










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista internacional de contaminación ambiental
Print version ISSN 0188-4999
Rev. Int. Contam. Ambient vol.31 n.1 Ciudad de México Feb. 2015
Microbial community and pollutantns survey in sediments of biologically important wetlands in Lerma, Mexico
Evaluación de contaminantes y de la comunidad microbiana en sedimentos de importancia biológica de los humedales de Lerma, México
Arturo Aburto-Medina1*, Derik Castillo1, Irmene Ortíz2, Ernesto Hernández3, Rurik List1 and Eric Adetutu4
1 Departamento de Ciencias Ambientales, Universidad Autónoma Metropolitana Lerma, Av. Hidalgo Pte. 46, Col. La Estación, Lerma de Villada, Estado de México, C.P. 52006
2 Departamento de Procesos y Tecnología, Universidad Autónoma Metropolitana Unidad Cuajimalpa, Av.Vasco de Quiroga 4871, Col. Santa Fe Cuajimalpa, Cuajimalpa de Morelos, México, D.F., C.P. 05300
3 Centro Nacional de Investigación y Capacitación Ambiental (CENICA), Periférico 5000, Col. Insurgentes Cuicuilco, Coyoacán, México, D.F., C.P. 04530
4 Royal Melbourne Institute of Technology, RMIT University, GPO Box 2476, Melbourne VIC 3001, Australia *Autor de correspondencia: aarturo1309@gmail.com
(Received March 2014;
accepted October 2014)
ABSTRACT
Wetlands pollution is of great concern given their importance to conservation and as water and food sources for the local population. Therefore, microbial and chemical investigations were carried out on the Lerma (Chimaliapan) and Almoloya del Río (Chiconahuapan; both Ramsar sites) wetland samples in Mexico for risk assessment purposes. 16S rRNA-based clone library analyses showed the prevalence of Entero-bacteriaceae, specifically the genus Shigella and E. coli species in both wetlands. While faecal coliform levels in both wetlands were below the accepted limit, higher total coliform counts (> 2400 MPN) were observed in ~ 40 % of the sampled sites at Lerma. Other microorganisms detected included organisms similar to those obtained from hydrocarbon-contaminated environments with alkanes and poly-aromatic hydrocarbons being detected in the sediments. Detected metals were below United States Environmental Protection Agency (USEPA) limits, decreasing in the sequence: Al > Fe > Mn >V> Zn > Pb > Ni > Cr > Cu > Co > Tl > As > Be (except Cr at Almoloya). However, Al concentrations were significantly above the permissible limits (2700 mg/kg) of the National Oceanic and Atmospheric Administration of the United States in both wetlands. This study therefore demonstrates that increased health risks and potential for metal biomagnification by edible species could be associated with the use of wetland water resources.
Key words: microbial diversity, metals, semimetals, wetlands, conservation, marsh.
RESUMEN
La contaminación de las ciénegas es de gran relevancia debido a su importancia en la conservación del ambiente y a que son fuente de recursos para las poblaciones locales. Por lo tanto, realizamos análisis químicos y microbiológicos en muestras de las ciénegas de Lerma (Chimaliapan) y Almoloya del Río (Chiconahuapan; ambos sitios Ramsar) ubicadas en México para estimar el riesgo que pueden representar estos cuerpos de agua debido a la contaminación. Librerías de clones basadas en el gen 16S rRNA demostraron la dominancia de miembros de Enterobacteriaceae, específicamente especies parecidas a E. coli y del género Shigella en ambas ciénegas. Aunque en general los niveles de coliformes fecales estuvieron dentro de los límites permisibles en ambas ciénegas, los números de coliformes totales excedieron la norma (>2400 NMP) en ~40 % de los sitios muestreados en la ciénega de Lerma. Otros microorganismos detectados fueron similares a aquellos observados en sitios contaminados con hidrocarburos, específicamente con alcanos e hidrocarburos poli-aromáticos detectados en los sedimentos. Los valores de metales estuvieron debajo de los límites permitidos por la Agencia de Protección Ambiental de los Estados Unidos (USEPA) y disminuyeron de la siguiente forma: Al > Fe > Mn >V> Zn > Pb > Ni > Cr > Cu > Co > Tl > As > Be, (excepto Cr en Almoloya). Sin embargo, las concentraciones de Al estuvieron significativamente por arriba de los límites permitidos (2700 mg/kg) por la Administración Oceánica y Atmosférica de los Estados Unidos en ambas ciénegas. Este estudio por lo tanto, demuestra que un incremento en los riesgos a la salud y el potencial de biomagnificación de metales podrían estar asociados al uso de los recursos de las ciénegas.
Palabras clave: diversidad microbiana, metales, semimetales, humedales, conservación, ciénega.
INTRODUCTION
At the time of the Spanish conquest, the highlands of central Mexico had large wetland systems that covered significant portions of the valleys and closed basins. Gradually, these were drained and converted into agricultural lands and urbanized. According to The Ramsar Convention on Wetlands, wetlands are areas of marsh, fern, peat land or water, whether natural or artificial, permanent or temporary, with water that is static or flowing, fresh, brackish or salt, including areas of marine water of which the depth at low tide does not exceed six metres (Halls 1997). The Ramsar convention lists 25 important wetlands in Mexico and one of them is the system of marshes at the headwaters of the Lerma river, that are located in the valley of Toluca, central Mexico. These marshes cover 3000 ha and are the last remnants of a former 27 000 ha lake, which drained over several decades, caused the loss of biodiversity and the extinction of at least one species, the Lerma Grackle (Quiscalus palustris). However, despite their reduced size, these marshes provide raw materials for centuries' old handcraft tradition of the manufacture of reed products, and local people obtain food and forage from the marshes. They also play important roles in flood control, are host to nearly 100 resident and migratory bird species (Vázquez 2004) and are refuge to endemic and endangered fish, birds, and a micro-endemic and critically endangered amphibian Ambystoma lermaense (Lyons et al. 1995, Aguilar and Casas 2009, Colón et al. 2009). A wide variety of aquatic plants, including endemic species (Ramos 1999) are also found in these wetlands. Recent studies have identified the aquatic vascular flora and changes in plant diversity (Zepeda-Gómez et al. 2012) as well as the ecology of epilithic diatoms (Segura-García et al. 2012). As a result of their importance, these marshes were designated a federally protected area (Refugio de Flora y Fauna Silvestre Ciénegas de Lerma; SEMARNAT 2002).
