fermentación in vitro y digestibilidad (Vm, S, L, RF, MF, LF, DEGMS, DEGMO) y producción de CH4, utilizando un diseño experimental en bloque con repeticiones en el tiempo, en el que cada tratamiento tuvo ocho repeticiones. Los datos fueron analizados en base a (Cochran & Cox, 1990) un modelo lineal general mediante con el procedimiento PROC ANOVA del SAS® 9.2 software (SAS Inst. Inc., Cary, NC, 2008). La comparación de medias se realizó mediante la prueba de Tukey (p < 0.05). Se utilizó el siguiente modelo matemático: Yij = µ + ηi + αj + εij (Montgomery, 2013).
Resultados
Composición química
La composición química del follaje de las arbóreas y arbustivas se presenta en la Tabla 1. El contenido de PC más alta se registró en G. sepium (21.78%), seguida de T. diversifolia (21.76%) y L. leucocephala (21.37%), y el contenido de PC más bajo fue en el pasto M. maximus (6.14%). Los contenidos de FDN y FDA variaron de 44.34% a 77.43% y de 16.12% a 49.53% de la MS en las diferentes especies. La concentración de TC de L. leucocephala fue mayor (1.16% de la MS), mientras que en G. sepium la concentración fue menor (0.08% de MS). Las especies P. piscipula y G. ulmifolia obtuvieron mayor contenido de Sa (0.90% y 0.91% de la MS), y las especies T. diversifolia, H. rosa-sinensis, M. maximus y B. alicastrum obtuvieron menor concentración en el rango de 0.09% a 0.07% de MS (Tabla 2).
Tabla 1 Composición química del follaje de las arbóreas y arbustivas (% de la MS).
| Especies | MS | MO | PC | EE | FDN | FDA |
| Guazuma ulmifolia | 45.51 | 5.60 | 13.23 | 3.30 | 61.78 | 45.29 |
| Brosimum alicastrum | 42.05 | 4.57 | 14.14 | 2.50 | 52.02 | 38.37 |
| Leucaena leucocephala | 37.41 | 3.33 | 21.37 | 5.07 | 59.52 | 32.42 |
| Piscidia piscipula | 35.19 | 3.86 | 12.91 | 2.76 | 58.90 | 43.33 |
| Cynodon nlemfuensis | 30.89 | 2.16 | 14.66 | 2.81 | 71.88 | 36.88 |
| Gliricidia sepium | 26.44 | 3.32 | 21.78 | 4.74 | 53.33 | 27.21 |
| Megathyrsus maximus | 24.21 | 2.29 | 6.14 | 7.85 | 77.43 | 49.53 |
| Morus alba | 22.48 | 3.01 | 16.94 | 4.54 | 44.34 | 27.90 |
| Hibiscus rosa-sinensis | 21.97 | 2.67 | 14.02 | 4.51 | 51.94 | 16.12 |
| Tithonia diversifolia | 16.48 | 3.06 | 21.76 | 4.84 | 50.65 | 36.80 |
MS: Materia seca; MO: Materia orgánica; PC: Proteína cruda; EE: Extracto eterio; FDN: Fibra detergente neutra; FDA: Fibra detergente ácida. F: forraje; M: madera; CV: Cercas vivas; G: gastronomía; L: leña; SSP: Sistemas silvopastoriles; BB: banco de biomasa; MD: medicinal; O: ornamental.
Fuente: Elaboración propia.
Tabla 2 Contenido de taninos condensados y saponinas en el follaje de arbóreas y arbustivas (% de la MS).
| Especies | TC (% de la MS) | SAP (% de la MS) | Tipo de crecimiento |
| Guazuma ulmifolia | 0.47 | 0.91 | Arbustiva |
| Brosimum alicastrum | 0.22 | 0.07 | Arbustiva |
| Leucaena leucocephala | 1.16 | 0.43 | Arbustiva |
| Piscidia piscipula | 0.19 | 0.90 | Arbustiva |
| Cynodon nlemfuensis | 0.11 | 0.13 | Pasto |
| Gliricidia sepium | 0.08 | 0.12 | Arbustiva |
| Megathyrsus maximus | 0.15 | 0.09 | Pasto |
| Morus alba | 0.13 | 0.15 | Arbustiva |
| Hibiscus rosa-sinensis | 0.13 | 0.09 | Arbustiva |
| Tithonia diversifolia | 0.12 | 0.09 | Arbustiva |
TC: taninos condensados; Sap: saponinas totales.
