











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El cultivo de Agave tequilana, así como el tequila, tienen alta importancia económica y social. El tequila ha sido la bebida más popular y tradicional de México. Sin embargo, en cuanto a investigación de las especies se le ha dado poca importancia en comparación con otros cultivos (Comité Regulador del Tequila [CRT] / Comité Técnico Agronómico [CTA], 2005; González, Del Real & Solís, 2007).
Se considera que la enfermedad tizón foliar causada por el hongo Cercospora agavicola, misma que se presenta de octubre a marzo, tiene requerimientos de alta humedad relativa y se caracteriza por provocar manchas ovaladas y secas de color gris en las hojas cercanas al cogollo, manifestando una clorosis generalizada y avanza hacia el centro del mismo hasta alcanzar la piña, impidiendo la recuperación de la planta. Al inicio las plantas afectadas aparecen en forma aislada, formando manchones conforme el contagio avanza. La enfermedad es muy agresiva dado que el síntoma es irreversible y si no se controla puede provocar la muerte de la planta en un lapso de 30 días, dependiendo de las condiciones ambientales.
Ayala, Yañez & Johannes (2005) mencionan que en enero de 2003 en el municipio de Pénjamo, Guanajuato, se observó una nueva enfermedad en A. tequilana. El material se llevó a Alemania para confirmar su identificación. Se completaron los postulados de Koch satisfactoriamente confirmando a Cercospora agavicola como el organismo causante de la mancha y necrosis de la hoja del agave. Cercospora agavicola se comparó con otras especies causantes de cercosporosis basada en la secuencia de la región de los genes ITS-rDNA y genes de la elongación del factor 1-a, actina, calmodulina e histona H3.
Groenewald, Groenewald & Corus (2005) argumentan que el género Cercospora es uno de los más diversos. Cercospora appi sensu lato es el nombre más antiguo del complejo morfológicamente indistinguible de especies de Cercospora que presenta en un amplio rango de hospederos. Se reportan más de 3000 especies de Cercospora, de las cuales actualmente se reconocen 659 y otros 281 nombres de otras especies morfológicamente idénticas. Generalmente, las especies de Cercospora son específicas a los hospedantes, al nivel de familia, género o planta, por lo que este concepto ha llevado a la descripción de un amplio número de especies.
Chen, Lyda & Halliwell (1979) reportan que Cercospora se ubica entre los patógenos de plantas más frecuentes y destructivos. Afecta un amplio rango de hospedantes, siendo uno de los factores de patogenicidad la producción de una exotoxina de color rojo denominada cercosporina.
Morfológicamente, los conidios de Cercospora son alargados de forma filamentosa y hialinas, se desarrollan sobre conidióforos agrupados que nacen de un punto común sobre el tejido vegetal; estos conidios son transportados por el viento hasta la presencia de una película de agua sobre el follaje, y los tubos germinativos penetran en la hoja a través de los estomas; una vez que el patógeno ingresa al tejido vegetal, produce toxinas, ocasionando la muerte de las células vegetales dando origen a las lesiones necróticas.
Vanegas, De León & Narro-León (2007), trabajando con líneas endogámicas de maíz, encontraron que la resistencia a Cercospora zea maydis se controla por efectos maternos o citoplásmicos. La acumulación de genes de resistencia y la conformación de variedades sintéticas resistentes es una alternativa a implementar en programas del mejoramiento de resistencia al tizón de maíces tropicales.
Con base en lo anterior, se planteó el objetivo de seleccionar genotipos resistentes evaluando la patogenicidad de Cercospora agavicola en hojas de vitroplántulas de A. tequilana Weber variedad azul irradiadas con Rayos Gamma Co60, así como seleccionar genotipos resistentes.
Materiales y Métodos
Colecta en campo de plantas enfermas
El material vegetal enfermo se colectó en plantaciones comerciales de agave de tres años de edad ubicadas en el municipio de Arandas, perteneciente a la zona de los Altos de Jalisco, México, ubicado en las coordenadas 20°41’58’’ latitud norte, 102°21’57’’ longitud oeste y una altitud de 2049 msnm.
