











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El uso de radiación gamma permite generar variabilidad intraespecífica y determinar el rango de radiosensibilidad (RRS) para el mejoramiento de plantas a través de mutagénesis. Las variaciones genéticas definen el potencial para evolucionar ante cambios ambientales y perpetuar la especie. En consecuencia, la generación de variabilidad es fundamental para el mejoramiento genético (Avendaño-Arrazate et al., 2012; Rosenberg, Shee, Frisch & Hastings, 2012). El Co60 es un isótopo radiactivo con alta capacidad ionizante que ha sido utilizado en mejoramiento genético de diferentes cultivos para inducir mutaciones (Krishna, Shivashankar & Nath, 1984; Ángeles-Espino et al., 2013).
Por otra parte, el pasto rosado [Melinis repens (Willd.) Zizka] es una gramínea nativa de Sudáfrica, considerada como una de las especies invasoras más agresivas en México. Esta especie fue introducida como planta ornamental a los Estados Unidos de Norte América por Florida entre 1870 y 1900 (David & Menges, 2011; Stokes, MacDonald, Adams, Langeland & Miller, 2011). Debido a su capacidad de dispersión y establecimiento, esta gramínea ha logrado desplazar especies nativas en diferentes ecosistemas del Desierto Chihuahuense (Díaz et al., 2012; Melgoza, Valladares, Mata & Pinedo, 2014). Otro factor de invasión es que, por el alto contenido en fibras y la baja digestibilidad que contiene el pasto rosado, es poco consumida por el ganado y fauna silvestre, por lo que al no ser depredada su establecimiento y dispersión son favorecidos (Valladares, Gianoli & Gomez, 2007). Lo anterior coloca a esta gramínea como un candidato para mejorar su valor nutritivo y a la vez, aprovechar su capacidad de establecimiento para control de erosión y rehabilitación de áreas degradadas. No obstante, en especies exóticas es difícil encontrar variabilidad genética. Para el caso del pasto rosado, el tiempo de su establecimiento en América es corto para efectos evolutivos (Nobel, 2009). En un estudio de caracterización morfológica y nutricional, Corrales-Lerma et al. (2017) encontraron que en el estado de Chihuahua el pasto rosado carece de variabilidad genética. En este sentido, la mutagénesis puede ser una alternativa viable para hacer mejoramiento genético en esta especie, debido a que esta técnica es capaz de inducir variabilidad mediante radiación gamma.
Al exponer un organismo a radiación ionizante y con dosis bajas, los impactos sobre la estructura celular o del ADN son mínimos, mientras que, con altas dosis, el genoma sufre múltiples impactos y puede ser causa de muerte (Songsri, Suriharn, Sanitchon, Srisawangwong & Kesmala, 2011; Thole et al., 2012; Olasupo, Ilori, Forster & Bado, 2016). Por esta razón, el primer paso para inducir mutagénesis efectiva con radiaciones ionizantes es determinar la dosis óptima de radiación a través de curvas de radiosensibilidad en diferentes variables. En semillas, la dosis letal media (DL50) se presenta con la concentración de radiación absorbida, donde se obtiene una sobrevivencia del 50% de germinación de la población expuesta (Ángeles-Espino et al., 2013; Álvarez-Holguín et al., 2018). Morela, González, Castroni, Díaz & Castro (2002) señalaron que en esta proporción existe mayor frecuencia de mutaciones útiles para programas de mejoramiento genético. En el mismo sentido, otros investigadores coinciden que, a través de radiación gamma en distintas especies vegetales, se puede determinar el RRS como punto de partida para realizar mejoramiento genético por mutagénesis (Lajonchere, Prieto & Sánchez, 1995; Songsri et al., 2011; Ángeles-Espino et al., 2013). Por otro lado, diversos estudios mencionan que otro parámetro con alta probabilidad para encontrar mutaciones efectivas, es el punto bajo la curva donde el crecimiento o desarrollo celular se reduce al 50% (GR50; Akgüm & Tosun, 2004; Khalil, Zamir & Ahmad, 2014). En este sentido, tanto la DL50 como la GR50 son parámetros conocidos como RRS efectivos para realizar mutagénesis inducida (Ariramana, Gnanamurthy, Dhanavelb, Bharathi & Murugan 2014; Álvarez-Holguín et al., 2018). En ambos casos se menciona que con dosis de radiación gamma inferiores al 50% de la curva, la modificación en la estructura del genoma es mínima o de bajo impacto (Ambavane, Gokhale, Sawardekar, & Sawantdesai, 2015; Álvarez-Holguín et al., 2018). Por el contrario, si la radiación supera la media en la curva, la estructura celular o ADN sufren mayor impacto de acuerdo con el incremento (Songsri et al., 2011; Thole et al., 2012).