Fuente: Elaboración propia.
Cinética de fermentación de gas in vitro y volúmenes fraccionales
El volumen máximo (Vm) y la tasa de fermentación (S) fue similar entre las especies incubadas (p > 0.05). Los Vm más altos se encontraron en T. diversifolia y B. alicastrum (348.40 y 373.81 mL/g de MS incubado, respectivamente) (Figura 1), mientras que los Vm similares fueron encontrados en H. rosa-sinensis, C. nlemfuensis, M. alba, G. sepium, L. leucocephala y M. maximus (305.21, 319.72, 333.57, 338.99, 336.25 y 348.89 ml/g de MS incubada). Los Vm más bajos fueron observados en G. ulmifolia y P. piscipula (234.06 y 304.97 ml/g de MS incubada). La tasa más alta de fermentación se registró en H. rosa-sinensis (0.04 h-1). Mientras tanto, T. diversifolia, M. maximus, C. nlemfuensis, G. sepium, P. piscipula, L. leucocephala, G. ulmifolia y M. alba tuvieron una S similar, observando un promedio de 0.03 h-1 (p > 0.05). La tasa más baja la obtuvo B. alicastrum (0.02 h-1). Finalmente C. nlemfuensis y M. maximus requirieron más tiempo (L) para iniciar la fermentación (de 6.46 h a 7.50 h) con respecto a P. piscipula, L. leucocephala, M. alba, G. ulmifolia y G. sepium (de 0.55 h a 3.92 h). La T. diversifolia, H. rosa-sinensis y B. alicastrum obtuvieron valores similares en la fase lag (4.72 h, 5.20 h y 5.27 h) y menores que C. nlemfuensis y M. maximus (p > 0.05). No se observaron diferencias en el volumen fraccional fermentable de la MS de los diferentes sustratos incubados (p > 0.05). La fracción más alta de FR (V0-8) se encontró en P. piscipula (100.96 ml/g de MS incubado); por otro lado, la fracción fermentable fue similar entre G. sepium, M. alba y L. leucocephala (83.69, 89.86 y 91.09 ml/g de MS incubado), H. rosa-sinensis y T. diversifolia (72.17 y 78.75 ml/g de MS) y C. nlemfuensis y B. alicastrum (61.48 y 68.07 ml/g de MS incubado) (p > 0.05). La fracción más baja de FR se encontró en G. ulmifolia y M. maximus (56.14 a 56.55 mg/g de MS incubado). Con respecto a FM (V8-24), G. sepium y H. rosasinensis tuvieron la mayor fracción (104.66 y 107.80 ml/g de MS incubado, respectivamente), seguidos por P. piscipula, M. alba, L. leucocephala, T. diversifolia, B. alicastrum, C. nlemfuensis y M. maximus (79.28 ml/g, 83.84 ml/g, 88.97 ml/g, 94.11 ml/g, 96.67 ml/g, 98.96 ml/g y 99.53 ml/g de MS incubado, respectivamente).
