











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El chile (Capsicum annuum L.) representa una de las actividades agrícolas más importantes en México. En el 2014 se produjeron 2 732 635.07 t con un valor de producción de más de 17 billones de pesos (Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación [Sagarpa], 2014). Dentro de los estados productores de chile en México, Chihuahua ocupa el primer lugar en producción con 722 708.61 t y el segundo lugar en valor de producción con más de 2951 millones de pesos (Sagarpa, 2014); sin embargo, el potencial productivo de este cultivo es afectado por diversas enfermedades (Guigón-López & González-González, 2001), siendo la mancha bacteriana, una de las más importantes. Esta enfermedad es causada por Xanthomonas campestris pv. vesicatoria (Nuez, Gil & Costa, 1996) y su mayor impacto es en la disminución de la calidad del fruto (Guigón-López & González-González, 2001; Velásquez-Valle & MedinaAguilar, 2005). El manejo de esta enfermedad es mediante el uso de semilla certificada y plántulas libres del patógeno; mediante rotación de cultivos y a través de compuestos a base de cobre o antibióticos (Jones & Permezny, 2003); sin embargo, estos productos son menos efectivos en condiciones cálidas y lluviosas o por que el patógeno se hace resistente a ellos (Nuez et al., 1996). Carrillo-Fasio et al. (2001) reportaron que X. campestris pv. vesicatoria presenta tolerancia a varias formulaciones de cobre y Minsavage, Canteros & Stall (1990) demostraron su resistencia a estreptomicina. Adicional a lo antes descrito, el uso inadecuado de estos productos químicos causa contaminación ambiental, afectando pérdida de microorganismos recicladores de nutrientes, cambio en las propiedades físico-químicas del suelo, contaminación de mantos freáticos, y pérdida de biodiversidad (Fleeger, Carman & Nisbet, 2003; Johnsen, Jacobsen, Torsvik & Sørensen, 2001; Reyes, Chaparro-Giraldo & Ávila, 2010). Para reducir los riesgos de contaminación ambiental, es importante aplicar otros métodos de control, siendo el control biológico una opción viable y ecológica. Las bacterias del género Streptomyces se consideran eficientes contra diversos fitopatógenos, incluyendo Fusarium oxysporum, Sclerotiniahomeocarpa y Rhizoctoniasolani (Gonzalez-Franco, Deobald, Spivak & Crawford, 2003; González-Franco & Robles-Hernández, 2009), Xanthomonas oryzae (Van et al., 2014) y Erwinia carotovora subsp. carotovora (Conn, Walker & Franco, 2008), entre otros, por lo que podrían ser una alternativa para el control de X. campestris pv. vesicatoria por la producción de antibióticos y enzimas involucradas en sus mecanismos de acción (González-Franco & Robles-Hernández, 2009). Por otra parte, Ganoderma lucidum es un basidiomiceto ampliamente utilizado en la medicina oriental por sus propiedades medicinales (Boh et al., 2004; Gao et al., 2002; Wasser & Weis, 1999; Wasser, 2002); así como en la supresión de patógenos de humanos (Chung & Jung, 1992; Smania, Delle, Yunes, Paulert & Smania, 2007). Los extractos bioactivos de este hongo han sido muy poco estudiados por su actividad antimicrobiana contra otras especies de bacterias fitopatógenas (Robles, 2004), por lo que también podrían ser una opción viable como agentes supresores de X. campestris pv. vesicatoria. El objetivo de este trabajo consistió en identificar el agente causal de la mancha bacteriana, determinar su patogenicidad en el cultivo de chile y evaluar su susceptibilidad a S. lydicus 5US-PDA8 y extractos bioactivos de G. lucidum en condiciones in vitro y de invernadero.
MATERIALES Y MÉTODOS
Recolección de muestras e identificacióndel agente causal
Se seleccionaron 18 lotes en seis localidades (Meoqui Estación Consuelo, Meoqui Lomas del Consuelo, Congregación Ortíz, Delicias Naica, Delicias Presa Francisco I. Madero y Cordillera Escuadra) de la región centro-sur del estado de Chihuahua y se colectaron 72 muestras foliares sintomáticas en bolsas de plástico previamente etiquetadas, las cuales se transportaron en una hielera a 4 0C para su procesamiento en el laboratorio de Microbiología Aplicada, Fitopatología y Fisiología de Poscosecha (MAFFP) de la Facultad de Ciencias Agrotecnológicas de la Universidad Autónoma de Chihuahua. El aislamiento se realizó en los medios de cultivo extracto de levadura-dextrosa-CaCO3 (YDC) y Tween (TM) y la identificación se hizo mediante pruebas bioquímicas de acuerdo a la metodología descrita por Schaad, Jones & Chun (2001).
