











 text new page (beta)
text new page (beta) Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La exposición a xenobióticos afecta cada vez más la biodiversidad de los ecosistemas acuáticos debido a que tienen el potencial de actuar como perturbadores endocrinos (PE). En hábitats acuáticos, los PE están fácilmente biodisponibles para los peces a través de una gran variedad de rutas de absorción tales como la vía respiratoria, la transferencia materna y la vía y oral. Los PE ocasionan severas alteraciones e interfieren en los sistemas endócrino reproductivo e inmune causando una gran variedad de trastornos en el desarrollo y la reproducción (Filby et al. 2007, Breves et al. 2018).
Los PE se encuentran ampliamente distribuidos, son muy persistentes y pueden acumularse debido a procesos de biomagnificación (Aris et al. 2014). Estos compuestos se caracterizan, en general, por ser lipofílicos (no solubles en agua), persistentes en el ambiente (vidas medias largas) y bioacumulables, algunos de ellos incluyen algunos plaguicidas (fungicidas, herbicidas e insecticidas), sustancias químicas industriales, productos sintéticos y algunos metales tóxicos (Kime et al. 1999, Siegenthaler et al. 2017). Entre las hormonas sintéticas, se encuentra el componente estrogénico de los anticonceptivos orales, el 17α-etinilestradiol (EE2), cuya presencia en aguas residuales, efluentes y aguas receptoras es de 10 a 50 veces mayor que la de otros estrógenos naturales (Verslycke et al. 2002, Segner et al. 2003, Zhang 2009, Velasco-Santamaría et al. 2010). La vida media de eliminación en la mujer es de unas 25 horas, siendo metabolizado inicialmente en el hígado por hidroxilación aromática, formándose una gran variedad de metabolitos hidroxilados y metilados. Estos metabolitos se presentan de forma libre o conjugada como glucurónidos o sulfatos, el 70 % del EE2 conjugado se excreta en la orina y un 30 % es eliminado por las heces. Los metabolitos conjugados excretados por heces u orina pueden experimentar posteriormente procesos de desconjugación durante los tratamientos a los que se someten las aguas residuales (Desbrow 1998). Se ha descrito que en peces el EE2 se ve sometido a la circulación enterohepática, amplificando su efecto tóxico debido a una redosificación continua (Schultz et al. 2003).
El EE2 es una sustancia relativamente persistente en el medio acuático, aparece de forma frecuente y amplia en aguas superficiales con concentraciones detectables en el rango de 0.05 a 831 ng/L (Ternes 1999, Baronti 2000, Huang 2001 y Kolpin 2002). Aunque esas concentraciones son relativamente bajas en aguas superficiales, su potencia perturbadora es mayor que la de otros estrógenos (Aris et al. 2014). Se ha reportado que produce alteraciones en los machos, tales como incremento de la vitelogénesis (inducción de vitelogenina), feminización y hermafroditismo (gónadas masculinas y femeninas en un solo individuo), disminución del tamaño de las gónadas, baja fecundidad, alteración en las características seminales y cambios de comportamiento reproductivo (agresividad, cortejo y territorialidad), alteraciones que pueden llegar a producir un colapso de las poblaciones (Hansen 1998, Aris et al. 2014).
En la región de la Orinoquia, A. metae está siendo utilizada como modelo de estudios ecotoxicológicos por su gran potencial como bioindicador y se han demostrado resultados interesantes en otras variables de respuesta (Corredor-Santamaría et al. 2016, 2019, 2020, Velasco-Santamaría et al. 2019). A. metae de la familia Cichlidae, se encuentra distribuida en la mayoría de los cuerpos de agua de América del Sur, con hábitos detritivoros es omnívora, de pequeño tamaño, tiene ciclos de vida con alta dinámica reproductiva y ha demostrado resistencia a cuerpos de agua contaminados (Corredor-Santamaría et al. 2016, 2019, 2020, Velasco-Santamaría et al. 2019). Estas características la convierten en una especie apropiada para ser utilizada como bioindicador y como modelo biológico en la región de la Orinoquia colombiana. De esta manera, el objetivo de esta investigación es conocer los efectos causados por el EE2, un potente perturbador endócrino en adultos de A. metae.