El muestreo se llevó a cabo en tres predios, seleccionando 10 surcos aleatoriamente con una frecuencia cada 5 o 10 surcos, dependiendo de las dimensiones de cada plantación. Se hicieron recorridos observando las plantas del surco seleccionado y del surco adyacente, en los que se localizaron plantas con los síntomas del hongo. Se separó la hoja de la planta, se cortó la sección donde se presentaba la presencia del hongo, se envolvió en papel periódico humedecido y se colocaron en bolsas de plástico, mismas que se identificaron con el predio, la fecha de la colecta y el número secuencial de acuerdo con las hojas que se obtuvieron.
Aislamiento del hongo (Cercospora agavicola)
El medio de cultivo utilizado para el aislamiento, purificación y multiplicación del hongo C. agavicola fue papa dextrosa agar y agave (PDAA) (Espitia, Limón & Calleros, 2012). El medio de cultivo se preparó moliendo 100 g de hojas de agave de la parte central de la planta (hojas jóvenes) en un litro de agua. Se pesaron 250 g de papa, se cortaron en trozos y se pusieron en un matraz Erlenmeyer que contenía un litro de agua destilada. Se dejó hervir durante 20 min y se filtró con algodón. Se midieron 500 ml tanto del filtrado procedente del molido de las hojas de agave, como del concentrado de la cocción de la papa.
Posteriormente, se adicionaron 20 g de agar bacteriológico y 10 g de dextrosa anhidra. Se ajustó el pH a 6.0 con NaOH y HCl 1N. Se esterilizó en autoclave a 15 lb de presión y 121 ºC durante 15 min. El medio estéril se dejó enfriar hasta una temperatura aproximada de 40 ºC y se adicionaron 2 g/L de AGRYGENT (Gentamicina).
Se vaciaron de 15 mL a 20 mL de agar en cajas de Petri estériles y se dejaron solidificar. Las cajas de medio preparadas fueron guardadas en refrigeración para su uso posterior utilización.
Las hojas de agave con los síntomas del hongo se mantuvieron cubiertas con papel sanitario en bolsas de plástico previamente humedecidas a 25 ºC y 80% de humedad relativa para favorecer la germinación de las esporas.
Las hojas se seccionaron dejando únicamente las áreas que presentaban las lesiones del hongo (de 3 cm a 5 cm), mismas que se colocaron en cajas Petri en la cámara de flujo laminar en condiciones asépticas.
La inoculación se llevó a cabo al realizar una herida con aguja sobre la superficie de las hojas con los síntomas del hongo. Además, se colocaron esporas en tubos con 1 mL de agua destilada esterilizada para distribuir uniformemente el agua con la concentración de esporas en toda la caja Petri con el medio de cultivo PDAA. Las cajas Petri se sellaron, rotularon y se pasaron a la cámara de incubación a una temperatura de 25 ºC en oscuridad.
Purificación del hongo
Los materiales inoculados se revisaron cada cinco días para dar seguimiento al desarrollo del hongo. Se seleccionaron las muestras que presentaron los síntomas que induce C. agavicola. Se tomó una muestra y se colocó en un portaobjetos y se observaron al microscopio. Los aislamientos que presentaron las características morfológicas de Cercospora se re-aislaron y se sembraron nuevamente en medio fresco PDAA para su multiplicación.
Periódicamente se llevaron a cabo las revisiones de las inoculaciones, de las que se tomaron muestras para la observación en microscopio y el reconocimiento de las esporas, así como la presencia de la toxina “cercosporina”. El proceso se continuó hasta que se desarrollaran todos los síntomas sin que estuvieran contaminadas, ya fuera por otros hongos o por bacterias.
Además de la identificación de estructuras morfológicas del hongo, se corroboró la presencia de toxina de Cercospora por la presencia de un pigmento rojo, característico del hongo y que corresponde a producción de la toxina “cercosporina”. Una vez que se alcanzó la purificación del hongo, se cambió de medio para su multiplicación cada 21 días, corroborando la morfología de los conidios, así como la presencia de la toxina “cercosporina”.
Radiación
Previo a la inoculación, las plántulas se irradiaron en el Instituto Nacional de Investigaciones Nucleares (ININ), utilizando un irradiador Gammacell Modelo GO-220® (Ontario, Canadá) para aplicar siete dosis de irradiación: 0 (testigo), 5 Gy, 10 Gy, 15 Gy, 20 Gy, 25 Gy y 30 Gy con rayos gamma Co60.