Pruebas de patogenicidad
Los bioensayos de patogenicidad se establecieron en un diseño completamente al azar, con seis repeticiones, 10 tratamientos biológicos (MLC17, COR8, COR91, DN3, CC18, LMAM4, LIAM2, LI0AM4, LI3AM4 y L7AM3) y un control (solución salina al 0.25%). Se produjeron plántulas a partir de semilla de chile jalapeño, cultivar Híbrido Grande. Las plántulas se trasplantaron en macetas de polietileno de 20 cm x 20 cm con una mezcla de peat moss y arena tindalizada en una relación 3:1 y se colocaron en el invernadero para su establecimiento durante cinco días, realizando un riego a los tres días. Posteriormente, las plántulas se sometieron a un periodo de acondicionamiento en oscuridad a 26 ºC, con una humedad de 90% por 48 h en una cámara de crecimiento de plantas. Se prepararon soluciones bacterianas a una concentración de 1 x 106 UFC/ml a 1 x 107 UFC/ml. La concentración se ajustó en un espectrofotómetro UV (LaboMed Inc, modelo UV-2602), a una longitud de onda de 550 nm y una densidad óptica (DO) de 0.7 a 1.0 de acuerdo a Trigiano, Windham & Windham (2008). Las plántulas se inocularon por infiltración con las soluciones bacterianas en cuatro hojas por planta (Trigiano et al., 2008). En la hoja se hicieron dos orificios con una aguja estéril por el envés y luego se infiltraron 30 μL de cada tratamiento. Las plántulas control se inocularon únicamente con solución salina al 0.25%. Las plántulas se pusieron en un segundo periodo de acondicionamiento en las mismas condiciones descritas anteriormente y luego se pasaron al invernadero para su evaluación, las cuales se regaron cada tercer día y se fertilizaron cada 8 días con una solución nutritiva al 100% (Rubio et al., 2011) durante los 45 días que duró el experimento. A la aparición de los síntomas (siete días después de la inoculación [ddi]) se midieron las variables incidencia, severidad e índice de severidad. La incidencia (IE) se calculó utilizando la fórmula propuesta por Wolcan, Lori, Ronco, Mitidieri & Fernández (2001). La severidad y el índice de severidad se determinaron de acuerdo a Dugassa, Alten & Schönbeck (1996) con modificaciones, utilizando para este estudio una escala de cinco categorías (0, 1, 2, 3 y 4), donde: 0 = planta sana, 1 = de 1% a 25% de infección, 2 = de 25.1% a 50% de infección, 3 = de 50.1% a 75% de infección y 4 = de 75.1% a 100% de infección.
Los datos de las variables se procesaron con Microsoft Office Excel 365 y el porcentaje de infección se estimó utilizando Microsoft Picture Office Manager 2007, calculando el área de infección en función del área total de la hoja mediante el manejo de la imagen, la infección foliar se expresó en porcentaje.
Evaluación de la sensibilidad de los aislados bacterianos a los tratamientos biológicos in vitro
Los bioensayos para estas pruebas se realizaron bajo un diseño completamente al azar con tres repeticiones y tres tratamientos (extractos bioactivos [EsBsGl], S. lydicus 5US-PDA8 [St5US] y agar nutritivo como control [ANC]). Los experimentos se hicieron por triplicado. Streptomyces lydicus se sembró en agar nutritivo (AN) cubriendo 3.8 cm en una orilla de la caja y se incubó a 28 °C durante 8 días. Posteriormente, los aislados de X. campestris pv. vesicatoria se sembraron en líneas de 1.5 cm en sentido perpendicular al antagonista y con una separación entre ellas de 1.5 cm; el control se preparó de la misma forma pero sin el antagonista. Las cajas se incubaron a 27 ºC durante 48 h para su evaluación, el porcentaje de inhibición de los aislados bacterianos se determinó a las 24 h y 48 h. Por otro lado, los tratamientos con los extractos bioactivos (obtenidos por fermentación en caldo de papa dextrosa [PDB] casero que incluye la preparación de una infusión de papa a diferencia del PDB comercial) se prepararon de acuerdo a la metodología propuesta por Robles (2004) con modificaciones, utilizando el método de medios envenenados, donde se mezcla el agar nutritivo con los extractos bioactivos para preparar los tratamientos. Las cajas con los tratamientos respectivos se marcaron con nueve divisiones y se inocularon con 20 μL de las soluciones bacterianas preparadas de cada aislado de X. campestris pv. vesicatoria como se describió previamente. Las cajas se incubaron a 27 ºC por 48 h para su evaluación. Posteriormente, se determinó la inhibición del crecimiento a las 24 h y 48 h, utilizando los niveles de inhibición propuestos por Robles (2004): (−) = sin inhibición, (±) = inhibición parcial y (+) = inhibición total.