MATERIALES Y MÉTODOS
Material biológico
Se utilizaron 216 adultos (108 hembras y 108 machos), clínicamente sanos, con longitud corporal aproximada de 7.5 ± 5.0 cm y peso corporal de 7.2 ± 5.0 g, criados en estanques en tierra, provenientes de la estación piscícola del Instituto de Acuicultura de los Llanos (IALL), Universidad de los Llanos, Villavicencio-Meta-Colombia.
Diseño experimental
Se utilizaron peces aclimatados en tanques plásticos (400 L) durante un periodo de 30 días y posteriormente 10 días en acuarios de vidrio (20 L), con temperatura del agua promedio de 27 ± 5.0 ºC, monitoreada diariamente, y con fotoperiodo 12L:12O. Cada tratamiento (T) estuvo conformado por seis peces y tres réplicas cada uno, por sexos separados, los cuales fueron pesados (g) y medidos (cm) antes del experimento.
Los peces fueron expuestos durante 21 días a cuatro concentraciones de EE2 disuelto en el agua, denominadas T1: 0.5, T2: 5, T3: 50, T4: 250 ng/L, testigo negativo (C) y testigo con solvente (etanol) (CE) al 0.1 %. Los peces fueron alimentados a saciedad, dos veces al día (mañana y tarde) con alimento balanceado comercial (Mojarra 30 %, Italcol). Se realizaron recambios de agua con limpieza de fondo del acuario, eliminando heces y residuos de comida cada tercer día con renovación del 20 % del agua con las concentraciones iniciales de EE2.
17a-etinilestradiol (EE2)
El EE2 se adquirió de Sigma-Aldrich Chemie GmbH (Steinheim, Alemania, Cat. No E4876), diluido en etanol (Sigma-Aldrich) manteniendo una disolución patrón de 0.1 mg/mL.
Muestreo
Finalizado el tiempo de exposición al EE2, los peces de cada tratamiento fueron capturados y anestesiados con 2-fenoxietanol (J. T. Baker, Phillisburg, EUA) a 300 ppm hasta la pérdida de su eje de nado; se pesaron (balanza digital Ohaus Scout Pro®) y midieron (ictiómetro). Posteriormente, se tomaron muestras de sangre con jeringas de insulina previamente heparinizadas mediante punción de la vena caudal. La sangre fue sometida a centrifugación (12 000 rpm) durante cinco minutos (Hettich centrifuguen EBA 12, Fohrenstr, Alemania) y, el plasma fue almacenado a -20 ºC hasta su posterior análisis. Finalmente, se realizó la disección de órganos para extraer las gónadas y el hígado y llevar a cabo los análisis de índices somáticos e histológicos. Las gónadas y el hígado fueron extraídos y pesados para determinar el índice gonadosomático (IGS) y el hepatosomático (IHS), calculados como: (peso órgano / peso corporal) × 100. Posteriormente, una porción de cada órgano (gónada e hígado), se fijó en solución comercial Bouin (Sigma) para el análisis histológico, en el que se evaluaron seis peces por tratamiento, tres por cada sexo.
Cuantificación de calcio y fósforo plasmático por el método de Elisa
La cuantificación de calcio y fósforo plasmático se realizó mediante la técnica Enzyme-Linked Immuno Sorbent Assay, kit Elisa Spinreact de inmunoensayo, siguiendo el procedimiento de análisis y de lectura de acuerdo con las instrucciones del fabricante. Para determinar calcio se usó el kit BSIS08-E (Spinreact) y para fósforo se usó el kit BSIS15-E (Spinreact).
