











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
El cambio climático es uno de los desafíos más complejos de comienzos de nuestro siglo. Ningún país está inmune. Ningún país puede, por sí solo, afrontar los desafíos interconectados que plantea el cambio climático, entre los que se incluyen decisiones políticas controvertidas, un cambio tecnológico y consecuencias mundiales de gran alcance. El cambio climático está alterando la estructura y funcionalidad del ecosistema que afecta la disponibilidad de recursos ecológicos, interacciones entre ecosistemas y el clima, así como la economía que depende de los ecosistemas. Es difícil comprender el efecto de la variabilidad del cambio climático y el cambio que sufren los ecosistemas, la oferta de bienes y servicios, las consecuencias de las acciones humanas hacia el ambiente.
El Panel Intergubernamental de expertos sobre el Cambio Climático (PICC, 2014) menciona que en los últimos 100 años la temperatura superficial global promedio se ha incrementado entre 0.65 °C y 1.6 °C. Por lo que la temperatura superficial podría ascender entre 0.3 °C y 4.8 °C para finales de siglo, (PICC, 2014). Como resultado, se han reportado cambios en la distribución y composición de comunidades de especies marinas en zonas templadas y subtropicales (Albouy, Guilhaumon, Araujo, Mouillot & Leprieur, 2012; Ayala-Bocos, Reyes-Bonilla, Calderón-Aguilera, Herrero-Pérezrul & González-Espinosa, 2016; Perry, Low, Ellis & Reynolds, 2005; Pont, López, Carrel, Rogers & Haidvogl, 2015). Incluso en ese informe del PICC se lee textualmente que “Muchas especies terrestres, dulceacuícolas y marinas han modificado sus áreas de distribución geográfica, actividades estacionales, pautas migratorias, abundancias e interacciones con otras especies en respuesta al cambio climático en curso” (PICC, 2014).
El cambio climático ha roto el equilibrio natural y se ha modificado el estado de salud de las diferentes especies comerciales como los pargos (Lutjanus guttatus, L. peru y L. colorado principalmente), botete (Sphoeroides annulatus), roncacho (Haeulopsis leuciscus), curvina (Cynoscion sp.), robalo (Eucinostomus sp.). Por lo anterior, es importante evaluar causas y efectos del cambio climático sobre la incidencia y prevalencia de los eventos interanuales en su fase cálida como El Niño y su fase fría como La Niña, el cambio en las condiciones ambientales de las corrientes oceánicas, así como del hábitat, que ha repercutido además en una disminución de las pesquerías de importancia comercial y en producciones acuícolas.
El Golfo de California es la zona pesquera más importante de México, aproximadamente 80% de las capturas pesqueras del Pacífico mexicano se generan, o al menos se desembarcan, en costas de Baja California, Baja California Sur, Sonora, Sinaloa y Nayarit. El Golfo de California cuenta con la mayor población de pescadores ribereños y el mayor número de organizaciones pesqueras ribereñas del Pacífico mexicano. En este mar inició, en 2002, la pesquería de almeja de sifón Panopea spp. con una producción de 49 t y para 2014 las capturas declaradas fueron de 6951 t (Conapesca, 2016), que comprende ambas especies P. generosa y P. globosa. Sin embargo, P. globosa contribuye con más del 85% de las capturas totales. A pesar de su importancia comercial son pocos los trabajos publicados de esta especie: Aragón-Noriega, Alcántara-Razo, Calderón-Aguilera & Sánchez-Fourcade (2012) presentaron una revisión del conocimiento generado hasta ese momento y hacen mención a la necesidad de explorar nuevas áreas de pesca. Por esa razón la motivación de este trabajo fue, primeramente, la de localizar las áreas de distribución potencial de la especie con fines de explotación en el Golfo de California y posteriormente evaluar el efecto del cambio climático sobre su distribución futura partiendo de herramientas como evaluación de hábitat favorable.
