











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Greenhouse gas emissions continue to increase on a worldwide scale and world’s forests have been well documented as providing an extensive carbon sink. Clearly, action must be taken to reduce CO2 emissions (De la Vega-Meneses & Rivero-Villar, 2014; Valdés-Ramírez, 2011). However, global estimates of nature of that carbon (C) sink may be biased if they use only limited data on carbon amount stored in different forests. While forests of Mexico may provide a massive carbon sink, assessing variability of that carbon sink across the various ecosystems of Mexico will help researcher refine accuracy of their models of carbon sequestration.
Knowing how C content in forest ecosystems has changed over recent years can provide important information on its climate response. Such information can be used to understand climate change behavior and help identify actions to mitigate it. This information can be obtained through dendrochronological study. This technique provides historical information on environmental conditions of a given place, and is used to study changes in ecology, climatology, environmental chemistry, and geomorphology (Giraldo-Jiménez, 2011).
Current methods for estimating C capture in forests regularly use destructive methods for chemical analysis, having a negative impact on their ecosystems. Hence, it has become important to develop a methodology that does not involve destruction of the tree, without compromising reliability of results. Several studies involving calculation of C in forests have been conducted (Herrero de Aza, Turrión, Pando & Bravo, 2011; Jones & O’Hara, 2012; Martin, Thomas & Zhao, 2013; Martin & Thomas, 2013). However, few studies use dendrochronology for this purpose (Lamlom & Savidge, 2006).
Background for this study is presented in Pimienta, Domínguez-Cabrera, Aguirre-Calderón, Hernández & Jiménez-Pérez (2007), where estimates of biomass (B) and C content for Pinus cooperi Blanco were carried out in forests in the municipality of Pueblo Nuevo, Durango. Martínez-Barrón (2014) models C dynamics in forests of this region.
A practical way to calculate C is by estimating B. Návar (2009) proposes an allometric equation for this purpose. This equation uses normal diameter (D) and average density (ρ) to obtain an accurate estimate of C content for an individual tree.
The value most widely used for C concentration in B of trees is 50% (Lamlom & Savidge, 2006; Yerena-Yamallel, Jiménez-Pérez, Aguirre-Calderón, Treviño-Garza & Alanís-Rodríguez, 2012). When using estimates of B in such studies, it is assumed that there is a uniform distribution among all trees, i.e., that ρ is constant. This does not take into account that it varies dependent upon weather conditions and age of tree (Babst et al., 2014). The main implication of accepting this premise is that results are not accurate because temporal variations in ρ reflect on B, and thus on the C of a tree. Because width of ring (RW) and its average ring density (ρ) varies according to spatial and environmental conditions, it is necessary to consider these two variables to obtain more accurate estimates of C flows in ecosystems.
Briffa et al. (2001) used data on ρ to make a reconstruction of temperature changes over the past 600 years for a region that accounts for nearly the entire northern hemisphere.
Babst et al. (2014) performed an evaluation of B changes above the ground at five sites, using RW and ρ. Subsequently, these estimates of B were validated, and compared with monthly flow data for CO2. However, to our knowledge, there are no such data for Mexican forests.
In our study, chronologies of C capture in Pinus cooperi from Durango, Mexico were obtained using a novel combination of dendrochronology to date growth rings and allometric relationships to estimate B, incorporating variations in ρ. Such data can easily be incorporated into a practical and reliable model to calculate variations in C content of a forest ecosystem over a short-term basis. We hypothesized that neglecting ρ miscalculate cumulative C capture.
Materials and Methods
A population of P. cooperi located at Las Rusias in the Sierra Madre Occidental in Durango, Mexico (Table 1) was selected for study. Deformed or damaged trees were not sampled, neither were those whose growth was influenced by competition for light or nutrients (Pompa-García, Miranda-Aragón & Aguirre-Salado, 2014).
Table 1 Characteristics of the sampling site at Las Rusias, Durango, Mexico.
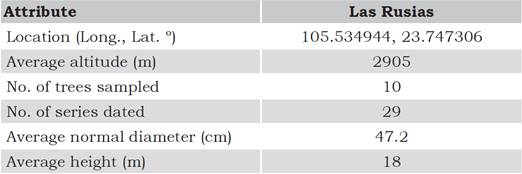
Source: González-Cásares, Pompa-García, Meléndez-Soto & Solís-Moreno. (2015).
At sampling site, of each individual, at least two radial wood cores were collected from a height of 1.3 m (above ground level) by a non-destructive method using increment borers (Ø = 5.1 mm). Each sample was processed (dried and polished), subsequently each year was recorded and each growth ring was measured (µm). Samples were sent to laboratory of wood anatomy and growth rings at Department of Forest Sciences at the University of São Paulo in Brazil. Using densitometric techniques, (ρ): (g/cm3) was calculated for each ring. Tree-ring width annual values (radial growth) were used to reconstruct historical tree diameters D (m), assuming circularity in growth rings. We used allometric equation for B estimation of northwestern Mexico forests (Návar, 2009) for P. cooperi (1). Thus, B was calculated for the aerial part, as well as thicker roots, as follows:
where yi = biomass in components of trees i; D = Normal diameter; ρ = Average ring density; a and b are statistical parameters. For total above the ground: a = 0.2018 and b = 2.2907; while for the thick roots: a = 0.0051 and b = 2.668.