Given that the marshes are sources of water and direct and indirect food resources to local people (Pérez-Ortiz and Valdez 2006), pollution of water and sediments may pose health hazards. Marshes can experience an advanced process of pollution due to industrial, agricultural and municipal run offs and a decline of water volumes due to aquifer exploitation (Esteller and Díaz-Delgado 2002, Gómez-Salazar 2012). The pollution in the Lerma River and the Alzate dam is mainly due to excessive amounts of organic matter and heavy metals. Previous studies have measured the metal concentrations in sediments of the river (Tejeda et al. 2006, Zarazúa et al. 2011) and the Alzate dam (Ávila-Pérez et al. 1999) and most of them are above the USEPA contamination threshold. Another study has established the ecological risk of nine metals (López-Galván et al. 2010) in different zones of the basin and Cu and Pb showed the highest risks in the dissolved phase (Gómez-Salazar 2012).
However, there is no information of the contaminants' identity or the microbial communities in these wetlands. Therefore this study aims to characterize the microbial community in the wetlands (Almoloya and Lerma) for health risk assessment purposes and to identify the organic and inorganic contaminants in sediments obtained from the marshes of these wetlands in order to evaluate potential impacts of anthropic activities.
MATERIALS AND METHODS
Site description and sampling
Samples were collected from the Almoloya del Río (Chiconahuapan) and Lerma (Chimaliapan) marshes in central Mexico. Detailed descriptions of the marshes have been previously reported (Zepeda-Gómez et al. 2012). The geographic coordinates for each of the sampled sites and the relative position of the samples in the wetlands are indicated in table I and figure 1, respectively. Sampling points were selected in order to cover most areas of each wetland with sampling being carried out during the rainy season that spans from late May to early October (annual precipitations range between 800 to 1200 mm). Water samples were collected in sterile 1-litre glass bottles. Sediments from the bottom of the wetlands were collected in 50 mL sterile centrifuge tubes in triplicates after which, they were frozen with dry ice and protected from light. Domestic and industrial wastewater is discharged into both wetlands although the vast majority is discharged mainly into the river.
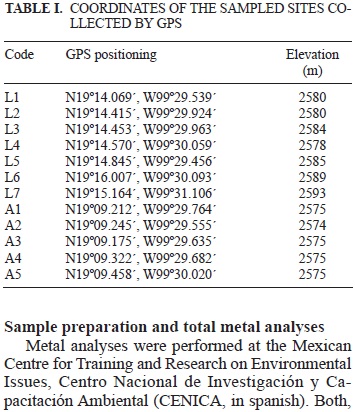
Sample preparation and total metal analyses
Metal analyses were performed at the Mexican Centre for Training and Research on Environmental Issues, Centro Nacional de Investigación y Capacitación Ambiental (CENICA, in spanish). Both, Lerma and Almoloya sediments were air-dried prior to analysis and sieve was not required. For all analyses, ultrapure deionized (nanopure) water was used throughout. All reagents were of analytical grade or higher purity. A rigorous quality control system was used including reagent blanks, duplicate samples and certified international reference materials (Channel Sediment BCR 320R from BCR). Precision and accuracy were better than 10 % for all analyzed components and the blanks were below the method detection limit (MDL).
Trace elements in sediments
Total trace elements were determined in 0.5 g of dried sediment using inductively coupled plasma atomic emission spectroscopy (ICP - AES) iCAP 6500, Thermo Scientific according to the USEPA 6010 method (USEPA 2007a). This was carried out after microwave assisted acid digestion in an oven Anton Paar, MULTIWAVE 3000 model, following the USEPA 3051 procedure (USEPA 2007b).
Identification of semivolatile organic compounds
The organic compounds were extracted from dried sediments by sonication (method 3550C, USEPA 2007c) using 3 g of soil and 95 mL of a mixture 1:1 n-hexane-ketone (Mallinckrodt, HPLC grade). The extracts were concentrated by roto-evaporation and changed to n-hexane as a solvent. The recovery percentage of this extraction method was validated using a soil sample with known concentrations of 17 reference compounds. CENICA provided this spiked sample and the percentage of recovery of all compounds was higher than 90 %. Regular controls with this soil were performed to ensure the quality of the data. The extracts were analyzed to identify organic compounds by gas chromatography (method 8270D, USEPA 2007d) coupled with a mass spectrum detector (Agilent 6890N, MSD 5975B, USA) using a 5 MS column (Agilent, USA). The temperatures of the detector and injector were 250 °C and 220 °C, respectively. The initial and final temperatures of the oven were 70 °C and 250 °C at a rate of 7 °C/min. Helium was used as the carrier gas and the mass scan range was from 50 to 450 z/m at 70 eV. Identification was made using the NIST05 Mass Spectral Library.
Statistical analyses
The statistical comparison of all heavy metal concentrations in sediments between both wetlands was performed using Hotelling's T2 (Rencher 2002), that corresponds to the multivariate version of the Student's t test.
Determination of total and faecal coliforms
The most probable number (MPN) of total coliforms, faecal coliforms and Escherichia coli followed the Mexican Norm NMX-AA-42-1987, which agrees with the ISO norm: ISO/DP 9308/2.
DNA extraction and PCR
Microbial community DNA was extracted directly from the slurries collected on September 21 using the UltraClean Soil DNA kit (MoBio Laboratories, Solana Beach CA) and PCR amplification of 16S rDNA genes was performed with the following primer pairs: 27F (TCT GGT TGA TCC CGC CAG) and 1392R (ACG GGC GGT GTG TAC; Lane 1991) for members of the Archaea, and 63F (CAG GCC TAA CAC ATG CAA GTC) and 1389R (ACG GGC GGT GTG TAC AAG; Marchesi et al. 1998) for the Bacteria as described previously (Aburto et al. 2009). The cycling conditions for both primer pairs were as follows: 1 cycle at 94 °C for 2 min, 30 cycles of 1 min at 94 °C, 1 min at 55 °C and 2 min at 72 °C and a final elongation at 72 °C for 10 min.
Cloning and sequencing
PCR products were cloned with a CloneJet cloning kit (Thermo) as described in the manufacturer's instructions using One Shot TOP10 chemically competent E. coli cells. Recombinant colonies were identified as white colonies and recovered from LB agar plates containing ampicillin (50 mg/mL). The screening of inserts from transformants was performed by direct PCR amplification from colonies using the 16S primers 63F (CAG GCC TAA CAC ATG CAA GTC) and 1389R (ACG GGC GGT GTG TAC AAG). Heating at 94 °C for 10 min preceded standard cycling conditions (as above). Amplified inserts were grouped on the basis of restriction fragment length polymorphism (RFLP) patterns using a combination of the two restriction endonucleases CƒoI and AluI (Sigma) at 37 °C for 3 h. Representative clones of each RFLP group were sequenced, giving between 45 and 50 clones per sample. The insertions were sequenced by the Sanger method using primer 63 F and were performed at the Biotechnology Institute (Instituto de Biotecnología), National Autonomous University of Mexico (Universidad Nacional Autónoma de México, UNAM). The length of the obtained sequences was 600 to 700 nucleotides in average.