En los últimos años se ha generalizado una herramienta que permite analizar objetivamente los patrones espaciales de presencia de organismos: los modelos de distribución de especies. Estos modelos se basan en procedimientos estadísticos y cartográficos que, partiendo de datos reales de presencia, permiten inferir zonas potencialmente idóneas en función de sus características ambientales. La idoneidad no es más que la relación matemática o estadística entre la distribución real conocida y un conjunto de variables independientes que se usan como indicadores. Estás variables suelen ser térmica, química y biológicas y condiciones batimétricas, se espera que con algunas de ellas, individualmente o en combinación, se puedan definir los parámetros ambientales que delimiten las condiciones favorables para la presencia de la especie. Uno de ellos es el modelo propuesto por Phillips, Anderson & Schapire (2006) de máxima entropía (MaxEnt). Este modelo estima la distribución potencial del hábitat de la especie con base en la localización de la distribución de MaxEnt (más cercana y uniforme), considerando que los valores esperados para cada una de las características analizadas bajo esta distribución estimada, coinciden en su valor empírico promedio. Sin embargo, el modelo no tiene en cuenta las interacciones biológicas entre las especies, tales como los fenómenos de competencia o sinergia. No considera la posibilidad de la especie de llegar a todos los sitios donde podría vivir (no incluye, por ejemplo, parámetros que describan la capacidad de dispersión).
El estudio se inició con las siguientes preguntas: ¿Cuál es la probabilidad de presencia de Panopea globosa en el Golfo de California? ¿El cambio climático afectará la distribución de Panopea globosa en el Golfo de California? Por lo tanto, el objetivo del presente estudio fue estimar el efecto del cambio climático en la distribución futura (al año 2050) de Panopea globosa en el Golfo de California.
Métodos
Distribución de Panopea globosa
Para determinar el hábitat potencial se utilizaron los datos provenientes de pesca comercial de P. globosa en el Golfo de California (San Felipe, Puerto Peñasco y la parte central de Sonora) y los registros de la lista de verificación de la macrofauna del Golfo de California (Aragón-Noriega et al., 2012; Hendrickx, Brusca & Findley, 2005). La Figura 1 presenta sitios donde existen actualmente permisos de pesca comercial o de fomento de las dos especies de almeja de sifón. Para P. generosa básicamente en el Pacífico mexicano y para P. globosa en el interior de Bahía Magdalena y el Golfo de California. Los sitios de pesca representan importancia social y económica ya que es una especie de alto valor comercial y que genera importantes empleos directos e indirectos (Aragón Noriega et al., 2012).

Datos ambientales
Se seleccionaron las variables ambientales por la importancia que puede tener en la de distribución de P. globosa y con sujeción a la disponibilidad de los datos. Por lo tanto, se usaron 12 variables ambientales que representaban las condiciones térmicas, químicas, biológicas y batimétricas derivadas de diferentes fuentes (Tabla 1). Se obtuvieron cuadrículas que representan el promedio total de la temperatura máxima y mínima de la superficie del mar por medio de imágenes de satélite mensuales de 2006 a 2008 con una resolución espacial de un grado. Para una mayor resolución se remuestrearon los datos ambientales en una red de 1 km, con un modelo de interpolación bilateral.
Tabla 1 Variables ambientales utilizadas para predecir el hábitat favorable de Panopea globosa.
| Variable | Unidad | Fuente |
|---|---|---|
| Temperatura superficial del mar | °C | National Oceanic and Atmospheric Administration (NOAA) |
| Temperatura máxima | °C | National Oceanic and Atmospheric Administration (NOAA) |
| Temperatura mínima | °C | National Oceanic and Atmospheric Administration (NOAA) |
| Concentraciones de Clorofila-α | mg/m3 | National Geophysical Data Center (NASA) |
| Salinidad | Pss | World Ocean Atlas (WOA) |
| Alcalinidad | µmol/kg | Ocean Data View (ODV) |
| Carbono inorgánico disuelto | µmol/kg | World Ocean Atlas (WOA) |
| Nitrato | µmol L | World Ocean Atlas (WOA) |
| Fosfatos | µmol L | World Ocean Atlas (WOA) |
| Silicatos | µmol L | World Ocean Atlas (WOA) |
| Oxígeno disuelto | mL L | World Ocean Atlas (WOA) |
| Batimetría | m | General Bathymetric Chart of the Oceans (GEBCO) |
Fuente: Elaboración propia.