Se utilizó un espectrofotómetro de placa ajustado a una longitud de onda de 570 nm (BioTek Cytation 3) a temperatura de 37 ºC, como blanco se utilizó agua destilada y se estandarizó el protocolo utilizando 10 μL de muestra para calcio y 5 μL para fósforo. Posteriormente, se mezcló e incubó durante cinco minutos a 37 ºC y se realizó la lectura de la absorbancia (A), del patrón y de la muestra, frente al blanco del reactivo después de ٤٠ minutos. Los resultados fueron ajustados de acuerdo con la siguiente fórmula:
Calcio plasmático = (A) Muestra - (A) Blanco / (A) Calibrador - (A) Blanco × 10 = mg/dL × 0.323 = mmol/L de calcio en la muestra.
Fósforo plasmático = Muestra / Patrón × 5 = mg/dL × 0.323 = mmol/L de fósforo en la muestra.
Histología gonadal y hepática
Las muestras de gónada e hígado fueron sometidas a procesos de hidratación y deshidratación con etanol (70 a 100 %) hasta ser incluidas en Paraplast (Fisherbrand, Fisher, WA, EUA). Se realizaron cortes histológicos seriados de 4 μm en un micrótomo Leica y fueron sometidas a tinción con hematoxilina y eosina (H&E) para evaluar la estructura general y la morfología de los tejidos. Tanto en ovario como en testículo se evaluó la estructura general del órgano, considerando alteraciones celulares, acumulación de material de melanomacrófagos, degeneración testicular, atresia, fibrosis intersticial, congestión, material vitelogénico y apoptosis.
Las evaluaciones de hallazgos histológicos en ovario y en testículo fueron realizadas siguiendo los criterios de Velasco-Santamaría et al. (2010, 2011). Las alteraciones primarias (por ejemplo, sincitios de células germinales), se clasificaron en cuatro grados diferentes: mínimo (grado 1, menos de 20 % de alteraciones), leve (grado 2, entre 20 y 50 % de alteraciones), moderada (grado 3, entre 50 y 80 % de alteraciones) y severa (grado 4, más de 80 % de alteraciones). Por su parte, las alteraciones secundarias (por ejemplo, proporción alterada de espermatocitos) utilizando la clasificación de presente / ausente.
El análisis de histología hepática se realizó de forma cuantitativa de acuerdo con la gravedad de las lesiones y su distribución (focal, multifocal y difusa), siendo 0 = no alteraciones evidentes, 1 = leve; 2 = moderado y 3 = severo, implicando una alteración significativa de la apariencia o la arquitectura (por ejemplo, actividad picnositaria degenerativa, activación de macrófagos, apoptosis, infiltración de células inflamatorias, vacuolización, dilatación y congestión). (Velasco-Santamaría et al. 2010, 2011).
Análisis estadístico
Se realizó un diseño completamente al azar. Los datos fueron expresados como media ± error estándar (SEM). Las pruebas de homogeneidad de varianzas y distribución normal se determinaron por medio de la prueba de Bartlett y Kolmogorov-Smirnov, respectivamente. Para determinar las diferencias estadísticas significativas entre los tratamientos se realizó un análisis de varianza (ANOVA) seguido del post-test de Tukey. En todos los casos el valor de p < 0.05 fue utilizado para revelar diferencias significativas. Los procedimientos estadísticos se realizaron por medio del paquete SAS para Windows versión 9.02 (2002-2006, SAS Institute Inc., Cary, NC, EUA).
RESULTADOS
Índices somáticos
En los resultados de los índices gonadosomático y hepatosomático obtenidos en la exposición a EE2 en hembras y en machos no se encontraron diferencias estadísticas significativas entre los tratamientos en comparación con los grupos testigo (Fig. 1).
