











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Dentro de las oleaginosas, el cultivo del cacahuate (Arachis hypogaea) o maní, que forma parte de las semillas denominadas nuts a nivel internacional, tiene un papel preponderante dentro de la producción agrícola, no sólo por la producción de semilla para consumo directo, sino por los derivados que se obtienen, como el aceite, elaboración de pastas que se utilizan como materia prima en la industria tanto de alimentos como la química por la diversidad de compuestos orgánicos y metabolitos secundarios que tienen alta demanda en la farmacología (Ortega & Ochoa, 2003; Robles, 1985).
En México, el área que se cultiva con cacahuate ha fluctuado de 55 000 a 75 000 ha anuales, correspondiendo la mayor superficie sembrada a condiciones de temporal. La producción nacional actual es de 56 000 t, cuyo rendimiento medio es de 1300 kg/ha. Los principales estados productores mexicanos son: Puebla Oaxaca, Jalisco, Chihuahua, Sinaloa, Veracruz, Guanajuato, Morelos y Nayarit (Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias [INIFAP], 2002).
A pesar de que se multiplica vía sexual a través de semilla, botánicamente es una planta autógama que tiende a la endogamia, por lo que la variabilidad genética es reducida. La explotación de fuentes genéticas a partir de especies silvestres es extremadamente difícil porque hay diferencias en la ploidía entre especies cultivadas tetraploides y las silvestres diploides, junto con las barreras de compatibilidad (Channayya, Nadaf, Ganapathi & Praveenkumar, 2010). Aunque las técnicas convencionales han contribuido en gran medida al desarrollo de tipos deseables, la mutagénesis inducida es una herramienta potencial que permite inducir variabilidad genética, la cual podría ser valiosa para la selección de tipos deseables (Brock, 1977).
La mutación se define como el cambio en el material genético, así como el proceso por el cual sucede dicho cambio, ya que un organismo que presenta un nuevo fenotipo es el resultado de una mutación. El mismo autor consigna que el término mutación se refiere a cualquier cambio repentino y hereditario en el que el genotipo de un organismo no puede explicarse por la recombinación de la variabilidad genética preexistente (Gardner, 2007).
Las mutaciones son cambios que se presentan en la secuencia del ácido desoxirribonucleico (ADN) y que llegan a ser evidentes cuando se manifiestan en el fenotipo. La inducción de mutantes a través de radiación ionizante es una alternativa para, por una parte, inducir variabilidad y, por otra, propiciar la uniformidad en la maduración de las vainas, así como mejorar los índices de cosecha (Robles, 1986). En la inducción de mutantes, los cambios negativos se dan cuando se alcanzan altas dosis de radiación, ocasionando daños cromosómicos, por lo que el conocer la dosis letal media (DL50) incrementa la probabilidad de obtener mutaciones favorables en los programas de mejoramiento genético (Ahloowalia & Maluszynski, 2001).
El objetivo de la presente investigación fue inducir variabilidad genética con rayos gamma Co60 en semilla de cacahuate criollo tipo Virginia y selección de quimeras en la generación M1.
Materiales y métodos
Material biológico
Se utilizaron semillas de cacahuate (Arachis hypogaea) criollo de la región de Iguala, Guerrero, tipo Virginia, con un contenido de humedad de 12%, mismas que se sometieron a radiación ionizante con rayos gamma Co60. Los tratamientos se aplicaron en el Instituto Nacional de Investigaciones Nucleares (ININ), utilizando un irradiador Gammacell Modelo GO-220® (Ontario, Canadá) para aplicar siete dosis de irradiación: 0 (testigo), 50 Gray, 100 Gray, 150 Gray, 200 Gray, 250 Gray y 300 Gray (Gy), obteniendo la primera generación mutante (M1).
El experimento se llevó a cabo en el Campo Agrícola Experimental del Instituto Tecnológico y de Estudios Superiores de Monterrey (ITESM), Campus Querétaro, localizado a 20° 30’ 0’’ latitud Norte; 100° 12’ 35’’ longitud Oeste y una altitud de 1910 msnm, una precipitación pluvial de 630 mm anuales y una temperatura media anual de 17 °C. La siembra se realizó en un terreno de 1500 m2 preparado con un barbecho, dos pasos de rastra cruzada, una separación entre surcos de 0.75 m y de 0.30 m entre plantas. Se aplicó el riego de siembra y dos riegos de auxilio durante el desarrollo del cultivo, además de dos escardas y tres deshierbes manuales. No se hicieron aplicaciones de plaguicidas. La unidad experimental constó de diez surcos de 5 m de longitud espaciados a 0.75 m y una distancia entre plantas de 0.3 m.