Modelado con MaxEnt
Para el desarrollo del modelado alternativo con base en el hábitat favorable se utilizó el programa MaxEnt 3.2.1. (Phillips et al., 2006). Para el desarrollo de este tipo de modelos se utilizan una serie de registros espaciales (latitud-longitud) en combinación con una serie de características ambientales. Para el modelado de la distribución de las especies, las localidades con presencia sirven como los puntos de muestreo, la región geográfica de interés es el espacio donde la distribución será definida y las características son las variables ambientales (Phillips et al., 2006). Este algoritmo calcula la distribución de probabilidad de presencia de la especie que más se extendió teniendo en cuenta las limitaciones derivadas de los datos disponibles y expresa la idoneidad del hábitat para las especies en función de variables ambientales (Phillips et al., 2006). El modelo proporciona una superficie de probabilidad en el espacio que representa un nicho ecológico (la idoneidad del hábitat). Los valores de logística de salida van desde 0 (hábitat inadecuado) a 1 (hábitat óptimo), cuando el valor de la función de distribución es > 0.5 indica que las condiciones son favorables para la presencia de la especie, valores contrarios (< 0.5) significan que la celda no produce condiciones necesarias para el establecimiento de las especies (Pearson, Raxworthy, Nakamura & Townsend-Peterson, 2007; Phillips & Dudík, 2008). Algunas de las ventajas de MaxEnt son que requiere datos de presencia ambientales por separado (Phillips et al., 2006).
Efecto incremento de temperatura sobre la distribución
Para evaluar si el aumento en la temperatura superficial del mar tiene algún efecto en los nichos ecológicos y distribución potencial de P. globosa, se incorporaron al modelo MaxEnt mapas de tendencias de temperatura estimadas para 2050. Para mejorar la coherencia entre los datos de entrada y resultados, se ajustaron los parámetros y las constantes del modelo (Rykiel, 1996). Salida de logística, un valor iteración máximo de 1000 y el modelo por defecto se usan los parámetros (un umbral de convergencia de 10 - 5 y regularización automática con un valor de 4.10). Estos ajustes han logrado mostrar un buen rendimiento (Phillips & Dudik, 2008) y, en general, la reciente comparación entre varias técnicas de predicción de la distribución de especies muestra que MaxEnt fue uno de los métodos más eficaces con un ajuste particularmente bueno cuando el conjunto de datos analizados cuenta con un número pequeño de registros de presencia, como en este caso (Pearson et al., 2007).
Para estimar la velocidad de cambio en la temperatura superficial del Golfo de California, se utilizaron registros satelitales generados por la Agencia Nacional Oceanográfica y Atmosférica de los Estados Unidos (NOAA, por sus siglas en inglés). Como línea base, se seleccionaron registros del 1984 al 2008 el cual es empleado por la NOAA y otras agencias para evaluar efectos del calentamiento sobre corales arrecifales y otros organismos (Eakin, Lough & Heron, 2009). Con un análisis de regresión lineal simple y tomando como factor el año, se estimó la tasa anual de cambio termal (equivalente a la pendiente de la recta). La información generada se incluyó en un sistema de información geográfica y se interpoló usando la extensión Geostatistical Analyst del programa ArcMap 9.3 para construir mapas prospecto de las tendencias de elevación de temperatura en 2050.
Resultados
Distribución actual
El modelo de MaxEnt predijo un hábitat potencial adecuado para P. globosa con altas tasas de éxito (Area Under the Curve [AUC] = 0.995) que indica un buen ajuste del modelo. Las curvas de respuesta para cada una de las variables y los rangos óptimos de distribución de las especies (Figura 2a-c) ilustran la predicción de logística de las mejores condiciones del hábitat o áreas donde es más probable que se encuentre la especie. Los resultados muestran una relación positiva entre P. globosa en aguas poco profundas (<100 m) y las áreas productivas con altas concentraciones de clorofila-a (>8 mg/m3) y nitrato (1.5 - 2. Mol/l). Estas son las condiciones típicas del Alto Golfo de California y el Golfo de California. De las 12 variables, la batimetría muestra la información más útil porque si la excluimos disminuye la ganancia total del modelo (Figura 2d).