Evaluación de la sensibilidad de los aislados bacterianos a los tratamientos biológicos en condiciones de invernadero
El experimento se estableció sobre un diseño completamente al azar con tres repeticiones y 16 tratamientos, los cuales fueron: plantas inoculadas con cuatro de los aislados de X. campestris pv. vesicatoria más virulentos (MLC17, COR8, COR91 y CORDN3); plantas tratadas con extractos bioactivos de G. lucidum (EsBsGl) y sus respectivas combinaciones con los aislados bacterianos (EsBsGl+MLC17, EsBsGl+COR8, EsBsGl+COR91 y EsBsGl+CORDN3); plantas tratadas con S. lydicus 5US-PDA8 (St5US) y sus combinaciones con los aislados bacterianos (St5US+MLC17, St5US+COR8, St5US+COR91 y St5US+DN3); controles negativos: plantas no infiltradas (PS) y plantas infiltradas con solución salina (SS) al 0.25%.
Para la preparación de los tratamientos, S. lydicus 5US-PDA8 se creció en PDA a 28 0C hasta tener una abundante esporulación. Las esporas se colectaron y se resuspendieron en agitación por 5 min en 100 ml del medio líquido Crawford Medium (CM), conteniendo 0.3% (v/v) de quitina coloidal (Gonzalez-Franco et al., 2003). La solución de esporas se mezcló con 150 g de peat moss tindalizado en matraces de 2 L, los cuales se incubaron por 48 h a 28 0C y después se ajustó la población bacteriana a una concentración de 1 x 108 UFC/g. El peat moss inoculado se mezcló con arena tindalizada en una relación 3:1 y se colocó en bolsas de plástico de 20cm x 20 cm. Las plántulas de 15 días después de su germinación se trasplantaron en el sustrato, se colocaron en el invernadero, se regaron cada tercer día y se fertilizaron cada 8 días como se describió previamente.
Los tratamientos con extractos bioactivos se aplicaron en plantas con 20 hojas verdaderas establecidas en una mezcla de peat moss y arena (3:1). La aplicación de los extractos bioactivos se realizó con un atomizador de gota fina, cubriendo la superficie foliar hasta saturación. Dos días después, las plantas se sometieron a un acondicionamiento como se describió en las pruebas de patogenicidad. Las plantas correspondientes se inocularon con los cuatro aislados de X. campestris pv. vesicatoria por el método de infiltración con 20 μm de una solución bacteriana de 1 x 107 UFC/ml. La inoculación se realizó en cuatro hojas de cada planta como ya fue descrito previamente. Las plantas inoculadas recibieron un segundo periodo de acondicionamiento y luego se pasaron a invernadero para su evaluación. Se determinaron las variables infección foliar (IF), población bacteriana (PB), altura de la planta (A), biomasa (B) y clorofila (C). La IF se estimó con el software Picture office Manager 2007 a los 35 días después de inoculación (ddi). La PB se obtuvo al final del experimento (45 ddi) por el método de dilución seriada a partir de la maceración de un gramo de tejido foliar sintomático de cada repetición por tratamiento. Las variables A, B y C se determinaron a los 45 días después del trasplante (ddt). La altura de planta se midió desde el cuello hasta el ápice; la estimación de clorofila se realizó en 4 hojas de cada planta utilizando el medidor de clorofila SPAD-502 (Minolta); la biomasa se obtuvo secando las plantas en una estufa de aire forzado (Shel lab, modelo SGO6E) a 65 ºC hasta tener un peso constante.
Los datos se analizaron con el paquete estadístico Statgraphics centurion XV, realizando un análisis de varianza y una comparación de medias con Tukey (α = 0.05). Los análisis de correlación se hicieron con el software Microsoft Excel 365.
RESULTADOS
Identificación y pruebas de patogenicidad
De los 32 aislados seleccionados por sus características morfológicas en YDC y Medio Tween, 10 fueron identificados como X. campestris pv. vesicatoria. Los aislados presentaron crecimiento viscoso en YDC a 35 0C, hidrolizaron almidón y esculina, reaccionaron positivamente a la digestión de proteína, leche tornasol, utilización de glicerol, reacción ácida a partir de arabinosa y respondieron negativamente a la nucleación de hielo. Todos los aislados fueron patógenos de chile, con un 100% de incidencia y con diferentes niveles de virulencia. Las plantas control permanecieron sanas hasta el final del experimento. Los 10 aislados se ubicaron en la categoría 1 mayormente, pero COR91 y LIAM2 fueron los más infectivos; COR91 afectó el 66.6% de las plantas en la categoría 1 y el 33.3% en la categoría 4. LIAM2 infectó el 83.3% de las plantas en la categoría 1, y el 16.6% en la categoría 2. Se tuvo un índice de severidad de 25% a 47%, siendo mayores los aislados COR91 y LIAM2 (Tabla 1).