Phylogenetic analyses
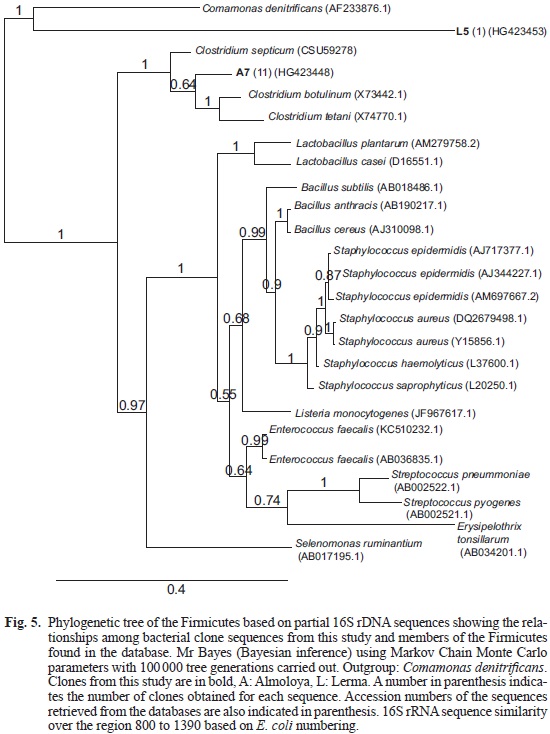
The sequences obtained were compared with the European Bioinformatics Institute and Genebank databases by online FastA searches (Pearson 1988) and the Check Chimera programme (Cole 2003) for detecting chimeras. Sequence alignments were carried out using MUSCLE (Edgar 2004) and alignment curation carried out using G-Block (Castresana 2000). Phylogenetic trees were constructed by Mr Bayes (Bayesian inference) using Markov Chain Monte Carlo parameters with 100 000 tree generations carried out. Tree sampling was performed after every 10 generations (Huelsenbeck 2001) with the final tree being viewed in TreeDyn (Chevenet et al. 2006, Dereeper et al. 2008, Dereeper et al. 2010)
RESULTS
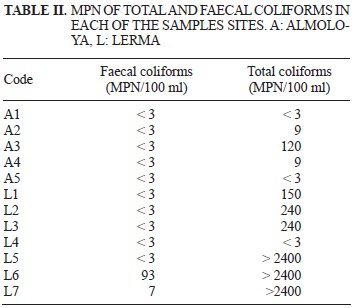
Total and faecal coliforms
Total and faecal coliforms determined with the MPN are shown in table II. Most of the sampling points (A1-A5 and L1-L4) total coliform values were below the limit (1000 MPN/100 ml) established by the Mexican Norm for Residual Waters to be reused in public services (NOM-003-SEMARNAT-1997) (SEMARNAT 1997). However, the total coliform values of sampling points L5, L6 and L7 in the Lerma wetland were significantly above (> 2400 MPN) the Norm limit.
Metals, semimetals and other elements
Concentrations for Ag, Al, As, Be, Cd, Co, Cr, Cu, Fe, Mn, Ni, Pb, Sb, Se, Tl, V and Zn were obtained for each of the sampling points within the Almoloya (A1-A5) and Lerma (L1-L7) wetlands (Table III). The concentrations of Ag, Cd, Sb, Se, As, Be and Tl were below the detection level for all sampling points (data not shown) except for A4 where 4.42, 1.88 and 14.6 mg/kg were detected for As, Be and Tl, respectively. Moreover, Tl was also detected in L7. The rest of the metal concentrations were above detection level in all sampling points except for Zn at a few locations (L3, L4, A1, A4; data not shown).
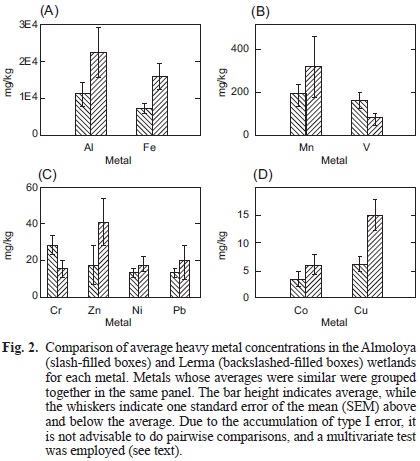
A statistical comparison of all element values between both wetlands revealed an overall similarity between them (p = 0.08) and a description of these values using means and standard error of means is displayed in figure 2. The overall similarity of the studied wetlands suggests that their influence area is substantially larger than both studied sites. The influence area includes wastewater discharges from industrial, agricultural and household sources.
Organic compounds
A total of 24 organic compounds were detected in the sediments of both wetlands and they include polyaromatic hydrocarbons, phthalates, fatty acids and higher alkanes (C15-C24) among others (Table IV). The higher alkanes were detected in all the sampling points (A1-A5, L1-L7), while sediments of sampling point L3 recorded the highest number of compounds with 12, followed by A1, L5 and L1 each with 11 and A3, A5, L2 and L4 each with nine compounds. Seven organic compounds were detected in A4 and six in L6 and L7. The detection quality of most of the compounds was above 95 % (Table IV).
Microbial diversity in situ
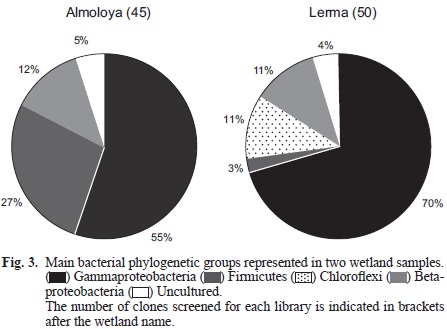
In order to identify the microbial diversity and their potential in the wetlands that give origin to the Lerma river, a clone library was created for each wetland: Almoloya (A) and Lerma (L). 16S rRNA gene PCR amplicons were obtained from the environmental samples using prokaryotic primers, and the main bacterial phylogenetic groups are summarized in figure 3. No archaeal amplicons were obtained. The Gammaproteobacteria followed by the Firmicutes dominates the Almoloya sample. Operational taxonomic units were formed on the basis of RFLP results and a total of 13 clones were 98 % similar to the uncultured Shigella sp. clone C199 (A4; Fig. 4). On the other hand 11 clones (A7; Fig. 5) were 95 % similar to those (affiliated to the Firmicutes) found in a study that analyzed the microbial composition of patients with inflammatory bowel diseases (IBD; Li et al. 2012). Other nine clones (A5; Fig. 4) were related to a Gammaproteobacteria isolated from soil contaminated with polyaromatic hydrocarbons (unpublished data). The rest of the clones (5 and 7 respectively) were similar to an uncultured Betaproteobacterium clone from epiphytic bacterial communities on native plant species in a midwestern Michigan creek (Olapade et al. 2011) and to other uncultured microorganisms from the bacterial community detected in different developmental stages of the housefly (Wei et al. 2013). The most abundant phylogenetic group of Bacteria in the Lerma wetland was also the Gammaproteobacteria (Fig. 3). Some of the organisms detected in the Lerma wetland were similar to uncultured microorganisms from diverse environments such as a lake in Panama (two clones), from a brewery wastewater treatment plant (three clones) and (as in Almoloya wetland) to plant-associated bacterial populations in freshwater from a Michigan creek (five clones; Olapade et al. 2011). Five other clones have close affiliation with an uncultured microorganism detected in a polychlorinated biphenyl (PCB) - contaminated core sediment from the Grasse River in New York undergoing dechlorination (Xu et al. 2012), while one was highly similar to another found in a plutonium contaminated soil (Kimber et al. 2012). Moreover, although two clones were similar to an E. coli isolate from healthy people gastric juices (Delgado et al. 2013) seven more were similar to uncultured bacteria detected in mammalian gut inflammation, and the majority of clones (20 clones) were related to a Shigella flexneri strain.