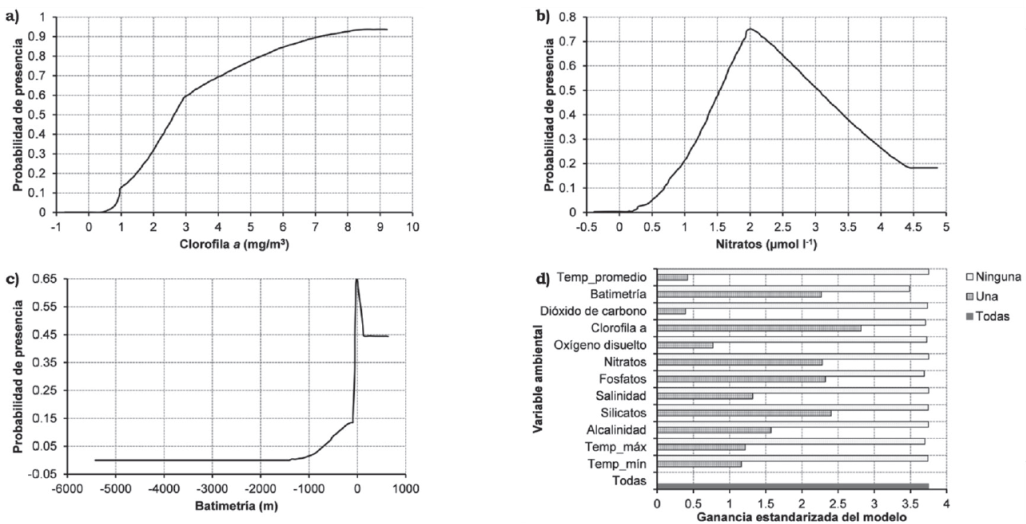
Fuente: Elaboración propia.
Figura 2 Curvas de respuesta entre las variables de mayor contribución y la probabilidad de ocurrencia de P. globosa (A-C). Resultados de la prueba “jack-knife” para estimar la importancia de cada variable (D).
Los resultados del modelo indican que el hábitat más adecuado para P. globosa está restringido dentro del Golfo de California y asociado a las áreas de alta productividad; por lo tanto se esperaría que el hábitat más adecuado se encuentre en las zonas del Alto Golfo de California, Guaymas, Sonora y la costa de Santa Rosalía y la Isla San Marcos en Baja California Sur.
Distribución en 2050
El cambio de la temperatura del mar en el Golfo de California no es homogénea; Bahía de Banderas y la porción Sur de Sonora presentan tasas de aumento de temperatura >0.2 °C al año, mientras que en el Alto Golfo de California la tasa de cambio es más estable con tendencia a disminuir la temperatura promedio en el año 2050. Bajo el supuesto del modelo empleado para estimar las tendencias de temperatura en los próximos años (2050), la diferencia de la temperatura superficial del mar en el Golfo de California con respecto a la actual podrá ser de más de 1 °C. Al comparar las curvas de respuesta de la temperatura promedio (obtenida del primer modelo) y la temperatura en 2050, se observa que el aumento de temperatura reduce el rango óptimo de mayor probabilidad de presencia de la especie hacia zonas más cálidas (24.5 °C - 25.8 °C). Estos resultados se observan también en el mapa de hábitat favorable (Figura 3) donde la distribución de P. globosa se restringe un poco más al norte del Golfo de California y desaparece en la porción norte de Sinaloa. En el Alto Golfo hay un aumento en la probabilidad de ocurrencia principalmente en el Golfo de Santa Clara y San Felipe (0.80 y 0.60 respectivamente). En San Felipe es donde se ha instalado mayor infraestructura pesquera de la especie y por tanto mayor beneficio social y económico. Los sitios de mayor probabilidad de ocurrencia en Sonora se reducen ligeramente hacia la costa sur y en Santa Rosalía e Isla San Marcos en donde las probabilidades disminuyen de 0.70 a 0.04.