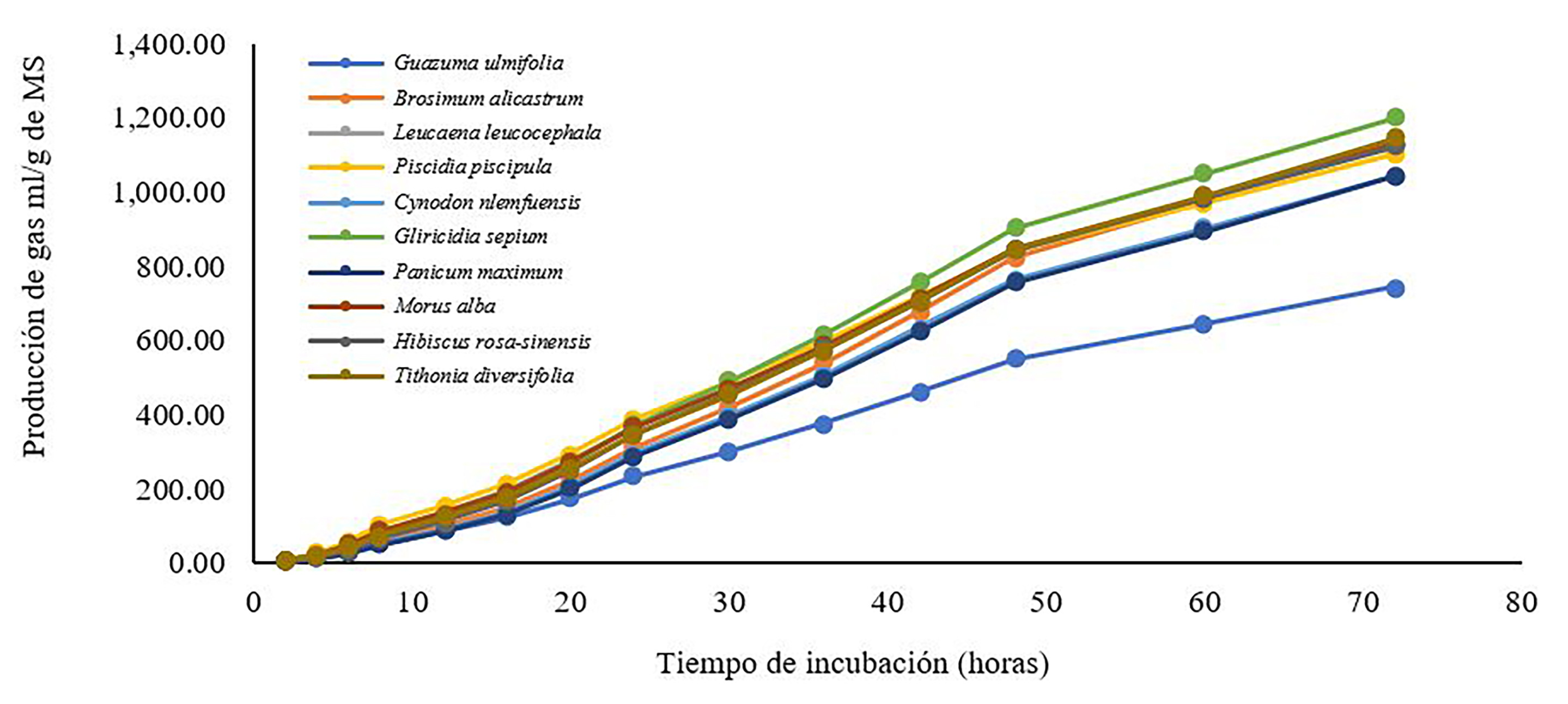
Fuente: Elaboración propia.
Figura 1 Producción de gas in vitro del follaje de arbóreas arbustivas durante 72 h de incubación.
La fracción más baja de FM se encontró en G. ulmifolia (56.75 ml/g de MS incubado). En la FL (V24- 72), T. diversifolia, M. maximus y B. alicastrum tuvieron la mayor fracción (172.54, 192.81 y 209.07 ml/g de MS incubado, respectivamente) y los G. sepium, M. alba, C. nlemfuensis y L. leucocephala fueron similares (150.64 ml/g, 154.71 ml/g, 159.28 ml/g, 161.31 ml/g de MS) (p > 0.05). La fracción más baja de FL se encontró en H. rosa-sinensis (125.24 ml/g de MS), P. piscipula (124.73 ml/g de MS) y G. ulmifolia (121.18 ml/g de MS) (Tabla 3).
Tabla 1 Parámetros de la producción de gas total y volúmenes fraccionales en el follaje de arbóreas arbustivas.