Tabla 1 Pruebas de patogenicidad de aislados de X. campestris pv. vesicatoria en plantas de chile jalapeño bajo condiciones de invernadero
|
Aislados bacterianos y control (solución salina) |
Incidencia (%) a) |
Severidad b) |
Índice de severidad (%) c) |
||||
| 0 | 1 | 2 | 3 | 4 | |||
| MLC17 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| COR8 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| COR91 | 100 | 0 | 4 | 0 | 0 | 2 | 47 |
| DN3 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| CC18 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| LMAM4 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| LIAM2 | 100 | 0 | 5 | 1 | 0 | 0 | 26 |
| LI0AM4 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| LI3AM4 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| L7AM3 | 100 | 0 | 6 | 0 | 0 | 0 | 25 |
| Solución salina | 0 | 6 | 0 | 0 | 0 | 0 | 0 |
a), c) Los datos de incidencia e índice de severidad son la media de 6 repeticiones.
b) Número de plantas infectadas en cada categoría (de 0 a 4), donde: 0 = 0% de infección, 1 = de 1% a 25% de infección, 2 = de 25.1% a 50% de infección, 3 = de 50.1% a 75% de infección y 4 = de 75.1% a 100% de infección.
Fuente: Elaboración propia.
Con relación a la sintomatología, las plantas infectadas con el aislado COR91, desarrollaron manchas irregulares grandes y clorosis general; LIAM2 y CC18 indujeron el desarrollo de manchas de tamaño medio, irregulares, rodeadas por un halo amarillo y una clorosis general ligera; LIAM4 causó el desarrollo de manchas pequeñas rodeadas de un halo amarillo y una clorosis ligera en la lámina foliar; MLC17, ocasionó la formación de manchas pequeñas y clorosis general en la hoja (Figura 1).
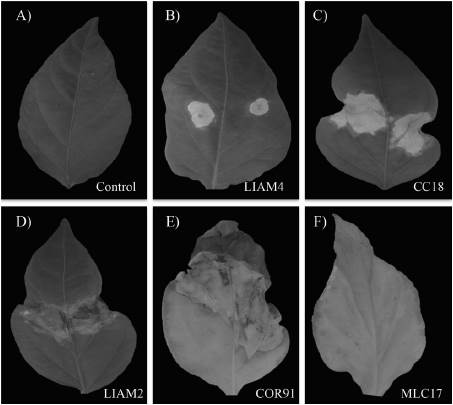
Fuente: Elaboración propia.
Figura 1 Síntomas inducidos por cinco aislados de X. campestris pv. vesicatoria en hojas de chile jalapeño a los 21 días después de inoculación. A) hoja sana tratada con solución salina al 0.25% (control); las imágenes B, C, D, E y F muestran las hojas inoculadas con cinco de los aislados bacterianos LIAM4, CC18, LIAM2, COR91 y MLC17, respectivamente con los síntomas característicos de la mancha bacteriana con distintos niveles de severidad.
Susceptibilidad de los aislados bacterianos a los tratamientos biológicos
Bioensayos in vitro
Todos los aislados bacterianos fueron susceptibles a los extractos bioactivos de G. lucidum (EsBsGl) y a S. lydicus 5US-PDA8 (St5US) durante los dos periodos de incubación (tabla 2). A las 24 h, St5US presentó un rango de inhibición de 9.2% a 100%. Este antagonista inhibió totalmente a cinco de los aislados bacterianos y uno de ellos en un 83.3%, mientras que EsBsGl presentaron un 100% de inhibición en todos los aislados bacterianos. A las 48 h, el grado de inhibición máximo de St5US bajó de 9.2% a 70.8%, siendo LI3AM4 el más inhibido y EsBsGl mantuvieron la inhibición total en ocho aislados y una inhibición parcial en los demás. El aislado MLC17 fue el menos afectado por St5US en ambos periodos de incubación, mientras que con los EsBsGl fue inhibido totalmente a las 24 h y parcialmente a las 48 h. Ambos tratamientos biológicos tuvieron un efecto similar en la inhibición del aislado LI3AM4 en los dos periodos de incubación. El control no causó ningún efecto en el crecimiento de los aislados bacterianos durante los dos periodos de incubación (Tabla 2). En la Figura 2, se presentan los resultados de la inhibición del crecimiento de nueve de los aislados bacterianos tratados con los EsBsGl a las 24 h y 48 h de incubación, en la cual se observa la inhibición total de todos los aislados bacterianos a las 24 h en comparación con el crecimiento vigoroso y normal de éstos en el tratamiento control (Figura 2a y 2b). A las 48 h de incubación, COR8, COR91, DN3, CC18, LMAM4, LIAM2, LI0AM4 y L7AM3 fueron inhibidos totalmente y MCL17 y LI3AM4 solo se inhibieron parcialmente, mostrando un crecimiento menos desarrollado y vigoroso que los mismos aislados en el tratamiento control (Figura 2c y 2d).