Pruebas de patogenicidad
Las pruebas de patogenicidad se llevaron a cabo en hojas de plántulas de agave azul tequilero obtenidas in vitro a través de yemas axilares (Ángeles-Espino et al., 2012) mediante el método de hoja desprendida (Pettitt, Wainwright, Wakehamb & White, 2011).
Obtención del inóculo
Se preparó una suspensión de agua bidestilada esterilizada conteniendo las esporas del hongo. En la cámara de flujo laminar con una aguja de disección se obtuvieron las esporas del hongo y se colocaron en tubos con 1 mL de agua bidestilada. Se tomó una gota agua y se colocó en la cámara Newbauer para determinar el número de esporas en la suspensión.
Conteo de esporas
Se colocó la muestra bajo el cubreobjetos con la suspensión de esporas alcanzando una altura de 0.1 mm. Se contaron las esporas que estuvieron dentro de los cuadros grandes (o sea, en 25 cuadrados medianos), por lo que la concentración en cada muestra se calculó con la siguiente fórmula:
Tomando en cuenta que las esporas se colocaron en una dilución de 1 mL, las esporas en la concentración quedaron:
La inoculación en el tejido se llevó a cabo con una concentración de 20 000/mL-1, por lo que el volumen a aplicar se calculó de la siguiente manera:
Volumen = concentración de esporas a inocular /número de esporas contadas.
Diseño experimental
Se diseñó un experimento factorial con arreglo combinatorio y una distribución completamente a azar con tres repeticiones, donde el factor “A” fueron las dosis de radiación (0 Gy, 5 Gy, 10 Gy, 15 Gy, 20 Gy, 25 Gy y 30 Gy) y el factor “B” las plantas por tratamiento (Reyes, 1978). Los datos se procesaron por análisis de varianza y las medias se compararon mediante la prueba de rangos múltiples de Tukey al 5% de probabilidad. Además, se realizó un análisis de regresión lineal entre las dosis de radiación y el diámetro de las lesiones. Las mediciones del diámetro de las lesiones se hicieron a los 21 días posteriores a la inoculación.
Inoculación
La inoculación se llevó a cabo en cajas Petri, a las que se les colocó papel secante, se envolvieron en papel aluminio y se esterilizaron en autoclave a una presión de 1.4 kg/cm2 a una temperatura de 121 ºC durante 15 min. Además, se esterilizó agua destilada para la dilución de las esporas. La inoculación se llevó a cabo en cinco plántulas (genotipos) por dosis de radiación, usando la técnica de hoja desprendida, colocando tres foliolos por cada genotipo. En la cámara de flujo laminar bajo condiciones de asepsia se llevaron los contenedores con las plántulas que se encontraban en ambiente estéril. Se extrajeron las plántulas y se colocaron en cajas Petri previamente esterilizadas, donde se retiraron las hojas, mismas que se fueron colocando en nuevas cajas Petri con el papel secante y esterilizadas. Una vez que se retiraron las hojas, las plántulas se regresaron a sus contenedores y se mantuvieron bajo condiciones de esterilidad. El conteo de las esporas se llevó a cabo en la cámara New Bauer y se hicieron las diluciones a una concentración de 20 000 esporas/mL-1. Con una jeringa estéril, se añadieron 10 mL de agua previamente esterilizada en cada caja Petri, humedeciéndolo a saturación, pero sin que sobrepasara el grosor del papel secante. A cada hoja se le hicieron tres heridas de manera longitudinal al tamaño de esta y se añadieron 0.5 mL en cada hoja, procurando que este quedara uniformemente distribuido a lo largo de las lesiones que se hicieron. Se sellaron, rotularon y se colocaron en un estante, verificando que no se hubiera derramado de las hojas el agua con las esporas inoculadas.
El testigo estuvo conformado por dos tratamientos en los que las vitroplántulas no fueron irradiadas. A las hojas del primero se le hicieron las heridas y se inoculó con agua bidestilada estéril, en las hojas del segundo se le hicieron las heridas y fueron inoculadas con las esporas del hongo.