| Especies | Parámetros | Volúmenes fraccionales (ml g-1 MS) | |||||
| EM (MJ/kg) | Vm (ml g-1 MS) | S (ml h-1) | L (h) | V 0-8 | V 8-24 | V 24-72 | |
| G. ulmifolia | 9.07 | 234.06 | 0.03 | 2.97c | 56.14 | 56.75 | 121.18 |
| B. alicastrum | 16.47 | 373.81 | 0.02 | 5.27b | 68.06 | 96.67 | 209.07 |
| L. leucocephala | 14.09 | 336.25 | 0.03 | 1.84d | 91.09 | 83.84 | 161.31 |
| P. piscipula | 12.81 | 304.97 | 0.03 | 0.55e | 100.96 | 79.28 | 124.73 |
| C. nlemfuensis | 14.07 | 319.72 | 0.03 | 6.46ª | 61.48 | 98.96 | 159.28 |
| G. sepium | 15.91 | 338.99 | 0.03 | 3.92c | 83.69 | 104.66 | 150.64 |
| M. maximus | 14.94 | 348.89 | 0.03 | 7.50ª | 56.55 | 99.53 | 192.81 |
| M. alba | 14.65 | 333.54 | 0.03 | 1.85d | 89.86 | 88.97 | 154.71 |
| H. rosa-sinensis | 14.41 | 305.21 | 0.04 | 5.20b | 72.17 | 107.80 | 125.24 |
| T. diversifolia | 16.38 | 348.40 | 0.03 | 4.72b | 78.75 | 94.11 | 175.54 |
| Valor de p | 0.305 | 0.584 | 0.158 | 0.001 | 0.075 | 0.132 | 0.529 |
| EEM | 3.806 | 41.474 | 0.004 | 1.000 | 11.296 | 11.705 | 30.854 |
Vm: Volumen máximo de gas; S: tasa de producción de gas; L: Fase lag; V0-8: Volumen fraccional en la fracción rápidamente fermentable (0 h-8 h); V8-24: Volumen fraccional de gas en la fracción media (8 h-24 h); V24-72: Volumen fraccional de gas en la fracción de lenta fermentación (24 h-72 h); EEM: error estándar de la media. Letras distintas en la misma columna indican diferencias (p < 0.05).
Fuente: Elaboración propia.
Producción de CH4 ruminal y degradabilidad de la materia seca y orgánica
La producción de CH4 fue similar entre follajes incubados (p < 0.05). Sin embargo, se observó una menor emisión en L. leucocephala (14.4 ml/g MS incubada) con respecto a T. diversifolia, P. piscipula, M. alba, B. alicastrum, M. maximus y G. ulmifolia (20.17, 27.26, 32.11, 32.30, 34.00 y 34.06 ml/g MS incubado, respectivamente), mientras que las especies con mayor producción fueron C. nlemfuensis, G. sepium y H. rosa-sinensis (37.60 ml/g, 40.49 ml/g y 47.64 ml/g MS incubado) (Tabla 4). Se encontraron diferencias entre los follajes para DEGMS y DEGMO (p < 0.05). La DEGMS más alta (p < 0.05) se encontró en G. sepium, T. diversifolia, M. alba y H. rosa-sinensis (610.33, 760.40, 800.16 y 800.66 g/kg de MS). Se registró una DEGMS menor para B. alicastrum, C. nlemfuensis, G. ulmifolia, L. leucocephala y P. piscipula (550.90 g/kg, 490.33 g/kg, 470.00 g/kg, 450.56 g/kg, 400. 63 g/kg de MS), mientras que la más baja se registró en M. maximus (390.40 g/kg de MS). Los valores más altos de DEGMO (p < 0.05) se encontraron en T. diversifolia, M. alba y H. rosa-sinensis (698.90 g/kg, 762.30 g/kg y 766.70 g/kg de MO, respectivamente), seguidos de G. sepium, B. alicastrum, C. nlemfuensis y G. ulmifolia (542.00, 482.30, 408.00 y 388.90 g/kg de MO, respectivamente). La L. leucocephala, P. piscipula y M. maximus tuvieron una DEGMO menor (366.70 g/kg, 306.40 g/kg y 287.40 g/kg de MO, respectivamente) (Tabla 4).
Tabla 2 Producción de metano y degradabilidad in vitro del follaje de especies arbóreas.
| Especies | CH4 (ml/g MS) | DEGMS (g/kg de MS) | DEGMO (g/kg de MS) |
| Guazuma ulmifolia | 34.06 | 470.00c | 388.90c |
| Brosimum alicastrum | 32.30 | 550.90c | 482.30c |
| Leucaena leucocephala | 14.47 | 450.56c | 366.70d |
| Piscidia piscipula | 32.11 | 400.63d | 306.40d |
| Cynodon nlemfuensis | 37.70 | 490.33c | 408.00c |
| Gliricidia sepium | 40.49 | 610.33b | 542.00b |
| Megathyrsus maximus | 34.00 | 390.40d | 287.40d |
| Morus alba | 27.26 | 800.16ª | 762.30ª |
| Hibiscus rosa-sinensis | 47.64 | 800.66ª | 766.70ª |
| Tithonia diversifolia | 20.17 | 760.40b | 698.90b |
| EEM | 9.58 | 30.70 | 43.70 |
| Valor de P | 0.5374 | 0.0001 | 0.0001 |
CH4: Metano; DEGMS: Degradabilidad in vitro de la materia seca; DEGMO: degradabilidad in vitro de la materia orgánica; EEM: error estándar de la media. Letras distintas en la misma columna indican diferencias (p < 0.05).