DISCUSSION
Metals, semimetals and other elements in sediments
The mean of the values detected in this study for As, Cr, Cu, Ni, Pb and Zn were below the ones obtained previously (Pérez-Ortiz 2005) in both wetlands except for Cr (Almoloya) suggesting a movement of the sediments due to the water flow. Also, these values are below the threshold of contamination as established by the USEPA (USEPA 1977) and permit aquatic life although with the lowest toxicity effect of Cr according to the Ontario Ministry of the Environment and Energy (OME 1976). In addition, these values are also below the ones reported for sediments in the Lerma River (Tejeda et al. 2006, Zarazúa et al. 2011) and the Alzate dam (Ávila-Pérez et al. 1999). This suggests that the high contamination reported in these studies was due to discharges from domestic and industrial activities along the river course (Ávila-Pérez et al. 1999). However, Al concentrations were the highest among all metals in both wetlands and they are significantly above the permissible limits (2700 mg/kg) of the National Oceanic and Atmospheric Administration of the United States (NOAA 1999). A recent study analyzed Al toxicity in zooplankton (rotifers, cladocerans and copepods) at the Lerma wetland suggesting that these organisms are good candidates for biological monitoring since they are more sensitive than other groups and have a high ecological relevance (García-García et al. 2012) . In that study only one species was frequently found where high concentrations (above 2350 mg/kg) of Al were recorded in sediments, what is consistent with previous studies reporting the negative effects of Al on zooplankton and fish populations (Havens and Heath 1989, Havens 1990, Gensemer and Playle 1999, Cherry et al. 2001). Moreover, we detected sediment concentrations of Al nearly three times higher than those detected from March 2008 to February 2009 (García-García et al. 2012) and almost ten times higher than the permissible limits (2700 mg/kg). This might be related to the increase of Al production and melting by the nearby industries in the recent years. Additionaly acid rain (consequence of industrial activities and burning of fossil fuels) makes the wetland waters more acidic and this, in turn, increases the solubility of Al (Moore 2006). Although a couple of values detected for Fe and Mn were above the USEPA limits (both at sampling point L7), their means were not only below those limits but also those reported for sediments along the upper course of the Lerma River (UCLR; Zarazúa et al. 2011). In addition, their source is presumed mainly natural as was established for the UCLR sediments (Zarazua et al. 2011). The mean of vanadium values for both wetlands are above the USEPA phytotoxic limit of 2 mg/kg for soil but within the range of what is considered typical in sediments (7-500 mg/kg) of the US irrespective of the parent material (Irwin et al. 1998). The highest values for all metals except V (2nd highest) were observed at sampling point L7 and this may be explained by the fact that it is next to the recently inaugurated (August 2011) highway Lerma-Tenango, suggesting its construction and usage has had an immediate impact on the wetlands.
The analysis of sediments is important because it is a direct indicator of the changes throughout time since the sediments absorb and accumulate metals that cannot be biodegraded (Rosales-Hoz et al. 2000). A portion of those metals can be transported vertically to the water column. Zooplankton is partially responsible for such transport since those organisms feed by filtration and incorporate metals from the sediments. This incorporation may lead to biological accumulation and bio magnification, potentially affecting humans through consumption of wetland-derived species such as fish, crayfish and axolotl (Ambystoma lermaense). Therefore, although the amounts of metals are in general below the limits permitted by the USEPA for lake sediments and significantly below the ones observed in the UCLR and the Alzate Dam sediments, it remains necessary to investigate the concentration of heavy metals and other toxic substances in biological tissues from all trophic levels in order to rule out biological magnification. Moreover, the introduction of selected species of macrophyte into the wetlands should be considered since it has been a viable solution to the treatment of highway runoff (Mungur et al. 1995).
Organic compounds detection
Among the identified organic compounds, benzenecarboxylic acid, naphthalene, 2,6-dimethyl-, 2,5-cyclohexadiene-1,4-dione, 2,6-bis (1,1-dimeth-ylethyl)-, diethyl phthalate, dibutyl phthalate, benzyl butyl phthalate, benzophenone, pentadecane, eico-sane, heptadecane, nonadecane, hexadecane and heneicosane have been listed as chemicals for potential endocrine disruptor screening and testing (USEPA 2012). Also 2,5-cyclohexadiene-1,4-dione, 2,6-bis (1,1-dimethylethyl)-, dibutyl phthalate, pentadecane, heptadecane, pentacosane and nonadecane have been found in samples of water with historical contamination and a related toxicity effect on fishes (Mancas. et al. 2002). Higher alkane hydrocarbons from C15 to C24, benzenecarboxylic acid and 2,5-cyclohexadiene-1,4-dione, 2,6-bis (1,1-dimethylethyl)- were found in all of the samples and some cycloalkanes (C14 and C15) were also frequently found. Higher alkane hydrocarbons are the main components of fuel and lubricating oils while benzenecarboxylic acid is used as a food preservative due to its inhibitory effects on the growth of mold, yeast and some bacteria and it is also an important precursor for the synthesis of many other organic substances. The 2,5-cyclohexadiene-1,4-dione, 2,6-bis (1,1-dimethylethyl)-, also known as 2,6-di-tert-butylbenzoquinone is considered a semi volatile priority organic pollutant of the steel finishing subcategory in the iron and steel manufacturing (USEPA 2002).