Diseño experimental y análisis estadístico
Se estableció un diseño en bloques al azar con siete tratamientos y cuatro repeticiones (Reyes, 1978). Se obtuvo el análisis de varianza (p ≤ 0.01), y los promedios se compararon mediante diferencia mínima significativa (DMS) a una probabilidad del 5% (p ≤ 0.05). Se hicieron los análisis de correlación y regresión para determinar el efecto ocasionado por las dosis de radiación en los caracteres evaluados a través de estimar la dosis letal media o dosis reductiva media (GR50) y la dosis reductiva (GR30) como fuente de inducción de mutantes favorables (Robles, 1970).
Variables evaluadas
Altura de planta. En cada unidad experimental se tomaron tres plantas al azar a las que se les midió (m) el porte tomando de la base del tallo al ápice. La medición se llevó a cabo posterior a la floración cuando la planta alcanzó su máximo desarrollo una vez que se inició el elongamiento de los ginóforos, obteniendo el promedio por repetición.
Ginóforos por planta. Se tomaron tres plantas al azar de cada unidad experimental y se contabilizaron los ginóforos, resultando el promedio por repetición.
Longitud de ginóforo. Se midieron del nudo de formación hasta el nudo de unión con la vaina previo a su introducción en el suelo. Se tomaron tres ginóforos por planta y tres plantas por tratamiento de radiación, obteniendo el promedio de nueve plantas por repetición.
Porciento de floración. A los 50 días posteriores a la siembra se evaluó el porcentaje de floración por tratamiento de radiación en las cuatro repeticiones, contabilizando las plantas que presentaron flores en relación con el total de las plantas en cada una de las unidades experimentales.
Selección de quimeras. Se seleccionaron plantas que presentaron diferencias fenotípicas con respecto al testigo (0 Gy) en los caracteres de altura de planta, longitud de ginóforo y forma de la vaina. A la cosecha de cada planta se seleccionaron dos vainas basales, dos intermedias y dos apicales, con el objeto de evaluaciones en ciclos posteriores e incrementar la probabilidad de identificar mutaciones favorables en los genotipos seleccionados con base en los caracteres previamente descritos.
Resultados y discusión
Altura de planta
La tabla 1 muestra los resultados estadísticos de las variables que se evaluaron, mismas que se consideraron para la selección de mutantes que se llevó a cabo en el germoplasma irradiado. La altura de planta mostró diferencia significativa al 1% de probabilidad entre las dosis de radiación evaluadas con un coeficiente de variación de 13%. Mediante la prueba de comparación de medias (DMS), la altura se clasificó en tres grupos: el primero cuyo porte fue superior a los 30 cm que correspondió a las dosis de 0 Gy, 50 Gy, 100 Gy y 150 Gy; seguido de porte medio en las dosis de 200 Gy y 250 Gy y el porte menor en 300 Gy, como se muestra en la tabla 2 y la figura 1. Estos resultados concuerdan con los obtenidos por Lemus, Méndez, Cedeño & Otahola-Gómez (2002), quienes evaluaron el efecto de la radiación en la longitud de las plántulas de dos genotipos de Vigna unguiculata (L.) Walp., y encontraron diferencias significativas entre dosis absorbidas y entre genotipos, así como en la interacción. Los autores reportaron tres tipos de respuesta: 1) plántulas altas que correspondieron a las no irradiadas e irradiadas con dosis de 150 Gy; 2) plántulas de porte intermedio irradiadas con dosis de 300 Gy a 700 Gy para uno de los genotipos; y 3) plantas pequeñas con dosis de 300 Gy para el otro genotipo.
Tabla 1 Análisis de varianza en altura de planta, número y longitud de ginóforos y porciento de floración en plantas de A. hypogaea irradiadas con Co60.
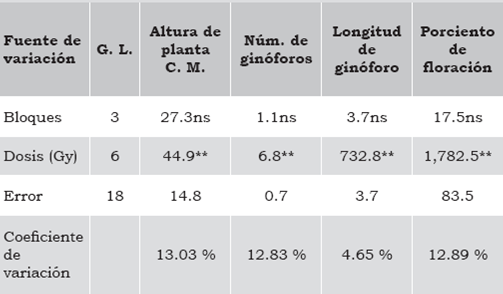
** = diferencia altamente significativa (p < 1%), ns = diferencia no significativa (p < 5%). Fuente: Elaboración propia.
Tabla 2 Efecto de la radiación con Co60 en altura de planta, número y longitud de ginóforos y porciento de floración en plantas de A. hypogaea.