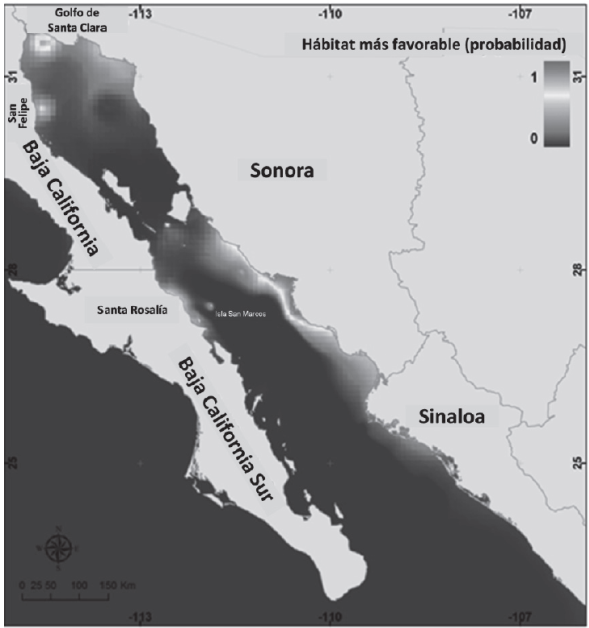
Discusión
Los estudios de distibución espacial de especies ha generado una gran cantidad de modelos de distribución de especies (Pont et al., 2015). Estos modelos dependen de las relaciones estimadas entre las condiciones ambientales presentes y presencia-ausencia de especies. Se han utizado con éxito para pronosticar los cambios de distribución de especies causadas por el calentamiento global (Ayala-Bocos et al., 2016; Perry et al., 2005). Se han convertido en la base de estudios de macroecología y biogeografía ya que son parte del estudio de nicho ecológico o condiciones habitables de la especie en el ambiente (Martínez-Meyer, Díaz-Porras, Peterson & Yáñez-Arenas, 2013; Schwartz, 2012), reconociendo esto es pertinente dejar en claro que la discusión sobre los diferentes modelos para el estudio de nicho ecológico no es la prioridad en el presente estudio, sino la aplicabilidad para pronosticar los cambios en la distribución de la especie bajo estudio ante efectos del cambio climático.
La modelación de la distribución de especies representa un área científica importante (Martínez-Meyer et al., 2013) que merece la atención de la comunidad académica ya que presenta algunos retos interesantes. Este trabajo muestra cómo usar MaxEnt para predecir la distribución de la almeja de sifón, MaxEnt solo requiere de muestras positivas y en el presente estudio mostró un buen desempeño. Los modelos generados por MaxEnt tienen una interpretación probabilística natural, dando una progresión suave de más a menos condiciones adecuadas. También se ha demostrado que los modelos pueden ser interpretados fácilmente, una propiedad de gran importancia práctica. Mientras MaxEnt aborda el problema de modelación de distribución de especies de manera eficaz, hay muchas otras técnicas que podrían ser utilizadas como campos aleatorios de Markov o modelos de mezcla. Es de destacar que MaxEnt (así como otros modelos de nicho ecológico) se han aplicado con mucho éxito en el medio terrestre (Andrén, 1994), pero en el medio acuático su uso ha sido más limitado. Esta limitación se ha adjudicado a la falta de información ambiental (Ibarra-Montoya, et al., 2012). Sin embargo, estos autores pudieron aplicar MaxEnt exitosamente (Area Under the Curve [AUC] = 0.90) para predecir el hábitat favorable de una cianobacteria en la presa Aguamilpa.
Para el caso del presente estudio los resultados del modelo MaxEnt muestran que la zona de San Felipe, en Baja California, no se encontró como la más adecuada para la distribución de almeja de sifón y es actualmente donde mayor captura se obtiene. En cambio en la region oriental del Golfo, particularmente la parte central, el hábitat es el mas adecuado, pero la explotación es aún incipiente. La explicación a esto es que la zona central del Golfo de California (que incluye Guaymas) aún se encuentra subexplotada. Las razones las explican Aragón-Noriega et al. (2012) y entre otras discuten razones de mercado. Por esa razón las capturas comerciales no se deben considerar como evidencia de mayor presencia por hábitat favorable para la especie. Un estudio de densidad para cada zona aclararía mejor si Guaymas es mejor como hábitat para la especie bajo estudio en comparación con la zona de San Felipe.