Tabla 2 Efecto de los extractos bioactivos de G. lucidum (EsBsGl) y S. lydicus 5USPDA8 (St5US) sobre la inhibición in vitro de los aislados de X. campestris pv. vesicatoria.
|
Aislados bacterianos y control (agarnutritivo) |
% de inhibición conSt5US a) | Inhibición delcrecimiento con EsBsGl b) | ||
| 24 h | 48 h | 24 h | 48 h | |
| MLC17 | 9.2 | 9.2 | + | ± |
| COR8 | 20.8 | 14.2 | + | ± |
| COR91 | 83.3 | 20.0 | + | + |
| DN3 | 22.5 | 20.0 | + | + |
| CC18 | 21.7 | 15.8 | + | + |
| LMAM4 | 100.0 | 58.3 | + | + |
| LIAM2 | 100.0 | 48.3 | + | + |
| LI0AM4 | 100.0 | 49.2 | + | + |
| LI3AM4 | 100.0 | 70.8 | + | ± |
| L7AM3 | 100.0 | 56.7 | + | + |
| Agar nutritivo | 0.0 | 0.0 | - | - |
a)Los datos son la media de tres repeticiones de tres experimentos independientes.
b)Simbología utilizada para indicar la inhibición del crecimiento de los aislados bacterianos: (+) = inhibición total, (±) = inhibición parcial y (−) = sin inhibición.
Fuente: Elaboración propia.
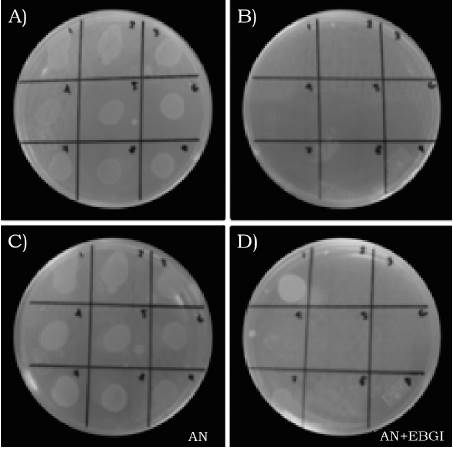
Fuente: Elaboración propia.
Figura 2 Inhibición del crecimiento de 9 aislados de X. campestris pv. vesicatoria con extractos bioactivos de G. lucidum a las 24 (A y B) y 48 horas (C y D) de incubación. AN = agar nutritivo, AN+EsBsGl = agar nutritivo mezclado con los extractos bioactivos de G. lucidum. Los números en las cajas corresponden al valor consecutivo de cada aislado bacteriano: 1 = COR8, 2 = COR91, 3 = DN3, 4 = CC18, 5 = LMAM4, 6 = LIAM2, 7 = LI0AM4, 8 = LI3AM4, 9 = L7AM3.
Bioensayos en invernadero
En la Tabla 3 se presentan los resultados del efecto de S. lydicus 5USPDA8 y los extractos bioactivos de G. lucidum sobre la reducción del porcentaje de infección y poblaciones bacterianas, así como en el desarrollo de plantas de chile jalapeño inoculadas con cuatro de los aislados de X. campestris pv. vesicatoria más virulentos en condiciones de invernadero. Todas las plantas inoculadas solo con los aislados de X. campestris pv. vesicatoria tuvieron los valores más altos de infección; sin embargo, en las plantas que fueron tratadas adicionalmente con los extractos bioactivos de G. lucidum, el porcentaje de infección bajó significativamente, especialmente con los tratamientos EsBsGl+COR8, EsBsGl+MLC17, EsBsGl+DN3 y EsBsGl+COR91 (1.7%, 1.9%, 2.6% y 2.8%, respectivamente). Por otro lado, en las plantas tratadas con S. lydicus 5USPDA8, solo se observó una ligera reducción en la infección foliar con los tratamientos St5US+COR8 y St5US+MLC17, los cuales fueron estadísticamente diferentes a los tratamientos COR8 y MLC17. En ninguna de las plantas del control negativo se presentó la infección. En cuanto a la población microbiana, los tratamientos que tuvieron un mayor efecto en la reducción significativa de esta variable fueron St5US+MLC17, St5US+COR8, EsBsGl+COR8, EsBsGl+MLC17. Aunque St5US+MLC17, St5US+COR8 fueron muy eficientes en la reducción de la población bacteriana, su efecto no fue significativo en la infección foliar, mientras que los tratamientos EsBsGl+COR8, EsBsGl+MLC17 bajaron la población bacteriana y la infección foliar. En los tratamientos EsBsGl+COR91 y EsBsGl+DN3, se observó un ligero incremento en el desarrollo bacteriano, en comparación con los tratamientos COR91 y DN3, pero disminuyeron significativamente la infección foliar, mientras que St5US+DN3, que también incrementó la concentración bacteriana, solo tuvo una ligera disminución en la infección foliar comparado con DN3. En altura de la planta, los tratamientos COR91 y EsBsGl+DN3 presentaron los valores más altos. En contenido de clorofila, St5US y PS fueron los más altos. En biomasa, EsBsGl+COR91 fue el más alto (tabla 3). En el análisis de correlación, en las plantas inoculadas únicamente con los aislados bacterianos, se observó que cuando la población bacteriana se incrementa, hay mayor infección foliar y menor contenido de clorofila (r = 0.939**, r = -0.6023*, respectivamente) y cuando se aumenta la infección foliar, también se reduce el contenido de clorofila (r = -0.6505*). En las plantas tratadas adicionalmente con extractos bioactivos o con S. lydicus 5USPDA8, solo se presentó una correlación débil en cada caso (r = 0.5851* y r = -0.5138*, respectivamente).