A los cinco días posteriores a la inoculación se revisaron las muestras, asegurándose que el agua con las esporas se encontrara en cada hoja, así como la humedad del papel secante. En las hojas que fue necesario se reinoculó manteniendo la concentración a 20 000 esporas/mL-1 sellando nuevamente las cajas. Las revisiones se hicieron cada 5 días, reponiendo la humedad cuando así se requirió.
A los 10 días posteriores a la inoculación se hizo la primera observación para detectar la presencia del hongo, contabilizando el número de hojas por tratamiento que presentaron algún indicio de lesión, obteniendo el porcentaje por tratamiento. Las mediciones se llevaron a cabo a los 21 días posteriores a la inoculación, tomando hojas procedentes de vitroplántulas, estableciendo un área de 2 cm2 tomando 2 cm de manera longitudinal y 1 cm a lo ancho de las hojas. A partir de esta superficie se hicieron las mediciones de las lesiones. Además, se obtuvieron los valores porcentuales del área afectada en cada tratamiento con respecto al tamaño promedio de la lesión del tratamiento sin irradiar, el que se consideró como el diámetro máximo que alcanzaron las lesiones (100%).
Resultados y Discusión
Aislamiento y purificación de Cercospora agavicola
Se obtuvo el aislamiento y la purificación del hongo colectado en campo. El proceso llevó 18 meses hasta obtener la cepa de C. agavicola. En cada siembra que se realizó del hongo en medio fresco se hicieron las observaciones en microscopio óptico para verificar que los conidios en el medio de cultivo correspondían a C. agavicola.
Una vez que se tuvieron los cultivos de C. agavicola sin la presencia de contaminantes, se observó la presencia de la toxina cercosporina que produce un color rojo oscuro intenso en el medio PDA (Amuaykanjanisn & Daub, 2009; Lapaire & Dunkle, 2009), lo que confirmó que el hongo cultivado correspondió a C. agavicola (Figura 1).

Pruebas de patogenicidad
En la primera evaluación se hizo el conteo de las hojas que presentaron indicios de lesión a los 10 días posteriores a la inoculación. El testigo absoluto que fue inoculado con agua destilada no se observó la presencia C. agavicola. En los tratamientos inoculados con el hongo, el testigo (0 Gy) y las dosis de 5 Gy y 10 Gy, el 21% de los foliolos tuvieron lesión, en las dosis de 15 Gy y 20 Gy el 10% de los foliolos con lesión y de 7% y 5% en las dosis de 25 Gy y 30 Gy, respectivamente (Tabla 1). A pesar que en las dosis de mayor radiación el porcentaje de hojas con lesión fue menor, los síntomas se presentaron en todos los tratamientos evaluados, lo que coincide con Pettit et al. (2011), quienes reportan la presencia de necrosis al inocular hojas de crisantemo (Dendrathema grandiflorum Ramat) separadas de la planta con Phytium sp.
Tabla 1 Infección de Cercospora agavicola en foliolos de vitroplántulas mutantes de Agave azul tequilero.
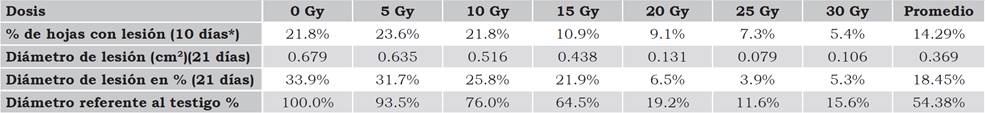
*días posteriores a la inoculación.
Fuente: Elaboración propia.
Por otra parte, al evaluar el diámetro de las lesiones a los 21 días, estas disminuyeron conforme la dosis aumentó y los valores porcentuales indican que en el testigo y la dosis de 5 Gy, las lesiones abarcaron el 32% del área del foliolo (2 cm2), disminuyendo a valores de 20% en las dosis de 10 Gy y 15 Gy; mientras que en dosis de 20 Gy y superiores, el área afectada correspondió al 5% de la hoja. La diversidad en el tamaño de las lesiones demuestra que la radiación ionizante absorbida por el tejido generó mutaciones en las células que inciden directa o indirectamente con los mecanismos de defensa, que son inducidos como consecuencia del reconocimiento, actuando en conjunto para detener el avance del patógeno (Madriz, 2002), ya que el comportamiento de las plántulas frente al ataque del hongo presentó una respuesta diferenciada en cada dosis de radiación que recibieron las plántulas (Figura 2).