Los resultados del modelo sugieren que menos de la mitad del área total incluida en el estudio presenta sitios con calidad de hábitat óptimo para la presencia de la especie (Figura 3). Los sitios con mayor probabilidad de presencia de P. globosa se localizan en Guaymas (0.83) el Alto Golfo de California (0.69) y en la costa de Santa Rosalía e Isla San Marcos (0.70). Estos resultados sugieren que la distribución de P. globosa está restringida hacia el interior del Golfo de California, ya que a pesar de incluir registros del sur de Sinaloa en el análisis, la probabilidad de que esta especie se distribuya cerca de la boca del Golfo de California o habite en la región de Bahía Magdalena (siendo simpátrica con P. generosa en el oeste de Baja California) es extremadamente baja (0.005). Aquí es importante señalar que a pesar de que el análisis arrojó baja probabilidad de presencia de P. globosa en Bahía Magdalena, Suárez-Moo et al. (2013) demostraron que la especie que se captura en ese cuerpo de agua es P. globosa. Es importante destacar que la zona central del Golfo de California se ve favorecida por eventos de surgencia (Hamman, Nevárez-Martínez & Green-Ruiz, 1998) lo que la hacen una zona de alta productividad primaria y al ser P. globosa una especie filtradora la zona es un hábitat favorable.
Al pronosticar el cambio en la distribución de P. globosa bajo un escenario de elevación de temperatura oceánica del Golfo de California, por efecto del cambio climático, se observa que su distribución se restringe al norte del Golfo de California cerca de las costas de Guaymas, Golfo de Santa Clara, Sonora y San Felipe, Baja California. Estos resultados deben ser tomados con cautela ya que las demás variables incluidas en el modelo se mantuvieron constantes lo que puede provocar resultados incompletos. Es importante destacar que los efectos del cambio climático en especies sésiles como son las almejas que viven adheridas al fondo marino puede ser más abrupto que en especies de peces que pueden migrar hacia zonas donde el clima es más favorable. Perry et al. (2005) y Pont et al. (2015) señalan que hay suficiente evidencia para afirmar que la respuesta de las principales especies de peces ante el calentamiento global es migrar hacia los polos, hacia aguas más profundas o hacia arriba, en términos de elevación, donde encuentran condiciones térmicas más adecuadas a su hábitat preferido. Perry et al. (2005) hacen una advertencia acerca del impacto que está causando la elevación de la temperatura del mar en las pesquerías comerciales del mar del norte por los continuos cambios en la distribución y alteraciones en las interacciones de la comunidad de peces. Su estudio lo hicieron en 25 especies de peces tanto comerciales como no comerciales.
Los pescadores que basan su economía en la explotación de recursos como las almejas (que es el caso de la especie objeto de estudio en el presente trabajo) serán los más directamente afectados ante un escenario de elevación de la temperatura promedio del Golfo de California, derivada de efectos del calentamiento global, por las razones arriba mencionadas. Al ser la explotación de este recurso administrada por bancos y cuotas (Aragón-Noriega et al., 2012) otorgadas a solicitantes que viven en las zonas aledañas de la distribución conocida de la especie, el resultado de este estudio es de importancia para prevenir los posibles cambios en la distribución ante el escenario de cambio climático. También es motivo para recomendar estudios como el presente para otros recursos pesqueros del Golfo de California y poder prever los potenciales cambios en distribución del resto de recursos pesqueros.
Conclusión
Los resultados del presente estudio indican que es posible generar mapas de predicción de distribución potencial de la almeja de sifón a partir de registros de presencia (ya sea derivados de la pesca comercial o de estudios científicos) en combinación con variables ambientales. Esto tiene un enorme potencial para la biología de la conservación y para la administración de recursos pesqueros ya que se identifican áreas críticas de distribución de las especies de importancia comercial (como la del presente estudio P. globosa). El modelado de nicho ecológico ha demostrado ser una herramienta útil para predecir la distribución potencial de las especies en el contexto del cambio climático global. En este estudio se utilizó el modelo MaxEnt para pronosticar la distribución de la almeja de sifón del Golfo de California con la idea de proyectar su distribución potencial bajo los efectos del cambio climático que se espera ocurran en el año 2050. Si bien los estudios de nicho ecológico son útiles para elucidar los factores que influyen en la distribución de P. globlosa, sería interesante continuar con la investigación explorando también registros de captura y posibles sitios de sobreexplotación tomando en cuenta que, en un escenario de cambio climático, los sitios de captura pueden modificarse.