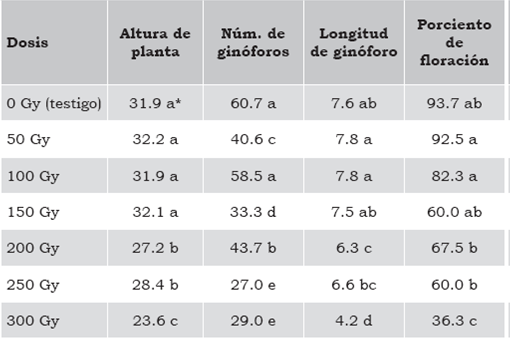
* Tratamientos con la misma letra no difieren estadísticamente (DMS 0.05). Fuente: Elaboración propia.
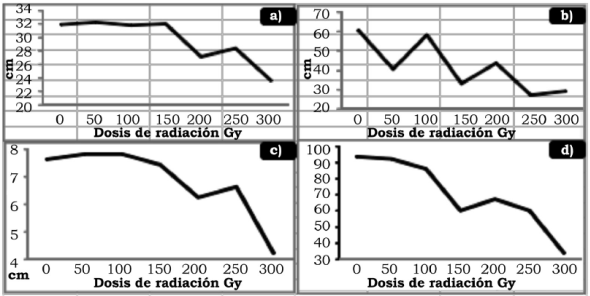
Fuente: Elaboración propia.
Figura 1 Efecto de la radiación gamma Co60 en la generación M1 en cacahuate (A. hypogaea). a) Altura de planta; b) Número de ginóforos; c) Longitud de ginóforo. d) Porciento de floración.
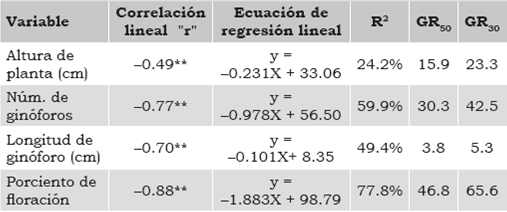
Por otra parte, DL50 o GR50 se presenta cuando un carácter manifiesta una disminución de 50% en su expresión con respecto al tratamiento testigo, debido a que la radicación absorbida provoca cambios en el ADN y origina mutaciones, mismas que causan alteraciones en la fisiología de la planta (Robles, 1970). Se obtuvo una correlación significativa al 1% de probabilidad (tabla 3), presentando una disminución en la altura conforme la dosis de radiación se incrementó; sin que esta se hubiese reducido en un 50%, lo que indica que la GR50 se presenta con dosis mayores a las evaluadas en la presente investigación. De acuerdo con la regresión, se requiere incrementar la radiación a 700 Gy para que surja la dosis letal media; sin embargo, la GR30 se presentó en la dosis de 300 Gy (23.5 cm); además, el coeficiente de determinación (R2) indica que solo el 24% de los cambios en la altura de planta es atribuible a la radiación absorbida, lo que explica, por una parte, el que no se haya presentado la GR50 y, por otra, que se requieran dosis mayores para que se den mutaciones favorables en la dosis letal media, o deletéreas. Además, estos resultados concuerdan con lo reportado por Polheim (1981), quien consigna que la probabilidad de tener una mutación favorable se encuentra en dosis superiores a los 200 Gy y cercanas a los 300 Gy.
Número de ginóforos
La inducción de ginóforos fue estadísticamente diferente al 1% de probabilidad entre las dosis de radiación con un coeficiente de variación de 12.8%, como se presenta en la tabla 1. Al analizar el efecto que tuvieron las dosis de radiación en la inducción de ginóforos, los valores promedio que aparecen en la tabla 2 muestran que la dosis de 100 Gy no difiere significativamente con el testigo. En las dosis de 100 Gy y 200 Gy se presentó el fenómeno de reversión citado por Robles (1986), debido a que el número de ginóforos fue estadísticamente superior (p < 5%) con respecto a las dosis inmediatas inferiores (50 Gy y 150 Gy, respectivamente) como aparece en la figura 1. Por otra parte, la variación que se tuvo entre las dosis de radiación se clasificó en cinco grupos: el primero formado por el testigo y la dosis de 100 Gy; el segundo por la dosis de 200 Gy inferior 27% con respecto al testigo; el tercero creado por la dosis de 50 Gy 33% menor al testigo; el cuarto por la dosis de 150 Gy con una disminución de 45%; y el quinto por las dosis de 250 Gy y 300 Gy con el 50% de ginóforos con respecto al testigo. Además, la GR30 se presenta en dosis cercanas a los 150 Gy, mientras que la GR50 se ubica en dosis cercanas a los 200 Gy, rango donde se favorece la aparición de mutaciones útiles en los programas de mejoramiento genético (Ahmed & Mohamed, 2009). Asimismo, es significativo que en esta variable se haya manifestado el fenómeno de reversión en dos dosis (100 Gy y 200 Gy), y que esta última haya coincidido con la GR50. Este comportamiento puede deberse a que en las dosis mencionadas el efecto de la radiación haya ocasionado una menor alteración del DNA y, por consiguiente, la inducción de mutantes haya sido menor.