Tabla 3 Efecto de los extractos bioactivos de G. lucidum (EsBsGl) y S. lydicus 5US-PDA8 (St5US) sobre la reducción de infección y el desarrollo de plantas de chile jalapeño inoculadas con X. campestris pv. vesicatoria en invernadero.
| Tratamientosa) |
Infección foliar (%) [IF] |
Población bacteriana (UFC/ml) [PB] |
Altura de la planta (cm) [A] |
Clorofila (U-SPAD) [C] |
Biomasa (g) [B] |
| MLC17 | 4.3 a b) | 3.4x106b | 30.0 bc | 24.8 e | 5.3 abcd |
| COR8 | 4.0 ab | 2.9x106c | 25.0 de | 30.7 bcde | 5.3 bcde |
| COR91 | 3.4 abc | 1.8x106 e | 36.0 a | 29.6 bcde | 5.8 ab |
| DN3 | 3.5 abc | 2.5x106 d | 24.0 e | 29.4 bcde | 4.5 fg |
| EsBsGl+MLC17 | 1.9 d | 1.6x106 ef | 30.0 bc | 32.9 bcde | 5.0 def |
| EsBsGl+COR8 | 1.7 d | 1.3x106 f | 31.0 b | 32.0 bcde | 4.7 defg |
| EsBsGl+COR91 | 2.8 bcd | 3.0x106 c | 31.0 b | 28.5 cde | 6.1 a |
| EsBsGl+DN3 | 2.6 cd | 3.4x106 b | 34.3 a | 32.4 bcde | 5.7 abc |
| St5US+MLC17 | 3.6 abc | 0.2x106 h | 31.0 b | 32.4 bcde | 4.5 fg |
| St5US+COR8 | 3.2 abc | 0.7x106 g | 27.7 cd | 27.5 de | 4.1 g |
| St5US+COR91 | 3.5 abc | 1.5x106 ef | 28.3 bc | 32.9 bcde | 5.0 cdef |
| St5US+DN3 | 3.3 abc | 3.8x106 a | 29.7 bc | 24.6 e | 4.6 defg |
| PS | 0 e | 0 h | 27.3 cd | 38.7 ab | 4.8 defg |
| SS | 0 e | 0 h | 27.3 cd | 36.7 abcd | 4.6 defg |
| EsBsGl | 0 e | 0 h | 30.0 bc | 37.4 abc | 5.1 bcdef |
| St5US | 0 e | 0 h | 29.0 bc | 42.5 a | 4.6 defg |
| HSD | 1.2 | 0.3x106 | 3.3 | 9.5 | 0.76 |
a)Tratamientos: plantas inoculadas con 4 de los aislados de X. c. pv. vesicatoria más virulentos (MLC17, COR8, COR91y CORDN3); plantas tratadas con extractos bioactivos de G. lucidum (EsBsGl) y sus respectivas combinaciones con los aislados bacterianos (EsBsGl+MLC17, EsBsGl+COR8, EsBsGl+COR91 y EsBsGl+CORDN3); plantas tratadas con S. lydicus 5US-PDA8 (St5US) y sus combinaciones con los aislados bacterianos (St5US+MLC17, St5US+COR8, St5US+COR91 y St5US+DN3); controles negativos: plantas no infiltradas (PS), plantas infiltradas con solución salina al 0.25% (SS).
b)Los valores de las medias seguidas por la misma letra dentro de cada columna no son estadísticamente diferentes (Tukey α = 0.05, n = 3).
Fuente: Elaboración propia.
DISCUSIÓN
Los resultados de incidencia obtenidos en este trabajo fueron superiores (100%) a los reportados en la literatura. Velásquez-Valle & Medina-Aguilar (2005) registraron una incidencia del 50%, Mirik, Aysan & Cinar, (2005) una del 33% y Black, Seal, Abubakar, NonoWomdim & Swai (2001) una del 5%. Con relación a la severidad, en este estudio se obtuvo un valor promedio de 36%, mayor al encontrado por Velásquez-Valle & Medina-Aguilar (2005). La variación de incidencia y severidad de la mancha bacteriana podría estar relacionada con las condiciones ambientales y la genética del patógeno. Araújo et al. (2011) reportaron que la incidencia de la mancha bacteriana del chile se incrementa en condiciones de alta humedad relativa y temperaturas frescas, pero que, en situaciones de sequía y altas temperaturas por periodos largos de tiempo, esta se reduce. O’Garro (1998) y Gore & O’Garro (1999) señalan que X. campestris pv. campestris tiene varias razas fisiológicas con diferentes niveles de virulencia que influyen en el desarrollo de la mancha bacteriana.