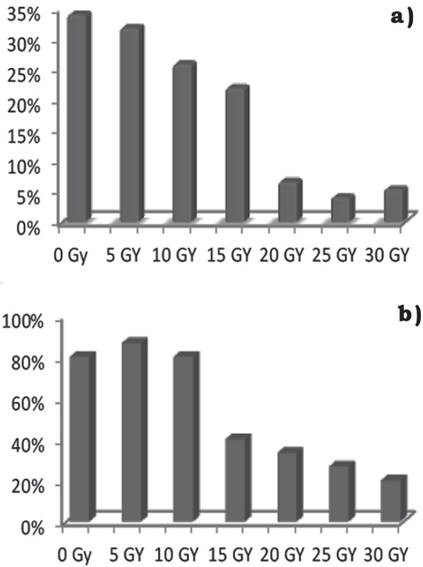
Fuente: Elaboración propia.
Figura 2 Porcentaje de foliolos infectados por C. agavicola 10 días posteriores a la inoculación: a) porcentaje en relación con las dosis de radiación. b) porcentaje dentro de cada dosis de radiación.
Por otra parte, al evaluar el porcentaje de las lesiones con respecto al testigo, se tuvieron tres grupos: el primero conformado por las dosis de 0 Gy y 5 Gy, cuya lesión cubrió más del 90%; el segundo con el 75% y 65% de la lesión que correspondió a las dosis de 10 Gy y 15 Gy; y el tercero donde la lesión abarcó del 10% al 20% del foliolo y que correspondió a las dosis de 20 Gy, 25 Gy y 30 Gy (Tabla 1). Con base en los resultados, se determinó la tendencia que indica que las lesiones con menor diámetro se presentaron en los foliolos que fueron irradiados con dosis iguales o superiores a 20 Gy, donde la inducción de mutaciones es mayor. Resultados similares fueron obtenidos por Gosal et al. (1998) al irradiar yemas de papa cultivadas in vitro a 20 Gy y 40 Gy con rayos Gamma Co60. Los micro-tubérculos se multiplicaron in vitro y se plantaron en macetas. Las plantas obtenidas se seleccionaron para tizón tardío, encontrando que las plantas irradiadas con 40 Gy mostraron resistencia, mientras que esta fue menor en los tratamientos con 20 Gy. La progenie resistente se plantó en campo y se inoculó con esporas del hongo. En campo, aproximadamente el 56% de la progenie segregante mostró resistencia, mientras que para la siguiente propagación el patrón de resistencia se incrementó al 72% en las plantas multiplicadas in vitro M1V3.
Es importante mencionar que el planteamiento de las dosis de radiación se consideraron pretendiendo conocer la respuesta de las vitroplántulas frente a la infección ocasionada por el hongo C. agavicola, y así establecer una tendencia que muestre la mejor respuesta en los tratamientos frente al ataque de acuerdo con el tamaño de la lesión que se origina en cada una de las dosis de radiación, así como el comportamiento de los tratamientos cercanos a la dosis letal media (DL50). Correa-Victoria (2001) menciona que la pérdida de resistencia a Pyricularia grisea Sacc. en arroz se debe a que se busca una resistencia vertical con genes mayores, generando resistencia en el patógeno. Con la idea de comprobar la resistencia poligénica, se obtuvieron 37 mutantes con rayos gamma de una línea comercial y fueron inoculadas con el hongo. Los resultados mostraron que se requiere la evaluación de poblaciones grandes de mutantes para incrementar la probabilidad de identificar una resistencia genética a los genotipos evaluados como una fuente de resistencia complementaria frente al patógeno.
De modo paralelo, los resultados de esta investigación indican que se indujeron mutaciones que modificaron diversos procesos dentro de los mecanismos de defensa químicos y bioquímicos que desarrollan las plantas frente a la presencia de un patógeno. Es importante notar que la inoculación se hizo en tejido joven (vitroplántulas), por lo que no estaba desarrollado, además que no alcanzó la suberización y lignificación que caracteriza a la familia de las agaváceas, además que se encuentran en un ambiente estéril y que no están adaptadas a las condiciones del medio ambiente.