Por otra parte, se obtuvo una correlación significativa 1% (r = 0.49) entre el porte de la planta y el número de ginóforos, indicando que a mayor tamaño de planta, el número de ginóforos se incrementa. Ello explica que en las dosis mayores a 200 Gy los ginóforos disminuyeran significativamente, ya que las plantas a esas dosis presentaban un porte 60% menor en relación con el tratamiento sin irradiar. Al igual que en la altura de planta, la regresión fue negativa y altamente significativa, como se aparece en la tabla 3, con un coeficiente de determinación que indica que el 60% de los cambios en la inducción de los ginóforos se debieron a las dosis de radiación, y el resto a factores que no están relacionados con la radiación absorbida. Lo anterior explica el comportamiento positivo (y = 1.269X + 4.23) entre el porte y la inducción de ginóforos, indicando que a mayor altura se incrementan los ginóforos, por lo que en las dosis de 250 Gy y 300 Gy la diferencia no es significativa, debido a que el porte de la planta disminuyó conforme la dosis se incrementó, presentándose también una disminución drástica en el número de ginóforos por planta. De ello resultó que esta variable fuera severamente afectada, ya que los de menor tamaño no desarrollaron vaina. Estos resultados coinciden con lo reportado por Patil & Mouli (1980), encontrando una disminución en el número y longitud de las ramas y, por consiguiente, el número de ginóforos ocasionada por el efecto de la radiación.
Longitud de ginóforo
El análisis de varianza de la tabla 1 muestra diferencia altamente significativa entre los tratamientos, por lo que la radiación absorbida generó mutaciones que propiciaron cambios en la longitud de los ginóforos con una tendencia a reducir la longitud conforme la radiación se incrementó, como se muestra en la tabla 2 y la figura 1. Sin embargo, en los tratamientos de 50 Gy y 100 Gy, la longitud promedio de los ginóforos fue mayor que en tratamiento sin irradiar (0 Gy); mas no se pude asegurar que se haya presentado una reversión, ya que la diferencia entre estos tratamientos no fue significativa, por lo que la variación es atribuible a factores aleatorios y ajenos a la radiación, al igual que entre los tratamientos de 250 Gy y 200 Gy. Además, la reducción en la longitud conformó tres grupos: el primero donde no hay diferencia significativa entre el tratamiento sin irradiar y las dosis de 50 Gy, 100 Gy y 150 Gy, donde se presentó la mayor longitud de ginóforos; el segundo correspondió a las dosis de 200 Gy y 250 Gy que no manifestaron diferencia estadística; y el tercero a la dosis de 300 Gy con la menor longitud y el mayor número de ginóforos abortados y que no se desarrollaron vainas.
En las dosis de 200 Gy, 250 Gy y 300 Gy se presentaron plantas de porte muy bajo, donde los ginóforos fueron muy cortos que no se desarrollaron, por lo que abortaron o se necrosaron, indicando que se generaron mutaciones que ocasionaron cambios en la viabilidad de los ginóforos, lo que explica la disminución en su longitud. Por otra parte, no se presentó la GR50, por lo que se requieren dosis mayores a los 300 Gy, mientras que la GR30 que corresponde a una longitud promedio de 5.3 cm se ubica entre los 250 Gy y 300 Gy (tabla 2). Además, se obtuvo una correlación negativa y significativa al 1% de probabilidad, así como la regresión que explica la disminución en la longitud de los ginóforos conforme la dosis de radiación se incrementó. De modo paralelo, el coeficiente de determinación (R2 = 49%) indica que el 49% de la disminución en la longitud de los ginóforos se debió a las mutaciones que causaron las dosis de radiación conforme estas se incrementaron y el restante a factores ajenos, tales como el porte de la planta y otros relacionados con la inducción y el desarrollo de los ginóforos. Tomando en cuenta la correlación que hay entre el porte de la planta y el número de ginóforos, se explica el comportamiento que se presentó en los ginóforos de menor longitud, tanto en la falta de desarrollo como en los que abortaron o se necrosaron; además, el efecto negativo que ocasionó la radiación conforme esta se incrementó tuvo el mismo comportamiento en las dos variables.