The group of phthalates compounds has also been reported as endocrine-disrupting chemicals. Dibutyl phthalate is the predominant phthalic acid ester present in agricultural soils and it can significantly affect paddy soil microbial diversity, regarding its population size and species representation (Zeng et al. 2008, Xie et al. 2013). Although the phthalates found in the samples are not classified as either mu-tagenic or carcinogenic, they are included in the list of priority pollutants by the USEPA (Kamrin 2009, Wang et al. 2013).
On the other hand, benzophenone related compounds are utilized as organic UV filters not only in cosmetics and personal care products but also in food packaging, pharmaceuticals, plastics, textiles and vehicle-maintenance products to prevent photodegradation of polymers and pigments (Cuquerella et al. 2012, Gago-Ferrero et al. 2012). These compounds are considered as emerging environmental pollutants because of their increasing use over the last decade related to the concern of skin damage due to UV-solar radiation. Despite being scattered and limited, current ecotoxicological data indicate that the potential risk posed by these widely used chemicals requires further investigation, particularly terrestrial environments should be more widely studied to identify their fate and effects (Gago-Ferrero et al. 2012). The presence of hydrocarbons in the sediments may be due to natural biological sources such as plant waxes or diagenetic transformation of functionalized lipids into the sediments (Volkman et al. 1992). However, we suggest that they are from anthropic activities since most of the detected hydrocarbons in the wetlands sediments are the main components of lubricating oils. This is supported by the fact that these type of oils are the major source of hydrocarbon pollution in many estuaries and coastal zones around Australia (Volkman et al. 1992) but more importantly, runoffs containing residues of these oils from the Lerma-Tenango highway may be increasing the hydrocarbon levels in the sediments.
Microbial diversity in situ
In general, the microbial diversity observed in both wetlands is low. This may be due to the ongoing environmental degradation in these water bodies.
The Almoloya wetland harbours microorganisms similar to an uncultured Shigella clone C199 and to an uncultured Firmicutes previously detected in patients with IBD. While uncultured bacteria found in mammalian gut inflammation (Stecher et al. 2012) and a strain of Shigella flexneri were the closest organisms to the clones retrieved from the Lerma wetland. Infection by Shigella is an important cause of morbidity and mortality around the world. An estimate from the World Health Organization indicates there are 164 million episodes of shigellosis per year, which resulted in 1.1 million deaths, mostly children under 5 years old (Kotloff et al. 1999). More importantly, the majority of the episodes and deaths occur in the developing world (163 million) and the main isolates correspond to S. flexneri, being the serotype 2a the most common (Kotloff et al. 1999). However in order to assess the pathogenicity of the Shigella-related clones detected in the wetlands, it is necessary to look for the genes that code for those toxins or virulence and invasion plasmids (Sansonetti et al. 1982, Menard et al. 1993, Tang et al. 2005) in future studies.
Dominance of clones related to Shigella and to bacteria found in gut studies (Stecher et al. 2012) in both wetlands is not uncommon since they receive domestic effluents from the surrounding towns and this is confirmed by the large number of coliforms > 2400 MPN 100 mL (Table II) recovered from a sampling point very near a sewage pipe from the San Pedro Tultepec town that discharges directly into the Lerma wetland (sampling point L6; Table I). The latter was also found in previous studies (Pérez-Ortiz 2005) and MPN values above the limit from nearby sampling points L5 and L7.
Although both wetlands were dominated by microorganisms partially related to Gammaproteobacteria, there were other clones related to those found in hydrocarbon, PCB and plutonium-contaminated environments. Several clones retrieved from the Almoloya sediments were related to a Gammaproteobacterium previously detected in a polyaromatic hydrocarbon-contaminated soil and several species of Gammaproteobacteria have been reported to degrade polyaromatic hydrocarbons (Boonchan et al. 1998, Ma et al. 2006, Arun et al. 2008). Also, clones retrieved from the Lerma sediments were closely related to those affiliated to the Chloroflexi and previously found in a PCB-contaminated core sediment from a New York river undergoing dechlorination (Xu et al. 2012). Members of the Chloroflexi have also been found in several hydrocarbon-degrading consortia in different environments such as soil (Aburto-Medina et al. 2012), a petroleum contaminated aquitard (Van Stempvoort et al. 2009), a BTEX contaminated aquifer (Berlendis et al. 2010) but more importantly in dimethyl phthalate and naphthalene-contaminated sludge (Liang et al. 2009, Cao et al. 2012); these hydrocarbons were also detected in the wetland sediments studied (Table IV). This may suggest the potential use of these available hydrocarbons by the microorganisms, however further laboratory experiments are needed in order to confirm such processes.
Another clone was similar to an uncultured bacterium found in a plutonium-contaminated soil in the United Kingdom (Kimber et al. 2012). Microbial processes performed by Clostridium have the potential to mobilize the plutonium by the reduction of Pu (IV) to Pu (III)(Francis et al. 2008, Kimber et al. 2012). This organism is also capable of performing hydrocarbon degradation (Sethuna-than and Yoshida 1973) and PCB dechlorination (Hou and Dutta 2000), suggesting a potential use of the available organic compounds detected in the wetland sediments by this species. However this bacterium is often found in soils and wastes and it is involved in the sulphur cycle, therefore there is also a need for further degradation experiments to validate those processes.
Despite the pressure on the marshes, contaminant levels are not extremely high in general suggesting that their self-cleansing system may still be functional. However, it is necessary to establish the origin of hydrocarbons and metals detected in the sediments in order to prevent an increase of their concentrations. Further experiments are required to rule out biomagnification of metals to protect both the local human population, who depends on the wetland and the endangered and endemic species, which live there.
ACKNOWLEDGMENTS
This research was funded by Universidad Autonoma Metropolitana-Lerma and the Consejo Nacional de Ciencia y Tecnología (CONACyT). The authors would also like to acknowledge the support of Claudia Granada M.Sc. for her help with the maps and Mr. Raúl Gutiérrez and Mr. Rufino Rodríguez Salas for giving access to the wetlands.