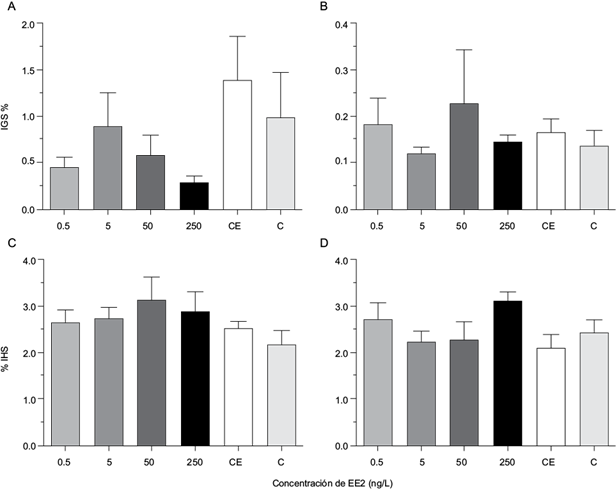
Fig. 1 Porcentaje del indice gonadosomático (IGS) en A) hembras y B) machos y porcentaje de índice hepatosomático (IHS) en C) hembras y D) machos adultos de Aequidens metae expuestos a diferentes concentraciones de EE2 durante 21 días, valores expresados con media error ± estándar. No se observaron diferencias significativas con respecto al testigo (p<0.05), N=6. Testigo negativo (C), Solvente testigo con etanol (CE).
Cuantificación de calcio y fósforo plasmático
En cuanto a los niveles de calcio plasmático, en hembras no se encontraron diferencias estadísticas entre tratamientos ni en relación con los grupos testigo. En machos se encontró un incremento significativo en los tratamientos T2 (5 ng/L) con 2.06 ± 0.09 mmol/L, T3 (50 ng/L) con 2.22 ± 0.06 mmol/L y T4 (250 ng/L) con 2.33 ± 0.1 mmol/L con respecto al testigo C (1.48 ± 0.17 mmol/L). Entre los demás tratamientos no se encontraron diferencias significativas (p > 0.05) (Fig. 2).

Fig. 2 Porcentaje de calcio plasmático en hembras (A) y machos (B) de Aequidens metae expuestos a diferentes concentraciones de EE2, valores expresados con media ± error estándar de la media. Entre tratamientos, valores con letras distintas son significativamente diferentes (p<0.05), N=6. Testigo negativo (C), Solvente testigo con etanol (CE).
En los niveles de fósforo plasmático, en hembras no se encontraron diferencias significativas entre tratamientos ni en relación con los grupos testigo. En machos, se observó un incremento significativo en el tratamiento T4 (250 ng/L) con 5.83 ± 1.09 mmol/L comparado con los peces del grupo testigo CE 3.096 ± 0.33 mmol/L. Entre los demás tratamientos no se encontraron diferencias significativas (p>0.05) (Fig. 3).
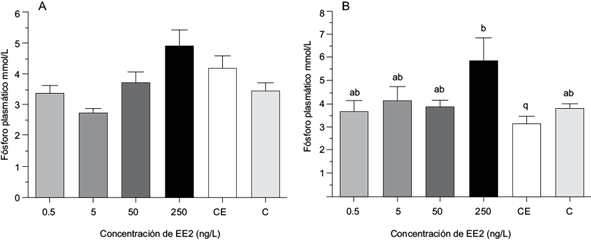
Fig. 3 Porcentaje de fósforo plasmático (mmol/L) en hembras (A) y machos (B) de Aequidens metae expuestos a diferentes concentraciones de EE2, valores expresados con media ± error estándar. Entre tratamientos, valores con letras distintas son significativamente diferentes (p<0.05), N=6. Testigo negativo (C), Solvente testigo con etanol (CE).
Análisis histológico
A nivel gonadal, se observó un incremento significativo de alteraciones histológicas con respecto a los grupos testigo. En ovario, en el tratamiento T1 (0.5 ng/L) se observó congestión y presencia de material vitelogénico en grado 1 (mínimo), en el T2 (5 ng/L) se evidenció apoptosis y congestión en grado 2 (leve), fibrosis intersticial, material vitelogénico y atresia folicular en grado 1 (mínimo). En el T3 (50 ng/L) se observó apoptosis, congestión, fibrosis intersticial, atresia folicular y material vitelogénico en grado 2 (leve) y, en T4 (250 ng/L) se encontró mayores contenidos de material vitelogénico, así como fibrosis intersticial, apoptosis, congestión y atresia folicular en grado 3 (moderado), ver fig. 4.