Para el caso específico del pasto rosado se desconocen trabajos para inducir mutagénesis con radiación gamma, lo cual puede ser una alternativa para mejorar atributos que mejoren su valor nutricional, control de erosión de suelo y control de dispersión. Por lo anterior, el objetivo fue determinar la dosis óptima de radiación gamma para inducir mutaciones efectivas en pasto rosado mediante DL50 y GR50.
Materiales y Métodos
La duración del estudio fue de un año (octubre de 2014 a octubre de 2015) y se dividió en las siguientes etapas: recolección y análisis de semilla, radiación de semilla, prueba de germinación y desarrollo de plantas. En octubre de 2014 se recolectó semilla silvestre de pasto rosado en los municipios de Belisario Domínguez, Chihuahua y General Ángel Trías, del estado de Chihuahua. Se realizó una mezcla homogénea de la semilla recolectada de los tres municipios que incluyó 0.5 kg de semilla por municipio. La semilla fue colocada en bolsas de papel y almacenada. Seis meses después se realizaron pruebas de germinación y pureza para su análisis de calidad (Morales et al. 2012), el cual presentó un 35% de semilla pura viable con el 4% de humedad.
En marzo de 2015 se realizó el proceso de irradiación de la semilla, el cual se llevó a cabo en el Complejo Moscamed de Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (Senasica)-Secretaría de Agricultura y Desarrollo Rural (Sagarpa) en Metapa de Domínguez, Chiapas, México. Las dosis se determinaron mediante el sistema de dosimetría Gafchromic y una cámara de ionización RADCAL modelo Accudose (Monrovia, CA. USA). El irradiador fue un equipo panorámico Gamma Beam 127 MDS Nordion (Ottawa, ON, Canada) con fuente de almacenamiento de 50 g de Co60 en seco. Se evaluaron ocho dosis de radiación sobre la semilla: 10, 50, 100, 150, 200, 250, 300, 350 Gray (Gy) y 0 (T-0; tratamiento control). Después de irradiada la semilla se realizó una prueba de germinación estándar y crecimiento de plántula, la cual se llevó a cabo en el laboratorio de la Facultad de Zootecnia y Ecología (FZyE) de la Universidad Autónoma de Chihuahua (UACH). El desarrollo de plantas se realizó bajo condiciones de invernadero en las instalaciones de esta institución.
La prueba de germinación se llevó a cabo en agosto de 2015. Se utilizaron cajas Petri provistas de algodón y papel filtro, donde fueron colocadas 50 semillas por caja con cuatro repeticiones por dosis. Las cajas Petri se colocaron en una incubadora Precision Scientific modelo 6M (New Delhi, India) a temperatura de 28 °C ± 2 °C y humedad permanente. La prueba se realizó bajo un diseño completamente al azar con una duración de 15 días (d). Las variables evaluadas en esta etapa fueron: porcentaje de germinación (%GE), la cual se evaluó mediante conteos diarios. Se consideró semilla germinada cuando la plúmula o radícula fue igual o mayor a 5.0 mm de longitud. El índice de velocidad de germinación (Ivge) se estimó con base en la ecuación utilizada por González-Zertuche & Orozco-Segovia (1996).
Donde: Ivge = índice de velocidad de germinación; SG = número de semillas germinadas; t¡= día de registro de semillas germinadas. DG = total de días que duró la germinación, desde el momento de la siembra hasta el último día que se registró la última semilla germinada. Para la medición de LR y LP se dejaron crecer 12 plántulas por dosis de radiación durante 14 días con tres individuos por caja Petri.