Como se muestra en la Tabla 2, se obtuvo una diferencia altamente significativa entre los tratamientos de irradiación y en las dosis aplicadas, mientras que entre las plantas que recibieron la misma dosis radiación, la diferencia no fue estadísticamente significativa al igual que en la interacción dosis por planta. En la prueba de Tukey se identificaron tres grupos en cuanto al diámetro de la lesión (Tabla 1): el primero con la dosis de 5 Gy que estadísticamente fue igual al testigo, con el 31% y 33%, respectivamente; el segundo lo conformaron las dosis de 10 Gy y 15 Gy, cuyas lesiones abarcaron el 26% y 21% del área foliar, respectivamente; y el tercer grupo corresponde a las dosis de 20 Gy en adelante, donde las lesiones cubrieron menos del 10% del área foliar. Por otro lado, el que no se haya presentado diferencia entre plantas que recibieron la misma dosis de radiación muestra que los cambios que se generaron en las células tuvieron el mismo comportamiento frente al ataque del hongo, debido a que el diámetro de las lesiones fue similar en los cinco foliolos evaluados en cada tratamiento. No hubo interacción debido a que el comportamiento de los tratamientos fue lineal (Y = -0.23 X + 0.713) con un coeficiente de determinación que indica que el 91% de los cambios en la disminución del diámetro de la lesión, se debieron al incremento en la dosis de radiación.
Tabla 2 Análisis de varianza y prueba de Tukey para diámetro de lesión causada por Cercospora agavicola en foliolos de agave azul tequilero irradiados con rayos gamma Co60.
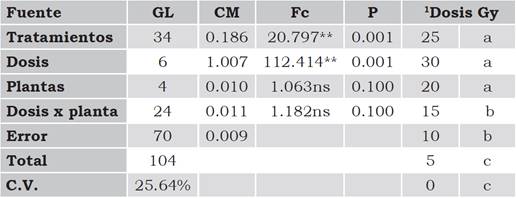
**Diferencia significativa (p < 1%); ns = diferencia no significativa (p > 5%)
1Tukey (5%). Tratamientos con igual letra no difieren estadísticamente (p > 5%)
Fuente: Elaboración propia.
La variación que se presentó en el tamaño de la lesión de acuerdo con la dosis de radiación absorbida por el tejido alteró los mecanismos de defensa de la planta, ya que las lesiones de C. agavicola se fueron reduciendo conforme la dosis se incrementó (Figura 3), por lo que una posibilidad es que las mutaciones que se produjeron hayan generado cambios en los genes que están relacionados de una manera directa o indirecta con la síntesis de metabolitos secundarios que forman parte de los mecanismos de defensa de la planta frente a un patógeno, tales como fitoalexinas, terpenoides, alcaloides, amidas, flavonoides, fenoles, compuestos proteicos antifúngicos o inhibitinas (Montes-Belmont, 2009).

Fuente: Elaboración propia.
Figura 3 Inoculación de Cercospora agavicola en vitroplántulas irradiadas con rayos gamma Co60. a) Testigo absoluto inoculado con agua destilada; b) 0 Gy inoculado con Cercospora agavicola; c) 5 Gy; d) 10 Gy; e) 15 Gy; f) 20 Gy; g) 25 Gy y h) 30 Gy.
La respuesta de la inoculación a las plántulas de agave concuerda con lo obtenido por Valdez & Orellana (2004), que regeneraron plántulas de caña de azúcar (Sacharum sp.) de la variedad SP 70-1284 obtenidas in vitro a partir de callos irradiados con 30 Gy, y se inocularon con la roya de la caña de azúcar Puccinia melanocephala. Se observó un 14.28% de los cambios fenotípicos totales en plántulas M1V1, se seleccionaron tres posibles mutantes con menor afectación frente a la enfermedad, por lo que el carácter de resistencia y la eficiencia del cultivo de tejidos y la inducción de mutaciones con la dosis de 30 Gy fue de un mutante por cada 125 plantas evaluadas en fase temprana de selección.