Fuente: Elaboración propia.
Discusión
Composición química
La mayoría de las especies arbóreas tienen un alto contenido de PC y son bajas en fibra a diferencia de C. nlemfuensis y M. maximus, gramíneas que contienen una menor concentración de PC y mayor concentración de carbohidratos estructurales (Archimède et al., 2011; Kennedy & Charmley, 2012). Estos factores reducen la digestibilidad y la fermentación ruminal, afectando la tasa de pasaje y el tiempo de degradación en el rumen (Piñeiro-Vázquez et al., 2018). En este estudio, tres de las diez especies mostraron alto contenido de PC, G. sepium (210.7 g/kg de MS), T. diversifolia (217.6 g/kg de MS) y L. leucocephala (213.7 g/kg de MS), estos valores son similares a los reportados por Soltan et al. (2013) y Vite et al. (2019). Sin embargo, los valores de FDN y FDA son más altos que lo reportado por los mismos autores. Estas variaciones podrían explicarse por la edad de la planta, época del año y tipo de suelo donde se encontraban las plantas; cabe mencionar que los datos de FDN y FDA son similares a los reportados por Albores-Moreno et al. (2018).
Todas las especies presentaron cantidades moderadas de TC y Sa (Tabla 2), esto varía ampliamente dependiendo de las especies de plantas. El contenido de TC y Sa puede afectar la digestión y el uso de nutrientes en rumiantes debido a la reducción microrganismos en el rumen (Patra et al., 2017), como pudo haber sido el caso de P. piscipula, G. ulmifolia y L. leucocephala; ya que son las tres arbóreas que obtuvieron mayor concentración en TC (0.19%, 0.47% y 1.16% de la MS) y Sa (0.90%, 0.91% y 0.43% de MS). El contenido de TC de L. leucocephala es similar a lo reportado por Soltan et al. (2013). Melesse et al. (2017) realizaron pruebas cualitativas del tamizaje fitoquímico de estos árboles y arbustos donde observaron moderada presencia de TC y Sa, comparables a lo reportado en este estudio.
Cinética de fermentación de gas in vitro y volúmenes fraccionales
El efecto de los TC y Sa en la digestión de rumiantes se refleja en la cinética de fermentación y en la producción de gas in vitro. En este estudio, es posible que el contenido de TC en las especies de P. piscipula, G. ulmifolia y L. leucocephala (0.19%, 0.47% y 1.16% de MS, respectivamente) y el contenido de Sa (0.43%, 0.90% y 0.91% de MS, respectivamente) resultaran en una baja producción total de gas (Vm) con respecto a T. diversifolia, M. maximus y B. alicastrum. Esto se debe a que los TC son capaces de inhibir o modular la actividad enzimática y microbiana en la MO (Patra et al., 2017), lo que provoca una reducción en la fermentación (Bhatta et al., 2015; Puchala et al., 2012). Resultados similares fueron reportados por Tan et al. (2011), quienes evaluaron la inclusión de TC de L. leucocephala en una dieta para rumiantes (incubando 10.0 mg, 15.0 mg, 20.0 mg, 25.0 mg y 30.0 mg en 0.500 g), observando una reducción en la tasa de fermentación (0.0002 h-1 a 0.0176 h-1) y una menor producción total de gas (86.4 ml g-1 a 49.8 ml g-1 de MS incubada). Diversos autores indican que la mayor producción de gas acumulado encontrada en las especies M. maximus, G. sepium, T. diversifolia y B. alicastrum (Tabla 3) está relacionada con su menor contenido de TC (0.08%, 0.12% y 0.22% de MS) y Sa (0.12%, 0.09% y 0.07% de MS) (Albores-Moreno et al., 2018).