El desarrollo de plantas para la evaluación en invernadero se realizó de julio a octubre de 2015. Se utilizaron las mismas dosis de semilla irradiada que en la prueba de germinación. Esta etapa se condujo bajo un diseño experimental en bloques completos al azar con 10 repeticiones por tratamiento. El criterio de bloqueo se tomó por la orientación y exposición a radiación solar de las plantas en el invernadero. Se sembraron 20 semillas de cada dosis por bolsa para asegurar emergencia y establecimiento; cada repetición consistió en una planta por bolsa establecida 21 d después de la siembra. Se usaron bolsas de polietileno negro de 25 cm de altura por 10 cm de diámetro, cada una con ocho perforaciones de 0.5 cm2 en la parte inferior. Las bolsas se llenaron a 21 cm - 22 cm con suelo franco-arenoso de origen aluvial. Se suministraron riegos periódicos a capacidad de campo en la maceta durante nueve semanas que duró la prueba (verano de 2015). Las variables evaluadas en esta prueba fueron: altura de follaje (AF), altura de planta (AP) y largo de hoja (LH). La AF se midió de la base de la corona a la lígula de la hoja bandera, hasta el final de la novena semana después de la emergencia. Para AP se midió la planta de la base de la corona hasta el ápice más alto de la inflorescencia en etapa de antesis temprana, al término de la semana nueve después de la emergencia. El LH fue medido al finalizar la semana cuatro después de la emergencia, se inició la medición de una hoja al azar de la parte central de la planta, lo cual se realizó durante seis semanas consecutivas. El índice de concentración de clorofila (ICC) se determinó con un medidor CCM-200® que detecta la fluorescencia de clorofila, en las mismas hojas seleccionadas para LH y AH.
En invernadero se monitorearon las variables ambientales: temperatura (T°), humedad relativa del aire (HR) y radiación solar (RdS) registradas de junio a septiembre de 2015. Para T° y HR se utilizaron sondas HMP60 y Vaisala Inc. (Woburn, MA, USA), respectivamente. Cada sensor fue sombreado y aspirado para lograr registros más precisos. Adicionalmente, se instaló un sensor de radiación de onda corta LI-200X, Li-Cor (Lincoln, NE, USA). Estas variables fueron registradas cada hora en un sistema de adquisición y almacenamiento de datos DataLogger CR1000, Logan, UT, USA. La T° media fue de 28 °C con una máxima de 48 °C (junio) y una mínima de 14 °C (septiembre). La media general para la HR fue de 26% con un valor mínimo de 15% en junio y un máximo del 47% en septiembre. La máxima RdS se presentó en julio con 760 W m-2 y el mes de agosto registró la menor radiación con 650 W m-2.
Los datos se analizaron mediante el procedimiento GLM del paquete estadístico Statistical Analysis System (SAS) 9.1.3 (2006). Se realizó análisis de tendencia con base en regresión lineal y cuadrática para comparar el mejor ajuste de acuerdo a la menor probabilidad de error (P value) para cada variable. Con base en la ecuación de regresión se determinó la DL50 para %GE. Con las ecuaciones resultantes de Ivge, LP, LR, AF, AP, LH e ICC, se determinó GR50 para cada una de estas variables. La dosis óptima de radiación gamma para mutaciones efectivas se calculó con medias ponderadas. Los valores obtenidos se ponderaron de acuerdo al peso de cada variable; 60% para DL50 en %GE y 40% para GR50, donde el 10% se asignó a Ivge, 5% a LP, 5% a LR, 5% a LH, 5% a ICC, 5% a AF y 5% para AP. Se incluyó Análisis de Varianza con comparación de medias. Se utilizó la prueba Dunnett para diferenciar el efecto de la dosis de radiación en cada variable del T-0. El factor de influencia fue la dosis de radiación y el nivel de significancia fue de α = 0.05.
Resultados
La Figura 1 presenta el comportamiento de las variables evaluadas en etapa de germinación. Para %GE se encontró efecto por dosis de radiación (p < 0.001). Los tratamientos 10 Gy, 50 Gy y 100 Gy no presentaron diferencia (p < 0.05) con T-0 en esta variable. Las dosis de radiación 150 Gy, 200 Gy, 250 Gy, 300 Gy y 350 Gy, presentaron menor (p < 0.001) %GE que T-0. Los resultados muestran una tendencia lineal descendente (p < 0.001). De acuerdo a la ecuación de regresión la DL50 en %GE se presentó a los 277 Gy (Figura 1a).