Los síntomas causados por los aislados bacterianos en plantas de chile en este estudio fueron similares a los reportados por Gore & O’Garro (1999) y Potnis et al. (2015), quienes también observaron manchas irregulares en hojas, con un aspecto seco y de color marrón, rodeadas por un halo amarillo, clorosis general en algunos casos y caída de hoja debido a infecciones severas.
Los tratamientos biológicos tuvieron una fuerte inhibición del patógeno bajo condiciones in vitro y en invernadero, reflejando una menor infección foliar y un mejor desempeño de las pantas de chile jalapeño; sin embargo, los mejores resultados se registraron con los extractos bioactivos de G. lucidum. Las especies de Streptomyces se han estudiado ampliamente por su capacidad de suprimir el crecimiento y desarrollo de fitopatógenos, particularmente de hongos, debido principalmente a la producción de una gran variedad de metabolitos secundarios con actividad antifúngica y enzimas hidrolíticas que favorecen el micoparasitismo (Barakate, Ouhdouch, Oufdou & Beaulieu, 2002; Doumbou, Hamby-Slove, Crawford & Beaulieu, 2001; Gonzalez-Franco et al., 2003; González-Franco & Robles-Hernández, 2009; Rosas, 2007; Strap & Crawford, 2006); sin embargo, su efecto inhibitorio también se ha reportado contra algunas bacterias fitopatógenas (Conn et al., 2008; Van et al., 2014). En este estudio, Streptomyces presentó un rango de inhibición del 9% al 100% de los 10 aislados bacterianos en condiciones in vitro. El rango de inhibición podría estar relacionado con la variación genética del patógeno (Gore & O’Garro, 1999) y por los mecanismos de acción de Streptomyces (Cao, Qiu, You, Tan & Zhou, 2005; Hiltunen, Linfield & White, 1995; Singh, Shin, Park & Chung, 1999). En invernadero, Streptomyces redujo el porcentaje de infección y las poblaciones bacterianas de dos de los aislados bacterianos más virulentos. Estos efectos podrían estar asociados con otros mecanismos de acción, ya que algunas especies de Streptomyces tienen la capacidad de inducir resistencia sistémica en planta a diferentes enfermedades cuando se asocian con el sistema radicular de la planta. Este mecanismo se expresa cuando los microorganismos antagonistas y fitopatógenos se aplican por separado en tiempo y espacio (Kloepper & Beauchamp, 1992), como ocurrió en este estudio. La resistencia sistémica inducida puede ser efectiva contra una gran diversidad de microorganismos fitopatógenos, incluyendo bacterias, hongos y virus (Kloepper & Beauchamp, 1992; Doornbos, Van Loon & Bakker, 2012). Conn et al. (2008) demostraron que Streptomyces sp. es capaz de inducir resistencia sistémica contra Fusarium oxysporum y Erwinia carotovora subsp. carotovora en Arabidopsis thaliana. En estas condiciones, las plantas activan genes de resistencia que codifican para varios tipos de enzimas involucrados en los mecanismos de defensa, destacando las peroxidasas (E.C. 1.11.1.7), polifenol oxidasas (E.C. 1.10.3.1), fenilalanina amonio-liasas (4.3.1.24), entre otras. Estas enzimas forman parte del metabolismo secundario de las plantas, del cual se obtienen metabolitos que ayudan a la formación de estructuras de defensa (Doornbos et al., 2012). La enzima fenilalanina amonio-liasa y compuestos fenólicos fueron los principales metabolitos involucrados en la supresión de Sclerotium rolfsii en plantas de garbanzo tratadas con Streptomyces (Basha, Sarma, Singh, Annapurna & Singh, 2006).
Tabla 4 Correlación entre las variables de infección y vegetativas de plantas de chile inoculadas con cuatro aislados de X. campestris pv. vesicatoria y tratadas con S. lydicus 5US-PDA8 o extractos bioactivos de G. lucidum en condiciones de invernadero.