Producción de metano entérico y degradabilidad de la materia seca y orgánica
Las plantas con mayor potencial para mitigar CH4 fueron L. leucocephala, T. diversifolia, P. piscipula, M. alba, B. alicastrum, M. maximus y G. ulmifolia (20.17, 27.26, 32.11, 32.30, 34.00 y 34.06 ml/g MS incubado); ya que emitieron menos CH4 (Tabla 4). La reducción en la producción de CH4 en la L. leucocephala, P. piscipula, B. alicastrum y G. ulmifolia está asociada con el mayor contenido de TC y Sa (Tabla 2), debido a que tiene efecto directo sobre los metanógenos, protozoarios y tienen la capacidad de incrementar la proporción molar de ácido propiónico y reducir la producción de CH4 (Patra et al., 2017; Soltan et al., 2013; Tan et al., 2011). Además, Beauchemin, McGinn, Martinez & McAllister (2007) sostienen que altas concentraciones de compuestos fenólicos mitigan la producción de CH4 debido a la eliminación parcial de los protozoarios y de archaeas metanogénicas (Yuste, Amanzougarene, de la Fuente, de Vega & Fondevila, 2019). Resultados similares son reportados por Albores-Moreno et al. (2018), donde observaron que las hojas de P. piscipula y L. leucocephala redujeron la producción de CH4 entérico en 15.6% a 31.6%, y aumenta la proporción de ácido propiónico y reducción en la proporción de ácido acético. En este sentido, Soltan et al. (2013) observaron que la inclusión de follaje de L. leucocephala que contiene TC disminuye la producción de CH4 en 14.1%, esto debido al contenido de TC y su efecto sobre la población de protozoarios y bacterias encargadas de la síntesis de CH4. Resultados similares fueron reportados por Tan et al. (2011), quienes evaluaron niveles crecientes de inclusión de L. leucocephala (10.0, 15.0, 20.0, 25.0 y 30.0 mg TC/500 mg de MS), reportando una reducción del 33.0% en la producción de CH4 en la dosis superior (30.0 mg TC/500 mg de MS). La producción de CH4 observada en este estudio en T. diversifolia y M. alba podría estar asociada a la alta digestibilidad de la materia orgánica (Tabla 4). En este sentido, diversos autores afirman que, a mayor calidad de la dieta, la tasa de emisión de CH4 es menor (Kurihara et al., 1999). En este estudio se observó que las especies con mayor producción de CH4 fueron C. nlemfuensis, G. sepium y H. rosasinensis (37.70, 40.49, 47.64 ml/g MS incubado) (Tabla 4), lo cual es explicado por los bajos contenidos de TC y Sa. Las reducciones de la DEGMS y DEGMO para P. piscipula y L. leucocephala, G. ulmifolia y B. alicastrum (Tabla 4) podrían estar relacionadas con el contenido de TC (0.19%, 1.16%, 0.47% y 0.22% de MS); ya que los TC pueden formar complejos con la lignocelulosa y proteínas (Reed, 1995) e inhibir directamente los microorganismos celulolíticos y la actividad de enzimas fibrolíticas, reduciendo la digestibilidad y la fermentabilidad de los sustratos (Patra et al., 2017). La menor DEGMS y DEGMO en M. maximus y C. nlemfuensis está asociada al alto contenido de fibras estructurales (> 70% de la MS) (Archimède et al., 2011; Kennedy & Charmley, 2012; Kurihara et al., 1999), lo cual reduce la tasa de pasaje e incrementa la cantidad de metano por unidad de MS incubada. Por otra parte, G. sepium, T. diversifolia, M. alba y H. rosa-sinensis obtuvieron mayor DEGMS (610.33, 760.40, 800.16 y 800.66 g/kg de MS) y DEGMO (542.00, 698.90, 762.30 y 766.70 g/kg de MO), esto podría explicarse por las diferencias en la proporción de carbohidratos digeribles (RF y MF) y la baja proporción de celulosa (LF) (Hess et al., 2003).
Conclusión
La inclusión de L. leucocephala, P. piscipula, B. alicastrum y G. ulmifolia en las dietas para rumiantes representa una alternativa de alimentación debido a su aporte de proteína cruda y baja producción de gas en condiciones in vitro, lo que podría contribuir a la reducción de las emisiones de CH4. No obstante, cabe señalar que T. diversifolia y M. alba también se identificaron como una alternativa para la alimentación, debido a su alta degradabilidad. En este sentido, las combinaciones de estas arbóreas y arbustivas con pastos podrían representar una estrategia para mejorar el suministro de proteína durante las épocas de sequía en la región tropical.