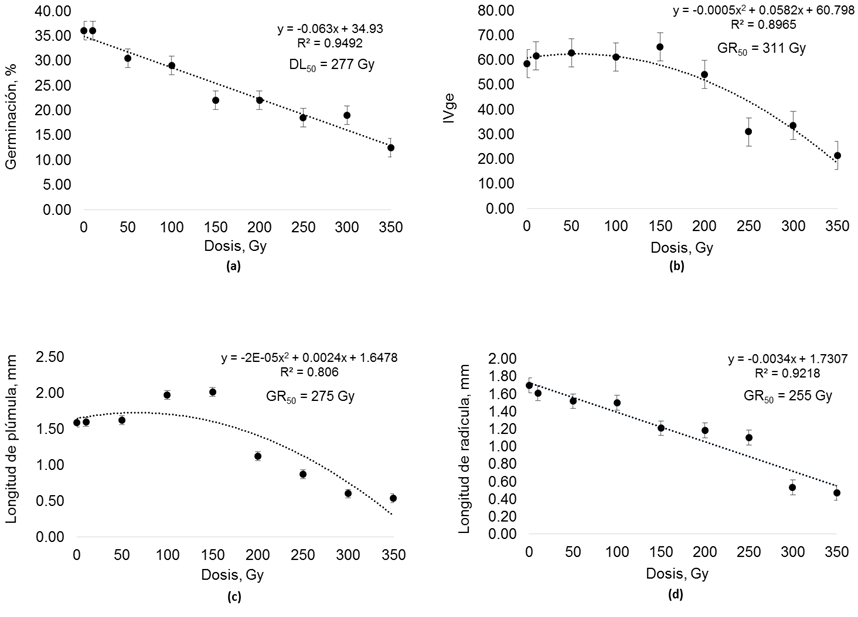
Fuente: Elaboración propia a partir de resultados del presente estudio.
Figura 1 Medias ± error estándar y curvas de radiosensibilidad (DL50 y GR50) para variables en etapa de germinación y plántula de pasto rosado (Melinis repens); con semilla expuesta a diferentes dosis de radiación gamma con cobalto 60. DL50 = dosis letal media, GR50 = reducción media de crecimiento, Gy = Gray, Ivge = índice de velocidad de germinación, escala 0-100.
El Ivge presentó efecto por radiación (p < 0.001). Las dosis 10 Gy, 50 Gy, 100 Gy, 150 Gy y 200 Gy no presentaron diferencia (p < 0.05) con T-0. En las dosis 250 Gy, 300 Gy y 350 Gy, la semilla fue más lenta (p < 0.001) para germinar que T-0. Esta variable mostró una tendencia cuadrática (p < 0.001) y la ecuación de regresión presentó la GR50 a los 311 Gy (Figura 1b).
En LP se encontró efecto (p < 0.001) por nivel de radiación. La LP de semilla germinada de los tratamientos 100 Gy y 150 Gy fue mayor (p < 0.05) al T-0. Es importante señalar que, con estas dosis de radiación gamma, se obtuvo una estimulación en el desarrollo de plúmula, ya que presentó mayor (p < 0.001) tamaño que el resto de los tratamientos. No obstante, de 200 Gy a 350 Gy la LP disminuyó categóricamente y presentó menor (p < 0.001) tamaño que en T-0. Esta variable mostró un comportamiento cuadrático (p < 0.001) y de acuerdo a la ecuación de regresión la GR50 se ubicó a los 275 Gy. Aunque en el presente estudio LP aumentó con dosis bajas de radiación, el comportamiento final de esta variable tendió a disminuir su tamaño con dosis altas de radiación (Figura 1c). Para LR se presentó efecto (p < 0.001) por nivel de radiación gamma. De los 150 Gy a los 350 Gy esta variable fue menor (p < 0.05) que en T-0. La LR mostró una tendencia lineal descendente (p < 0.001) y la ecuación indicó la GR50 a los 255 Gy (Figura 1d).