| Plantas infectadas con X. campestris pv. vesicatoria | |||||
| Aa) | IF | C | PB | B | |
| A | 1.0000b) | ||||
| IF | -0.2128 | 1.0000 | |||
| C | -0.2157 | -0.6505* | 1.0000 | ||
| PB | -0.4796 | 0.9139** - | 0.6023* | 1.0000 | |
| B | 0.6065*c) | 0.2606 | -0.1608 | -0.0059 | 1.0000 |
| Plantas infectadas con X. campestris pv. vesicatoria y tratadas con S. lydicus 5US-PDA8 | |||||
| A | IF | C | PB | B | |
| A | 1.0000 | ||||
| IF | 0.4522 | 1.0000 | |||
| C | 0.0552 | 0.2415 | 1.0000 | ||
| PB | -0.0172 | -0.2189 | -0.5138* | 1.0000 | |
| B | -0.1259 | 0.1547 | 0.1365 | 0.3010 | 1.0000 |
| Plantas infectadas con X. campestris pv. vesicatoria ytratadas con extractos bioactivos | |||||
| A | IF | C | PB | B | |
| A | 1.0000 | ||||
| IF | 0.1025 | 1.0000 | |||
| C | 0.2071 | -0.1363 | 1.0000 | ||
| PB | 0.0603 | 0.2929 | 0.4339 | 1.0000 | |
| B | 0.3093 | 0.4527 | 0.2028 | 0.5851* | 1.0000 |
a) Variables: altura de planta (A); infección foliar (IF); clorofila (C); población bacteriana (PB); biomasa (B).
b) El análisis se realizó utilizando los datos en conjunto a partir de los tratamientos con tres repeticiones.
c) Niveles de correlación (positiva o negativa) de acuerdo a Gutiérrez-Banegas (2012): correlación muy débil de 0.0 a 0.5, correlación débil de 0.51 a 0.7 *, correlación fuerte de 0.71 a 1.0**.
Fuente: Elaboración propia.
Por otra parte, G. lucidum ha sido estudiado principalmente para su uso en la medicina oriental (Ameri, Vaidya & Deokule, 2011; Chung & Jung, 1992; Smania et al., 2007), del cual han sido identificados cerca de 400 compuestos con propiedades medicinales (Wasser, 2002; Wasser & Weis, 1999), siendo los triterpenos y polisacáridos los constituyentes más importantes (Boh et al., 2004); sin embargo, las aplicaciones de este hongo en agricultura son escasas. Robles (2004) reportó por primera vez la actividad antimicrobiana de extractos bioactivos a partir de G. lucidum contra diversos microorganismos fitopatógenos, incluyendo bacterias, hongos y nematodos. Palacios et al. (2011) reportaron que los filtrados de Ganoderma sp. inhibieron el crecimiento micelial de Phytophthora nicotianae. La actividad antimicrobiana de los extractos bioactivos de G. lucidum encontrada en este estudio, podría estar relacionada con la producción de metabolitos antimicrobianos. Robles (2004) identificó un nuevo polisacárido de bajo peso molecular asociado con la inhibición de varias especies de bacterias fitopatógenas y Wasser & Weis (1999) reportaron polisacáridos constituidos de β-D-glucanos, con un peso molecular de 4 x 105 a 1 x 106 Da, pero solo con aplicaciones en medicina como agentes inmunomoduladores y antitumorales (Gao et al., 2002). Contrario a estos polisacáridos, los reportados por Robles (2004) tienen una potente actividad contra bacterias fitopatógenas. Los mecanismos de acción de estos extractos bioactivos en la supresión del crecimiento de X. campestris pv. vesicatoria podrían estar relacionados con la presencia de compuestos que inhiben la actividad ribosomal del microorganismo, como acurre con la proteína lyophyllin producida por Lyophyllum shimeji (Lam & Ng, 2001); estos extractos también podrían tener compuestos que inactivan la respiración microbiana como se ha reportado con varios tipos de estrobilurinas producidas por otros basidiomicetos como Strobilurus tenacellus, (Anke & Oberwinkler, 1977), Xerula longipes (Anke, Besl, Mocek & Steglich, 1983) y Crepidotus falvotomentosus (Weber, Anke, Steffan & Steglich, 1990). Es posible que los extractos bioactivos de G. lucidum también posean compuestos inductores de resistencia en planta como se reportó con la strobilurina F500 que causó un incremento en la concentración de ácido salicílico y varios tipos de proteínas involucradas en los mecanismos de defensa en plantas de tabaco (Herms, Seehaus, Koehle & Conrath, 2002).
CONCLUSIONES
De los 32 aislados seleccionados por sus características morfológicas, 10 fueron identificados mediante pruebas bioquímicas como X. campestris pv. vesicatoria. Todos los aislados identificados fueron patógenos de plantas de chile; COR91 y LIAM2 fueron los más virulentos. Todos los aislados fueron susceptibles a los tratamientos biológicos, pero los extractos bioactivos de G. lucidum registraron los mejores resultados, inhibiendo en su totalidad a todos los aislados en condiciones in vitro; mientras que en invernadero redujo la infección foliar y las poblaciones bacterianas, favoreciendo el desarrollo de la planta. Este es el primer estudio que muestra la efectividad de S. lydicus 5US-PDA8 y los extractos bioactivos de G. lucidum en el control de X. campestris pv. vesicatoria en condiciones in vitro y de invernadero; sin embargo, es necesario explorar más en los mecanismos de acción de los extractos bioactivos y evaluar su efectividad en condiciones de campo.