REFERENCES
Aburto A., Fahy A., Coulon F., Lethbridge G., Timmis K.N., Ball A.S. and McGenity T.J. (2009). Mixed aerobic and anaerobic microbial communities in benzene-contaminated groundwater. J. Appl. Microbiol. 106, 317-328. [ Links ]
Aburto-Medina A., Adetutu E., Aleer S., Weber J., Patil S., Sheppard P., Ball A. and Juhasz A. (2012). Comparison of indigenous and exogenous microbial populations during slurry phase biodegradation of long-term hydrocarbon-contaminated soil. Biodegradation 23, 813-822. doi:10.1007/s10532-012-9563-8. [ Links ]
Aguilar X. and Casas G. (2009). Anfibios y reptiles. En: Biodiversidad del Estado de México: Estudio de Estado (Secretaría del Medio Ambiente, Ed.). Gobierno del Estado de México y Comisión para el Conocimiento y Uso de la Biodiversidad. Toluca, Mexico, pp. 125-130. [ Links ]
Arun A., Raja P.P., Arthi R., Ananthi M., Kumar K.S. and Eyini M. (2008). Polycyclic Aromatic Hydrocarbons (PAHs) Biodegradation by Basidiomycetes Fungi, Pseudomonas isolate, and their cocultures: Comparative in vivo and in silico approach. Appl. Biochem. Biotechnol. 151, 132-142. doi:10.1007/s12010-008-8160-0. [ Links ]
Avila-Pérez P., Balcázar M., Zarazúa-Ortega G., Barceló-Quintal I. and Díaz-Delgado C. (1999). Heavy metal concentrations in water and bottom sediments of a Mexican reservoir. Sci. Total Environ. 234, 185-196. doi: http://dx.doi.org/10.1016/S0048-9697(99)00258-2. [ Links ]
Berlendis S., Lascourreges J.F., Schraauwers B., Sivadon P. and Magot M. (2010). Anaerobic biodegradation of BTEX by original bacterial communities from an underground gas storage aquifer. Environ. Sci. Technol. 44, 3621-3628. doi:10.1021/es100123b. [ Links ]
Boonchan S., Britz M.L. and Stanley G.A. (1998). Surfactant-enhanced biodegradation of high molecular weight polycyclic aromatic hydrocarbons by Stenotroph-omonas maltophilia. Biotechnol. Bioeng. 59, 482-494. [ Links ]
Cao X.K., Yang Q. and Hao C.B. (2012). Degradation kinetics of naphthalene by anaerobic sludge and analysis of the bacterial biodiversity. Chinese J. Environ. Sci. 33, 3535-3541. [ Links ]
Castresana J. (2000). Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 17, 540-52. [ Links ]
Cherry D.S., Currie R.J., Soucek D.J., Latimer H.A. and Trent G.C. (2001). An integrative assessment of a watershed impacted by abandoned mined land discharges. Environ. Pollut. 111, 377-388. doi: http://dx.doi.org/10.1016/S0269-7491(00)00093-2. [ Links ]
Chevenet F., Brun C., Banuls AL., Jacq B. and Chisten R. (2006). TreeDyn: towards dynamic graphics and annotations for analyses of trees. BMC Bioinformatics. 7, 439. [ Links ]
Cole J.R., Chai B., Marsh T.L., Farris R.J., Wang Q., Kulam S.A., Chandra S., McGarrell D.M., Schmidt T.M., Garrity G.M and Tiedje J.M. (2003). The Ribosomal Database Project (RDP-II): Previewing a new autoaligner that allows regular updates and the new prokariotic taxonomy. Nucleic Acids Res. 31, 442-443. [ Links ]
Colón C., Méndez Sánchez F. and Ceballos G. (2009). Peces. En: Biodiversidad del Estado de México: Estudio de Estado. (Secretaría del Medio Ambiente, Ed.). Gobierno del Estado de México y Comisión para el Conocimiento y Uso de la Biodiversidad. Toluca, Mexico, pp. 119-125. [ Links ]
Cuquerella M.C., Lhiaubet-Vallet V, Cadet J. and Miranda M.A. (2012). Benzophenone photosensitized DNA damage. Accounts Chem. Res. 45, 1558-1570. [ Links ]
Delgado S., Cabrera-Rubio R., Mira A., Suárez A. and Mayo B. (2013). Microbiological survey of the human gastric ecosystem using culturing and pyrosequencing methods. Microb. Ecol. 65, 753-772. doi:10.1007/s00248-013-0192-5. [ Links ]
Dereeper A., Audic S., Claverie J.M. and Blanc G. (2010). BLAST-EXPLORER helps you building datasets for phylogenetic analysis. BMC Evol. Biol. 10, 8. [ Links ] Dereeper A., Guignon V., Blanc G., Audic S., Buffet S., Chevenet F., Dufayard J.F., Guindon S., Lefort V., Lescot M., Claverie J.M. and Gascuel O. (2008). Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36, W465-469. [ Links ]
Edgar R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792-7. [ Links ]
Esteller M.V. and Díaz-Delgado C. (2002). Environmental effects of aquifer overexploitation: A case study in the highlands of Mexico. Environ. Manage. 29, 266-278. doi:10.1007/s00267-001-0024-0. [ Links ]
Francis A.J., Dodge C.J. and Gillow J.B. (2008). Reductive dissolution of Pu(IV) by Clostridium sp. under anaerobic conditions. Environ. Sci. Technol. 42, 2355-2360. [ Links ]
Gago-Ferrero P., Díaz-Cruz M.S. and Barceló D. (2012). An overview of UV-absorbing compounds (organic UV filters) in aquatic biota. Anal Bioanal. Chem. 404, 2597-2610. [ Links ]
García-García G., Nandini S., Sarma S.S.S., Martínez-Jerónimo F. and Jiménez-Contreras J. (2012). Impact of chromium and aluminium pollution on the diversity of zooplankton: A case study in the Chimaliapan wetland (Ramsar site) (Lerma basin, Mexico). J. Environ. Sci. Heal. Part A 47, 534-547. doi:10.1080/10934529.20 12.650554. [ Links ]
Gensemer R.W. and Playle R.C. (1999). The bioavailability and toxicity of aluminum in aquatic environments. Crit. Rev. Environ. Sci. Technol. 29, 315-450. doi:10.1080/10643389991259245. [ Links ]
Gómez-Salazar S., Domiguez-Mariani E., Solis-Correa H., López-Galvan E. and Barceló-Quintal I. (2012). Water quality assessment of José Antonio Alzate dam, the Lerma River and its tributaries in the State of Mexico, Mexico. J. Environ. Prot. 3, 878-888. [ Links ]
Havens K.E. (1990). Aluminum binding to ion exchange sites in acid-sensitive versus acid-tolerant cladocer-ans. Environ. Pollut. 64, 133-141. doi: http://dx.doi.org/10.1016/0269-7491(90)90110-X. [ Links ]
Havens K.E. and Heath R.T. (1989). Acid and aluminum effects on freshwater zooplankton: An in situ meso-cosm study. Environ. Pollut. 62, 195-211. doi: http://dx.doi.org/10.1016/0269-7491(89)90187-5. [ Links ]