La Figura 2 muestra el comportamiento de las variables morfológicas de las plantas de semilla irradiada con Co60 evaluadas en invernadero. En AF se encontró efecto por dosis de radiación (p < 0.001) con tendencia cuadrática (p < 0.001). Con las dosis de 100 Gy y 150 Gy el follaje fue más alto que el T-0. A los 200 Gy esta variable comenzó a disminuir, pero fue hasta 350 Gy que presentó menor (p < 0.001) tamaño que T-0. En el experimento no se encontró la GR50, sin embargo, la ecuación indica la GR50 para AF a los 373 Gy (Figura 2a). Para AP se encontró efecto (p < 0.001) por radiación en la semilla. Las plantas de 350 Gy presentaron menor (p < 0.001) altura que el T-0. El resto de las dosis no presentó diferencia estadística con T-0 (p < 0.05). Aunque en el experimento no se encontró la GR50, la AP mostró una tendencia cuadrática (p < 0.001) y la ecuación presenta la GR50 a los 414 Gy (Figura 2b). Para LH se encontró efecto (p < 0.001) por semilla irradiada. Las hojas de las plantas de las dosis 250 Gy, 300 Gy y 350 Gy fueron más cortas (p < 0.001) que las de T-0. Los tratamientos de 10 Gy, 50 Gy, 100 Gy y 150 Gy no presentaron diferencias (p < 0.05) con T-0. La tendencia en esta variable fue cuadrática (p < 0.001). No obstante, en el experimento no se presentó la GR50 pero la ecuación estimó la GR50 a los 467 Gy (Figura 2c).
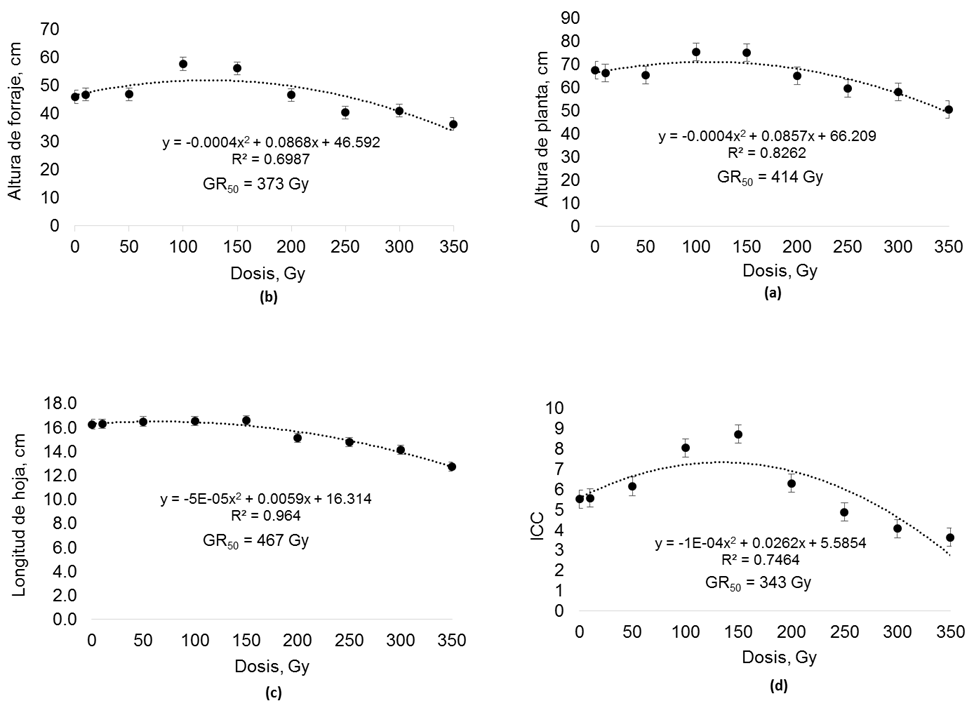
Fuente: Elaboración propia a partir de datos del presente estudio
Figura 2 Medias ± error estándar y curvas de radiosensibilidad (GR50) para variables morfológicas e índice de concentración de clorofila (ICC) de pasto rosado (Melinis repens) en etapa de planta; producidas de semilla expuesta a diferentes dosis de radiación gamma con cobalto 60. GR50 = reducción media de crecimiento, Gray= Gy.