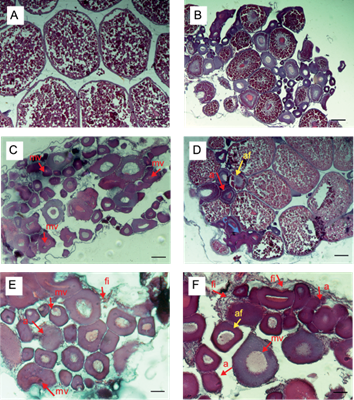
Fig. 4 Microfotografías de histología gonadal en hembras de Aequidens metae con H&E. A. Testigo negativo: estructura normal de ovario maduro con oocitos en fase vitelogénica. 200x B. Solvente testigo con etanol: ovario maduro con oocitos en fase perinuclear, alveolo cortical y vitelogénico. 100x. C. T1 (0.5 ng/L EE2) mínima pérdida de estructura y congestión con presencia de material vitelogénico (mv).100x. D. T2 (5 ng/L EE2) pérdida de estructura, atresia folicular (af), congestión (c), apoptosis (a). 200x. E. T3 (50 ng/L EE2) leve presencia de material vitelogénico, apoptosis, fibrosis intersticial (fi). 200x. F. T4 (250 ng/ L EE2) presencia de material vitelogenico, apoptosis, fibrosis intersticial, y atresia folicular. 200x. Se encontró un incremento significativo con respecto al testigo (p<0.05). Escala 5 µm. N= 3.
En testículo, en el T1 (0.5 ng/L) se observó presencia de material de melanomacrófagos, degeneración testicular y fibrosis intersticial en grado 1 (mínimo). En peces expuestos al T2 (5 ng/L) y T3 (50 ng/L) se observó fibrosis intersticial, material de melanomacrófagos y degeneración testicular en grado 2 (leve). En el T4 (250 ng/L) se observó material de melanomacrófagos en grado 2 (leve), degeneración testicular y fibrosis intersticial en grado 3 (moderado) (Fig. 5).
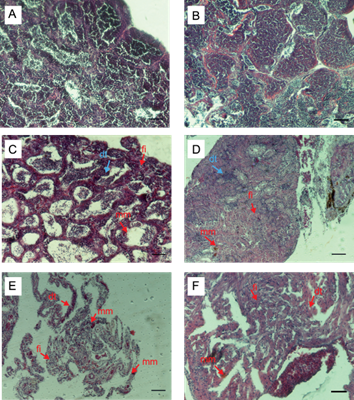
Fig.5. Microfotografías de histología gonadal en machos de Aequidens metae con H&E. A. Testigo negativo y B. Solvente testigo con etanol: estructura normal de testículos maduros. 200x. En C. T1 (0.5 ng/L EE2) presencia de material melanomacrófago (mm), degeneración testicular (dt), fibrosis intersticial (fi). 200x. D. T2 (5 ng/L EE2) leve incremento de material melanomacrófago, degeneración testicular y fibrosis intersticial. 100x. E. T3 (50 ng/L EE2) material melanomacrófago, degeneración testicular, fibrosis intersticial. 100x. F. T4 (250 ng/L EE2) moderada presencia de material melanomacrófago, degeneración testicular, fibrosis intersticial. 100x. Se encontró un incremento significativo con respecto al testigo (p<0.05). Escala 5 µm. N= 3.
A nivel hepático, en ambos sexos, se encontró un incremento significativo de alteraciones histológicas con respecto a los grupos testigo, en el T1 (0.5 ng/L) se observó actividad picnocitaria grado 1 (leve). En el T2 (5 ng/L) se observó apoptosis, actividad picnositaria, infiltraciones y dilatación de núcleos en grado 1 (leve). En el T3 (50 ng/L) se observó congestión, apoptosis, activación de macrófagos, dilatación de núcleos e infiltraciones grado 2 (moderado) y en el T4 (250 ng/L) se observó apoptosis, congestión, infiltraciones y dilatación de núcleos en toda la estructura en grado 3 (severo) (Fig. 6).