Hou L.H. and Dutta S.K. (2000). Phylogenetic characterization of several para- and meta-PCB dechlorinating Clostridium species: 16s rDNA sequence analyses. Lett. Appl. Microbiol. 30, 238-243. doi:10.1046/j.1472-765x.2000.00709.x. [ Links ]
Huelsenbeck JP. and Ronquist F. (2001). MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17, 754-5. [ Links ]
Irwin R.J., VanMouwerik M., Stevens L., Seese M.D. and Basham W. (1998). Environmental Contaminants Encyclopedia. National Park Service, Water Resources Division, Fort Collins, Colorado [online]. http://www.nature.nps.gov/water/ecencyclopedia/index.cfm. 7/10/2014. [ Links ]
Kamrin M.A. (2009). Phthalate risks, phthalate regulation, and public health: a review. J. Toxicol. Env. Heal. B 12, 157-174. [ Links ]
Kimber R.L., Boothman C., Purdie P., Livens F.R. and Lloyd J.R. (2012). Biogeochemical behaviour of plutonium during anoxic biostimulation of contaminated sediments. Mineral Mag. 76, 567-578. doi:10.1180/minmag.2012.076.3.08. [ Links ]
Kotloff K., Winickoff J., Ivanoff B., Clemens J.D., Swerd-low D., Sansonetti P., Adak G. and Levine M. (1999). Global burden of Shigella infections: implications for vaccine development and implementation of control strategies. B. World Health. Organ. 77, 651-666. [ Links ]
Lane D.J. (1991). 16S/23S rRNA sequencing. En Nucleic Acid Techniques in Bacterial Systematics (E. Stacke-brandtand M. GoodfellowEds.). John Wiley and Sons. Chichester, England, pp. 115-175. [ Links ]
Li E., Hamm C.M., Gulati A.S., Sartor R.B., Chen H., Wu X., Zhang T., Rohlf F.J., Zhu W., Gu C., Robertson C.E., Pace N.R., Boedeker E.C., Harpaz N., Yuan J., Weinstock G.M., Sodergren E. and Frank D.N. (2012). Inflammatory bowel diseases phenotype, C. Difficile and NOD2 genotype are associated with shifts in human ileum associated microbial composition. PLoS ONE 7, e26284. doi:10.1371/journal.pone.0026284. [ Links ]
Liang D.W., Fang H.H.P. and Zhang T. (2009). Microbial characterization and quantification of an anaerobic sludge degrading dimethyl phthalate. J. Appl. Microbiol. 106, 296-305. doi:10.1111/j.1365-2672.2008.04003.x. [ Links ]
López-Galván E., Barceló-Quintal I., Solís-Correa H., Bussy A., Ávila-Pérez P. and Delgadillo S. (2010). Calculation of the Ecological Risk Index in the José Antonio Alzate Dam, State of Mexico, Mexico. Biol. Trace Elem. Res. 135, 121-135. doi:10.1007/s12011-009-8501-z. [ Links ]
Lyons J., Navarro-Pérez S., Cochran P.A., Santana E.C. and Guzmán-Arroyo M. (1995). Index of biotic integrity based on fish assemblages for the conservation of streams and rivers in west-central Mexico. Conserv. Biol. 9, 569-584. doi:10.1046/j.1523-1739.1995.09030569.x. [ Links ]
Ma Y., Wang L. and Shao Z. (2006). Pseudomonas, the dominant polycyclic aromatic hydrocarbon-degrading bacteria isolated from Antarctic soils and the role of large plasmids in horizontal gene transfer. Environ. Microbiol. 8, 455-465. doi:10.1111/j.1462-2920.2005.00911.x. [ Links ]
Mancas G., Vartiainen T., Rantakokko P., Navrotescu T., Diaconu R., Mancas D. and Diaconu D. (2002). Chemical contaminants in drinking water: Mutagenic and toxic effects. Am. J. Prev. Med. 10, 63-74. [ Links ]
Marchesi J.R., Sato T., Weightman A.J., Martin T.A., Fry J.C., Hiom S.J. and Wade W.G. (1998). Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 64, 795-799. [ Links ]
Ménard R., Sansonetti P.J. and Parsot C. (1993). Nonpolar mutagenesis of the ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigella flexneri entry into epithelial cells. J. Bacteriol. 175, 5899-5906. [ Links ]
Moore P.D. and Garratt R. (2006). Biomas of the earth-wetlands. Chelsea House Publishers. New York, USA, 144 pp. [ Links ]
Mungur A.S., Shutes R.B.E., Revitt D.M. and House M.A. (1995). An assessment of metal removal from highway runoff by a natural wetland. Water Sci. Tech-nol. 32, 169-175. doi: http://dx.doi.org/10.1016/0273-1223(95)00617-6. [ Links ]
NOAA (1999). Coastal protection and restoration division. Report. National Oceanic and Atmospheric Administration HAZMAT. US Department of Commerce. Seattle, WA. USA, 12 pp. [ Links ]
Olapade O.A., Pung K., Goldman E. and Lyons-Sobaski S. (2011). Occurrence and diversity of epiphytic bacterial communities on two native plant species in a Michigan Creek. J. Freshw. Ecol. 26, 267-276. doi:10.1080/027 05060.2011.555214. [ Links ]
OME (1976). Evaluating construction activities impacting on water resources part III A. Handbook for dredging and dredged material disposal in Ontario. Legislation policies, sediment classification and disposal options. Ontario Ministry of the Environment and Energy. Guideline, Toronto, Canada, 49 pp. [ Links ]
Pearson W.R. and Lipman D.J. (1988). Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85, 2444-2448. [ Links ]
Pérez-Ortiz G. (2005). Diagnóstico ambiental como base para la rehabilitación de las Ciénegas del Lerma, Estado de México. Tesis de Maestría en Ciencias Biológicas. Universidad Nacional Autónoma de México. Mexico, 92 pp. [ Links ]
Pérez-Ortiz G. y Valdez M. (2006). El uso de la biodivers-idad en las ciénegas del Lerma. En: Atlas de la Cuenca Chapala. (A.H. Cotler, M. Mazari y J. de A. Sánchez, Eds.). Instituto Nacional de Ecología y Secretaría del Medio Ambiente y Recursos Naturales. Mexico, pp. 78. [ Links ]
Ramos VL. (1999). Estudio de la flora y la vegetación acuática vasculares de la Cuenca alta del Río Lerma, en el Estado de México. Tesis de Maestría. Facultad de Ciencias, Universidad Nacional Autónoma de México, Mexico, 151 pp. [ Links ]
Rencher A.C. (2002). Methods of multivariate analysis. John Wiley and Sons. New York, USA, 627 pp. [ Links ]
Rosales-Hoz L., Carranza-Edwards A. and López-Hernández M. (2000). Heavy metals in sediments of a large, turbid tropical lake affected by anthropogenic discharges. Environ. Geol. 39, 378-383. doi:10.1007/s002540050017. [ Links ]