La irradiación de semilla presentó efecto (p < 0.001) en el ICC de las plantas con una tendencia cuadrática (p < 0.001). En las dosis de 100 Gy y 150 Gy se registró mayor (p < 0.001) ICC que en T-0. Las plantas de 350 Gy registraron menor (p < 0.001) ICC que las de T-0. En esta variable no se obtuvo la GR50 con las dosis utilizadas en el experimento, sin embargo, la ecuación predice que la GR50 puede ser alcanzada a los 343 Gy (Figura 2d).
Discusión
Al incrementar la dosis de radiación gamma, los efectos se manifestaron con un decremento en el %GE. La DL50 en esta variable coincide con resultados de Ariramana et al. (2014), quienes en una prueba de germinación en semilla de frijol de palo (Cajanus cajan), leguminosa utilizada en el consumo humano que fue irradiada con Co60, encontraron la DL50 a 250 Gy en una prueba que duró 15 d. Bharathi, Gnanamurthy, Dhanavel, Murugan & Ariraman, (2013) al tratar una solanácea medicinal conocida como oroval o ashwagandha (Withania somnífera), encontraron la DL50 a los 250 Gy para %GE a los 15 d. Lo anterior puede ser un indicador para que semillas con características semejantes a estas especies, sean sometidas a estas dosis de radiación gamma. Así mismo, Melki & Marouani (2010) evaluaron semilla de trigo (Triticum aestivum) irradiada con Co60 y no encontraron diferencias en %GE con dosis de 0 Gy, 10 Gy, 20 Gy y 30 Gy. Con aplicación de otras fuentes de radiación como ultravioleta de onda corta (UV-C) y rayos X (RX), se intentó encontrar la DL50 en %GE para trigo (Triticum aestivum), girasol (Helianthus annuus), soya (Glycine max) y pasto buffel (Pennisetum ciliare), con dosis entre 0 Gy y 800 Gy (Foroughbakhch-Pournavab, 2015), sin embargo, la germinación de semilla en estas especies no fue afectada. Algunos estudios sugieren que la DL50 es el primer paso para el mejoramiento vegetal por mutagénesis, ya que determina el rango más efectivo de mutaciones (Ángeles-Espino, et al., 2013; Lajonchere et al., 1995; Morela et al., 2002; Songsri 2011;). Lo anterior, puede ser un indicador para que semillas ortodoxas con características similares a estas especies, sean consideradas para someterse a este rango de radiación gamma de Co60.
Con respecto al Ivge, estudios relacionados con esta variable reportan que, en semilla de soya, de la Fé, Romero, Ortiz & Ponce (2000) aplicó dosis entre 0 Gy y 480 Gy con radiación gamma de Co60 y reportó una tendencia cuadrática, ya que a los 280 Gy se obtuvo la mayor velocidad de germinación entre el tercer y noveno día. En otro estudio, Melki & Marouani (2010) midieron velocidad de germinación en trigo con semilla irradiada con Co60en dosis de 0 Gy, 10 Gy, 20 Gy y 30 Gy, sin obtener diferencias entre tratamientos.
El comportamiento decreciente en LR de este estudio fue similar a lo reportado por Ambavane et al. (2015), quienes irradiaron semilla de mijo africano (Eleusine coracana) con Co60 y encontraron que LP y LR disminuyeron a medida que la dosis de radiación aumentó. Para ambas variables estos investigadores reportaron la GR50 a los 500 Gy. Rafiuddin, Dahlan, Musa, Rasyid & Farid (2013) encontraron un efecto similar al irradiar semilla de tres variedades de maíz con Co60. En LP encontraron la DL50 a los 400 Gy para las variedades Bisma y Sukmaruga, mientras que en la variedad Lamuru la DL50 fue a los 500 Gy. En LR, estos mismos investigadores encontraron la GR50 a los 300 Gy en las tres variedades. El efecto de radiación en la expresión de LP y LR pudo deberse a que usualmente las radiaciones provocan desestabilidad en el desarrollo y división celular (Olasupo et al., 2016; Thole et al., 2012). De acuerdo con la intensidad radioactiva, en el presente estudio la expresión entre LP y LR fue diferente.