Este comportamiento indica que el diámetro de la lesión disminuyó conforme se incrementó la dosis de radiación, por lo que los resultados sugieren que puede estar presente un comportamiento genético aditivo, ya que la reducción no presentó valores contrastantes. Además, la diferencia en porcentaje entre el testigo y el tratamiento de 25 Gy fue de 30%, presentándose valores intermedios en las demás dosis.
La herencia de la resistencia poligénica es más compleja, debido a que los genes que intervienen son difíciles de identificarlos individualmente, conociéndose el efecto combinado de dichos genes como un todo que está asociada con genes menores de efectos aditivos que interaccionan con el ambiente (Russell, 1978).
Se presentaron cinco genotipos en los que no hubo presencia de lesión en la hoja, dos en la dosis de 20 Gy, dos en 25 Gy y una en la dosis de 30 Gy, que es una de las razones por las que los valores en porcentaje de las lesiones con respecto al testigo estuvieran por debajo del 10% con respecto al área infectada. Sin embargo, las lesiones en estas dosis son significativamente menores a los tratamientos con menor dosis de radiación, así como con el tratamiento sin irradiar.
Un aspecto a considerar es que de los porcentajes de infección, si bien la disminución en el tamaño de la lesión es progresiva con el incremento de la dosis de radiación, la diferencia en porcentaje entre el primer grupo (0 Gy y 5 Gy) con el segundo grupo (10 Gy y 15 Gy), oscila entre 5% y 10%; mientras que entre el primer grupo y el tercero (20 Gy, 25 Gy y 30 Gy) es de 30% en promedio, lo que indica que hay un efecto acumulativo y que estas dosis de radiación corresponden al valor de la dosis letal media DL-50.
García et al. (2000) estudiaron la variabilidad producida por las radiaciones Gamma para determinar la dosis óptima para los programas de mejoramiento de Mussa sp., cv. Gran Enano. Irradiaron brotes con dosis de 5 Gy a 40 Gy con Co60. Encontraron un aumento en la mortalidad a medida que aumentó la radiación, encontrando la LD50 entre 20 Gy y 25 Gy con cambios en la altura de las vitroplántulas, la morfología y pigmentación, así como un bajo porcentaje de plántulas con un comportamiento superior frente a la Sigatoka negra en comparación con el testigo. El efecto de la radiación obtenido en el cv. Gran Enano fue similar al que se presentó en las vitroplántulas de agave, ya que también la DL50 se encontró entre los 20 Gy y 25 Gy, así como el comportamiento frente a la presencia del fitopatógeno.
Morela, González, Castro & Castroni (2002) indujeron variabilidad genética en caña de azúcar (Sacharum sp.) al inducir mutaciones con rayos gamma en callos cultivados in vitro para obtener genotipos resistentes al Virus Mosaico de la Caña de Azúcar (SCMV, por sus siglas en inglés) a partir de variedades (B6749, B7987 y PR62258) susceptibles. Se irradiaron con Co60 (0.2 Kr, 4.8 Kr y 12 Kr). Se regeneraron 3502 plantas, mismas que se inocularon cinco veces con la raza B del SCMV, preparado de hojas de la variedad C571, altamente susceptible. De las plantas regeneradas, 82% permaneció libre de síntomas. En campo de 1607 plantas, 911 permanecieron sin síntomas, así de un 82% de las plantas que sobrevivieron a la inoculación artificial el 46% permaneció sin síntomas. Los resultados anteriores muestran que hay una tendencia a que la resistencia que presentan los genotipos al ser inoculados en plántula al evaluarse en campo no manifiesten los síntomas.
Conclusiones
Las pruebas de patogenicidad realizadas en las vitroplántulas irradiadas mediante hoja desprendida mostraron que hay cambios significativos en el diámetro de la lesión como una respuesta de los mecanismos de defensa de la planta, lo que indica un comportamiento genético aditivo (resistencia horizontal). En la etapa de plántula no permite identificar genotipos resistentes, por lo que la selección de mutantes (DL50 = 25 Gy) debe hacerse una vez que las hojas se encuentren en maceta y adaptadas (seis meses) con las hojas lignificadas y endurecidas, que favorezcan el establecimiento del patógeno en condiciones naturales, y así valorar el grado de resistencia para hacer la selección.