Results
Our results showed variation in RW and ρ per year (Figure 1). Curves of C content showed uniform growth, related to their annual accumulation of C. There is an increase in total content of C in our results, where ρ is included, compared with data obtained from traditional equation, which assumes a uniform ρ (González-Cásares et al., 2015). This increase represents more than 50% of total C content, confirming that traditional methodologies (i.e., using a constant ρ) underestimate carbon sequestration, and give inaccurate estimates of C capture. We intend to carry out study at different sites to evaluate variation in ρ with respect to various environmental conditions.
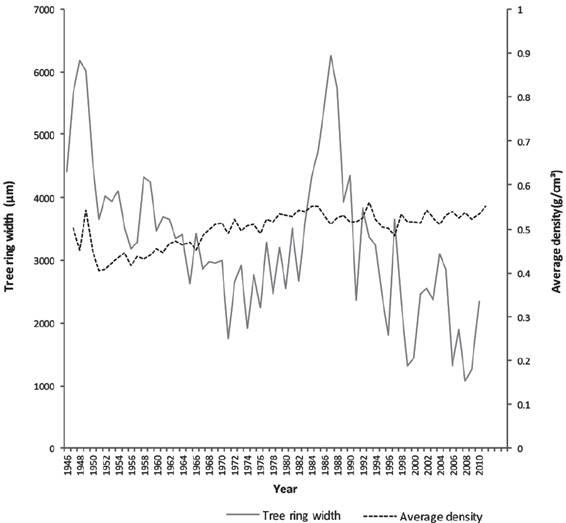
Source: González-Cásares et al. (2015).
Figure 1 Average ring density (ρ)estimated per year and ring width (μm) for 10 trees selected at the sampling site.
Figure 2 shows that during first years of growth, difference between the two estimates is not so large. However, from 1970 onwards, this difference increases markedly, resulting in a noticeable change at present time. We found that trees at this site accumulate more C with age. We also found a negative correlation between total width of the ring and contents of C, where as width decreases, C content increases.
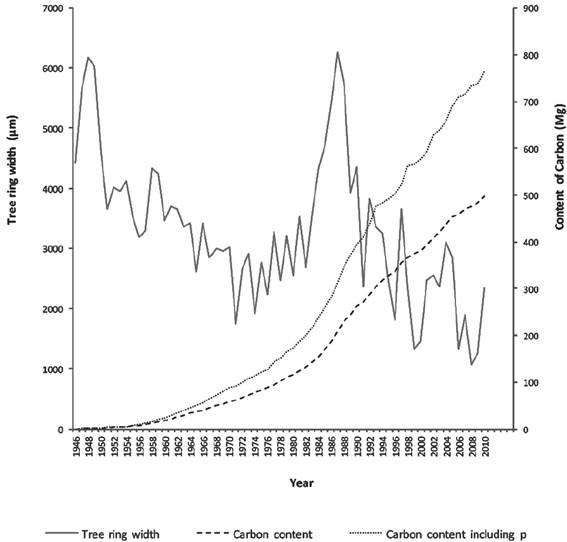
Source: González-Cásares et al. (2015).
Figure 2 Average cumulative carbon content (mg) estimated with and whithout ρ and ring width (μm) for 10 trees selected at the sampling site.
Discussion
Results obtained in the first study conducted at this site (González-Cásares et al., 2015) are markedly lower than those obtained in this study, taking into account ρ, resulting in an increase in C content cumulative average from 497.35 mg to 763.86 mg. However, in the early years of growth (1946-1960) some similarity was observed between two curves of C content. This similarity suggests consistency with annual biomass increases and tree age. Another aspect to be considered when calculating C content is wood formation processes. In this species, earlywood (EW) represents about 82% of the RW, while remaining 18% is latewood (LW), depending on the site (Pompa-García & Domínguez-Calleros, 2015). These differences in growth have implications for forest productivity, carbon cycles and other ecological processes faced by the species. This may be one of the most significant implications of taking into account ρ in this study. Even though this species has a higher percentage of EW compared with LW, it is important to note that LW tissues are more compact, resulting in a greater ρ. This explains what happens at trunk periphery, where despite the fact that RW decreases, C content increases because the LW volume is greater. Using values of ρ in allometric equation, allows us to assess variations through time, not related to growth processes (Babst et al., 2014). To date, few studies in this area have considered differences in EW and LW properties (Pompa-García & Domínguez-Calleros, 2015). We intend to expand our study to other sites to evaluate correlations between RW and ρ with respect to study site (Babst et al., 2014; Taki, Nobori & López-Cáceres, 2014) as well as environmental conditions. In particular, summer temperature has been shown to influence RW and ρ of wood (Fan, Bräuning, Yang & Cao, 2009; Grudd, 2008).
Conclusions
Our results suggest that estimating C, based on an allometric model sustained only by D of a tree results in inaccurate estimate of C content at a given site because variability in ρ that occurs in individual trees has not been taken into account. We clearly show that more accurate results are obtained by measuring and incorporating ρ into estimates of C based on B.