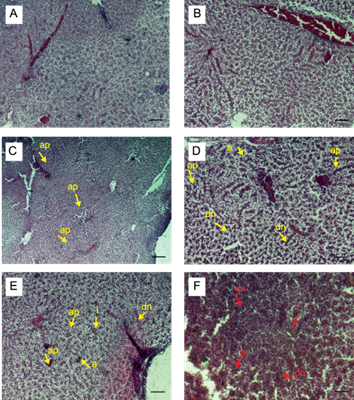
Fig. 6 Microfotografías de hígado de Aequidens metae con H&E. en A. Testigo negativo y B. Solvente testigo con etanol: estructura normal del componente hepático. 200x. C. T1 (0.5 ng/L EE2), presencia de actividad picnositaria (ap) 100x. D. T2 (5 ng/L EE2), apoptosis (a), dilatación de núcleos (dn) y actividad picnositaria (ap) 200x. E. T3 (50 ng/L EE2), moderada congestión, infiltraciones (i), dilatación de núcleos y apoptosis (a). 200x. F. T4 (250 ng/L EE2), severa congestión, apoptosis, infiltraciones y dilatación de núcleos. 200x. Se encontró un incremento significativo con respecto al control (p<0.05). Escala 5µm. N= 6.
DISCUSIÓN
En este estudio, los índices somáticos no muestran diferencias significativas en hembras y en machos. En el IGS no se encontraron diferencias estadísticas significativas. Varios autores reportan un efecto negativo y se asume que, valores de IGS más bajos de lo normal indican un desvío de la energía que habitualmente se usa para el crecimiento de las gónadas. El IGS se encuentra influenciado por factores ambientales como son los cambios de fotoperiodo, temperatura, concentración de oxígeno disuelto, niveles de agua, contaminación, estimulación acústica por la lluvia, disponibilidad de comida y densidad de la población (Goede y Barton 1990).
En el presente estudio, en el IHS en hembras y en machos no se encontraron diferencias significativas, estos resultados coinciden con Cabrera-Páez et al. (2008) y Thorpe et al. (2003). En contraste, estudios realizados por Porter y Janz (2003) y Van Dyk et al. (2012), reportan un incremento de IHS por consecuencia de la exposición a EE2. Otros autores afirman que el incremento en el IHS puede deberse a la hiperplasia o a la hipertrofia como una respuesta adaptativa para ampliar la capacidad de desintoxicación de compuestos extraños (Arcand-Hoy y Metcalfe 1999). Por su parte, Carragher y Sumpter (1990) reportan que la hipertrofia en el hígado se debe a la inducción de la vitelogenina por la presencia de tóxicos en el organismo. Sin embargo, también depende de la especie y del grado de tolerancia que presente a dicho compuesto, siendo esta una de las posibles explicaciones, ya que en estudios previos con A. metae se ha evidenciado esta tolerancia (Corredor-Santamaría et al. 2016, 2019).
A nivel histológico, en ovario se observó presencia de material vitelogénico en su estructura, apoptosis, congestión, fibrosis intersticial y atresia folicular. Estos resultados son similares a los obtenidos por Papoulias et al. (2000), quienes reportaron presencia de material vitelogénico en Oryzias latipes después de siete días a la exposición a 0.05 ng/L de EE2. A nivel de testículo, en el presente estudio se observó material de melanomacrófagos, degeneración testicular y fibrosis intersticial a partir de concentraciones de 5 ng/L de EE2, lo que coincide con lo reportado por Pérez et al. (2012) en Odontesthes bonariensis, a concentraciones de 0.5 ng/L quienes reportaron fibrosis intersticial, apoptosis y degeneración testicular. En estudios realizados por Velasco-Santamaría et al. (2011) en viruelas (eelpout Zoarces viviparus) expuestos a concentraciones de 5 ng/L se reportó hiperplasia folicular y fibrosis intersticial. De igual forma, los resultados de Cabas et al. (2011) son similares en concentraciones de 5, 50, 125 y 200 µg/g de EE2 suministrado en el alimento después de 7, 14 y 21 días. En estudios realizados por Silva de Assis et al. (2013) en concentraciones de 135 ng/L y 231 ng/L se reportó perdida de la estructura celular e infiltración de células.