En las variables morfológicas evaluadas en invernadero no se encontró la GR50. No obstante, la radiación provocó diferencias notables. Se esperaría que a medida que la dosis de irradiación aumentara, estas variables disminuyeran su valor, sin embargo, las dosis de 100 Gy y 150 Gy parecen presentar una radio estimulación en incremento, con respecto a las plantas de semilla sin irradiar. Por otro lado, se presentó un decremento por arriba de los 200 Gy. En estudios relacionados, Scheer et al. (2011) utilizaron dosis entre 0 Gy y 200 Gy con Co60 para irradiar semilla en dos variedades de arroz mejoradas genéticamente y registradas como BRS-Fronteira y BRS-Querência. No se encontró la GR50, sin embargo, se reportó que la variedad BRS-Fronteira presentó un comportamiento cuadrático y que la mayor altura de planta fue con dosis de 100 Gy; mientras que la altura de planta en la variedad BRS-Querência presentó un comportamiento lineal descendente. Lemus, Méndez-Natera, Cedeño & Otahola-Gómez (2002) irradiaron semilla en dos variedades de frijol caupí (TC 9-6 y A-4) con dosis de 0 Gy a 750 Gy con Co60 y reportaron que de 0 Gy a 300 Gy el follaje no se vio afectado, no obstante, a partir de 400 Gy el número de hojas por planta disminuyó significativamente. Además, en altura de planta encontraron que a partir de 300 Gy fue donde ambas variedades disminuyeron significativamente.
El ICC se incrementó con las dosis de 100 Gy y 150 Gy, no obstante, al aumentar la intensidad de radiación gamma en semilla de pasto rosado el ICC disminuyó a partir de los 200 Gy. Aun así, 350 Gy no fueron suficientes para encontrar la DL50. Al respecto, Krishna et al. (1984) irradiaron semilla de pasto rhodes (Chloris gayana) con Co60 en dosis de 0 Gy a 900 Gy y reportaron que, al aumentar la intensidad de rayos gamma, se incrementó la cantidad de mutaciones en el espectro de la clorofila.
Al considerar las curvas de tendencia de cada una de las variables que determinaron DL50 y GR50, el resultado en la fórmula de medias ponderadas sugiere 304 Gy de radiación gamma como dosis óptima para inducir mutaciones efectivas en pasto rosado. Cabe mencionar que, aunque en las dosis de 100 Gy y 150 Gy se presentó aumento en ciertas características, no significa que la expresión en la primera generación de mutantes (M1) de pasto rosado sean las que el investigador busca. Por ejemplo, para el aprovechamiento del forraje por el ganado o fauna silvestre, el aumento en biomasa aérea para pasto rosado pudiera ser una característica no deseable, si la materia seca (MS) presenta un alto contenido de lignina. Una de las principales razones de la propagación de especies invasoras, es que no tienen suficientes depredadores. El pasto rosado ya está establecido en México y se sigue propagando sin control, por lo que al ser genéticamente modificado se debe tomar en cuenta que es una gramínea exótica invasora. De tal manera que los mutantes que se generen deberán presentar características que ayuden a su mayor consumo por el ganado y fauna silvestre, además de no causar daños a los ecosistemas.
Con los resultados de este estudio se propone el RRS con radiación gamma de Co60 alrededor de los 300 Gy, como dosis óptima para inducir mutaciones efectivas a futuro.
Conclusiones
Se determinó la dosis óptima de radiación del pasto rosado a los 304 Gy, con base en las curvas de radiosensibilidad en ocho variables a través de radiación gamma con Co60 en la semilla. Los resultados de este estudio sugieren un rango de radiación en semilla de esta especie para buscar mutaciones efectivas. Lo anterior, se refuerza con la variabilidad que mostraron las variables evaluadas en el rango de radiación, considerado en este estudio.
Estos resultados permiten conocer la radiosensibilidad del pasto rosado presente en poblaciones silvestres del estado de Chihuahua, para facilitar la inducción de nuevo material genético de interés agronómico y ecológico.