Porciento de floración
Los tratamientos evaluados presentaron diferencia altamente significativa, como se muestra en la tabla 1, con un coeficiente de variación de 12% que confirma la validez de las varianzas. Al comparar el comportamiento de los tratamientos mediante DMS al 5% de probabilidad (tabla 2), se obtuvo una disminución en el número de plantas con presencia de flores conforme la dosis de radiación se incrementó, identificando tres grupos: el primero conformado por el tratamiento sin irradiar y las dosis de 50 Gy y 100 Gy con una floración promedio de 90%; el segundo incluye las dosis de 150 Gy, 200 Gy y 250 Gy con 60% de floración; y el tercero por la dosis de 300 Gy con solo el 36% de floración. Como se observa en la tabla 2, el porcentaje disminuye 30% entre el primero y el segundo grupo, así como entre el segundo y el tercero, demostrando que la floración se reduce significativamente a partir de la dosis de 150 Gy, que además coincide con la GR30.
Se obtuvo una correlación negativa y significativa (p < 0.01) entre las dosis de radiación y el porcentaje de floración, como aparece en la tabla 3, explicando el abatimiento conforme la dosis se incrementó. Asimismo, el coeficiente de determinación indica que el 77% de los cambios son atribuibles a la radiación absorbida y el 27% a factores ajenos a la radiación. La ecuación de regresión muestra que la GR50 se ubica en 28 Gy, mostrando la diferencia de 60% en el porcentaje de floración entre el primer grupo conformado por las dosis de 0 Gy, 50 Gy y 100 Gy y la dosis de 30 Gy; además, en esta última se presentaron plantas que indujeron ginóforos, pero no se desarrollaron las vainas. Hussein (1981) reporta que el sistema genético de la floración varía en tiempo en diferentes especies de plantas, y está basado en un amplio número de genes que difieren en su efecto, y el reconocimiento de estos loci es relativamente fácil en las mutaciones inducidas.
A pesar de que la dosis de 200 Gy es superior a 150 Gy, no se puede afirmar que se haya presentado un comportamiento reversivo, debido a que la diferencia en el porcentaje de floración no es estadísticamente significativa. Hussein (1981) reporta que esta variable está gobernada por genes recesivos, lo cual permite seleccionar en generaciones avanzadas genotipos precoces, favoreciendo al cacahuate tipo Virginia, ya que es de ciclo tardío.
Características de las quimeras
Las diferencias fenotípicas se establecieron con base en el testigo. Se presentaron plantas de porte erecto, ginóforos débiles y cortos, foliolos pequeños y numerosos, plantas arrocetadas que indujeron ginóforos, pero no se desarrollaron las vainas y retraso en la floración, así como albinas y/o cloróticas.
Selección de quimeras
La selección de quimeras se llevó en las dosis de 200 Gy, 250 Gy y 300 Gy, debido a que en todas las variables evaluadas mostraron diferencia significativa con respecto tanto al testigo como a las dosis de 50 Gy y 100 Gy; además, en las variables altura de planta y longitud de ginóforos, si bien no se presentó la GR50, la GR30 coincide con la GR50 en número de ginóforos y porciento de floración. Los mutantes seleccionados correspondieron a plantas erectas de porte medio, ginóforos grandes y con vainas basales, debido a que en las ramas axilares hay un incremento significativo en la probabilidad de encontrar mutantes favorables, ya que presentan más puntos mutagénicos de acuerdo con lo reportado por Polheim (1981) y Hussein (1981). Las vainas de las plantas seleccionadas se colocaron en bolsas, se etiquetaron, se secaron y se evaluará la M2 en ciclos posteriores.
Conclusiones
Las dosis de radiación tuvieron un efecto directo en altura de planta, inducción de ginóforos, su longitud y el porcentaje de floración, ya que se presentaron índices de reducción significativos en dosis iguales o superiores a 200 Gy. En los caracteres altura de planta y longitud de ginóforo no se presentó la GR50, por lo que se requieren dosis mayores; mientras que en inducción de ginóforos y porcentaje de floración se ubicó entre los 250 Gy y 300 Gy, presentando la GR30 en dosis mayores a 200 Gy. La inducción de ginóforos mostró un comportamiento reversivo en las dosis de 100 Gy y 200 Gy. Los mutantes seleccionados correspondieron a plantas erectas de porte medio, ginóforos grandes y con vainas basales.