Otro órgano de estudio considerado blanco para detectar efectos de contaminación es el hígado. Por tanto, en esta investigación, en los peces expuestos a concentraciones de 0.5 ng/L tanto en hembras como en machos, se encontró actividad picnositaria. En los peces expuestos a 5 ng/L, 50 ng/L y 250 ng/L se observó dilatación de núcleos, activación de macrófagos, apoptosis, infiltraciones y congestión en diferente grado de afectación, estos resultados son similares a los reportados por Van-Dyk (2012), Velasco-Santamaría et al. (2011) y Paulino et al. (2017) quienes encontraron estos mismos efectos en peces expuestos a otros xenobióticos que también tienen el potencial de actuar como perturbadores endócrinos. El hígado en los peces realiza funciones hepáticas y pancreáticas y es el principal órgano metabolizador de los compuestos que le llegan por vía sanguínea, por tanto, este órgano daño tisular causado por contaminantes del ambiente como pesticidas y metales pesados, entre otros. Aunque la histología hepática es una herramienta ampliamente utilizada en los estudios de PE, son escasos los reportes que han evaluado el efecto de la exposición a EE2.
En los peces marinos, una fuente abundante de calcio (Ca) está presente en el ambiente y, los alimentos sólo son esenciales para satisfacer la demanda de fósforo (P). En contraste, en los peces de agua dulce, los iones de Ca son escasos en el ambiente y la dieta contribuye a satisfacer los requerimientos de Ca y P. Si el Ca se convierte en limitante, puede comprometer la reproducción, el crecimiento y el desarrollo, ya que se induce la movilización tanto del Ca como del P de tejidos mineralizados, siendo estos importantes en la homeostasis corporal (Persson et al. 1997). En este estudio, los resultados obtenidos con calcio plasmático en hembras no presentaron diferencias significativas con respecto al grupo testigo. Para el caso de los machos, se encontraron diferencias significativas en los peces expuestos a concentraciones mayores a 5 ng/L de EE2 con respecto al grupo testigo. Estos resultados son similares a los obtenidos por Shirdel et al. (2016) con un incremento en el porcentaje de calcio plasmático en machos expuestos a nonilfenol, otro perturbador endocrino liposoluble de gran importancia.
Con respecto al fósforo plasmático, en hembras no se observaron diferencias significativas entre tratamientos. En machos se encontró diferencias significativas en aquellos expuestos a 250 ng/L comparado con el grupo CE. Estos resultados sugieren que algunos PE incluidos los contaminantes ambientales, como el EE2, bisfenol A (BPA), 2,3,7,8 tetraclorodibenzo p-Dioxina (TCDD), PCB 77 y el antagonista de estrógenos, ZM189,154 (ZM) pueden tener un impacto en el desarrollo del esqueleto, morfología y anomalías en los peces (Boudreau et al. 2004, Warner et al. 2007). La exposición de los peces a aguas contaminadas en las plantas de tratamiento de aguas residuales con posibles efectos estrogénicos causó cambios detectables en la morfología de la los arcos hemales (Rauson et al. 2008).
El estudio de los efectos de los PE es necesario para prevenir futuros disturbios en las especies de peces, la vida silvestre y el bienestar humano en general. Más específicamente, la perturbación estrogénica de los tejidos mineralizados puede tener una amplia gama de consecuencias, ya que un aumento en las anomalías esqueléticas, la densidad ósea modificada y la homeostasis mineral pueden afectar la natación y la capacidad de capturar presas y escapar de los depredadores. Además, el papel protector de las escamas y su contribución a la homeostasis mineral principalmente del Ca y el P pueden verse comprometidos y afectar la aptitud y productividad de los peces.
CONCLUSIONES
Los resultados obtenidos en la presente investigación demostraron que la exposición a concentraciones de EE2 similares a las reportadas en el ambiente, afectaron la histología e integridad de las gónadas y del hígado de los peces de la especie A. metae expuestos, así como los niveles de los iones de calcio y fósforo plasmáticos.