Sansonetti P.J., Kopecko D.J. and Formal S.B. (1982). Involvement of a plasmid in the invasive ability of Shigella flexneri. Infect. Immun. 35, 852-860. [ Links ]
Segura-García V., Cantoral-Uriza E.A., Israde I. and Maidana N. (2012). Epilithic diatoms (Bacillariophyceae) as indicators of water quality in the upper Lerma river, Mexico. Hidrobiológica 22, 16-27. [ Links ]
SEMARNAT (1997). Norma Oficial Mexicana NOM-003-SEMARNAT-1997. Que establece los límites máximos permisibles de contaminantes para las aguas residuales tratadas que se reusen en servicios al público. Diario Oficial de la Federación. Mexico, 6 de enero de 1997. [ Links ]
SEMARNAT (2002). Decreto por el cual se declara Área natural protegida, con el carácter de área de protección de flora y fauna, la región conocida como Ciénegas del Lerma. Diario Oficial de la Federación. Mexico, 27 de noviembre de 2002. [ Links ]
Sethunathan N. and Yoshida T. (1973). Degradation of chlorinated hydrocarbons by Clostridium sp. isolated from lindane-amended, flooded soil. Plant and Soil 38, 663-666. doi:10.1007/bf00010705. [ Links ]
Stecher B., Denzler R., Maier L., Bernet F., Sanders M.J., Pickard D.J., Barthel M., Westendorf A.M., Krogfelt K.A., Walker A.W., Ackermann M., Dobrindt U., Thomson N.R. and Hardt W.D. (2012). Gut inflammation can boost horizontal gene transfer between pathogenic and commensal Enterobacteriaceae. Proc. Natl. Acad. Sci. USA 109, 1269-1274. doi:10.1073/pnas.1113246109. [ Links ]
Tejeda S., Zarazúa-Ortega G., Ávila-Pérez P., García-Mejía A., Carapia-Morales L. and Díaz-Delgado C. (2006). Major and trace elements in sediments of the upper course of Lerma River. J. Radioanal. Nucl. Chem. 270, 9-14. doi:10.1007/s10967-006-0342-z. [ Links ]
USEPA (1977). Guidelines for pollution classification of Great Lakes Harbor sediments. United States Environmental Protection Agency Region V, Great Lakes Surveillance Branch. Chicago, IL. USA, 8 pp. [ Links ]
USEPA (2002). Development Document for Final Effluent Limitations Guidelines and Standards for the Iron and Steel Manufacturing Point Source Category. United States Environmental Protection Agency EPA-821-R-02-004 [online]. water.epa.gov/scitech/wastetech/guide/ironsteel/upload/2002_ 10_17_guide_ironsteel_reg_tdd_complete.pdf 7/10/2014. [ Links ]
USEPA (2007a). Method 6010C Inductively Coupled Plasma-Atomic Emission Spectrometry. United States Environmental Protection Agency. Test Methods for Evaluating Solid Waste, Physical/Chemical Methods. Washington, DC. USA, 34 pp. [ Links ]
USEPA (2007b). Method 3051A Microwave Assisted Acid Digestion of sediments, sludges, soils and oils. United States Environmental Protection Agency Test Methods for Evaluating Solid Waste, Physical/Chemical Methods. Washington, DC. USA, 30 pp. [ Links ]
USEPA (2007c). Ultrasonic Extraction, Method 3550C, United States Environmental Protection Agency. Washington, DC. USA, 17 pp. [ Links ]
USEPA (2007d). Semivolatile Organic Compounds, Method 8270D, United States Environmental Protection Agency. Washington, DC. USA, 72 pp. [ Links ]
USEPA (2012). Endocrine Disruptor Screening Program. Universe of Chemicals. United States Environmental Protection Agency [online]. http://www.epa.gov/endo/pubs/edsp_chemical_universe_list_ 11_12.pdf 7/10/2014. [ Links ]
Van Stempvoort D.R., Millar K. and Lawrence J.R. (2009). Accumulation of short-chain fatty acids in an aquitard linked to anaerobic biodegradation of petroleum hydrocarbons. Appl. Geochem. 24, 77-85. doi:10.1016/j.apgeochem.2008.11.004. [ Links ]
Vázquez R.H. (2004). Preferencias de hábitat por la avifauna presente en la laguna Chimaliapan, Ciénegas de Lerma, Lerma Estado de México. Tesis de Maestría, Facultad de Estudios Superiores Iztacala, UNAM. Mexico, 131 pp. [ Links ]
Volkman J.K., Holdsworth D.G., Neill G.P. and Bavor Jr. H.J. (1992). Identification of natural, anthropogenic and petroleum hydrocarbons in aquatic sediments. Sci. Total Environ. 112, 203-219. doi: http://dx.doi.org/10.1016/0048-9697(92)90188-X. [ Links ]
Wang X., Lina Q., Wang J., Lua X. and Wang G. (2013). Effect of wetland reclamation and tillage conversion on accumulation and distribution of phthalate esters residues in soils. Ecol. Eng. 51, 10-15. [ Links ]
Wei T., Hu J., Miyanaga K. and Tanji Y. (2013). Comparative analysis of bacterial community and antibiotic-resistant strains in different developmental stages of the housefly (Musca domestica). Appl. Microbiol. Biot. 97, 1775-1783. doi:10.1007/s00253-012-4024-1. [ Links ]
Xie K., Yang S., Xu P., Zhang H., Tang S., Zhang F., Huang X. and Gu W. (2013). Contamination of paddy soil by endocrine-disrupting chemicals affects soil microbe abundance and diversity. Soil Sediment Contam. An. Int. J. 22, 510-520. doi:10.1080/15320383.2013.750265. [ Links ]
Xu Y., Yu Y., Gregory K. and VanBriesen J. (2012). Comprehensive assessment of bacterial communities and analysis of PCB congeners in PCB-contaminated sediment with depth. J. Environ. Eng. 138, 1167-1178.doi:10.1061/(ASCE)EE.1943-7870.0000595. [ Links ]
Yang F., Yang J., Zhang X., Chen L., Jiang Y., Yan Y., Tang X., Wang J., Xiong Z., Dong J., Xue Y., Zhu Y., Xu X., Sun L., Chen S., Nie H., Peng J., Xu J., Wang Y., Yuan Z., Wen Y., Yao Z., Shen Y., Qiang B., Hou Y., Yu J. and Jin Q. (2005). Genome dynamics and diversity of Shigella species, the etiologic agents of bacillary dysentery. Nucleic Acids Res. 33, 6445-6458. doi:10.1093/nar/gki954. [ Links ]
Zarazúa G., Tejeda S., Ávila-Pérez P., Carapia L., Carreño C. and Balcázar M. (2011). Metal content and elemental composition of particles in cohesive sediments of the Lerma River, Mexico. Rev. Int. Contam. Ambie. 27, 181-190. [ Links ]
Zeng F., Cui K., Xie Z., Wu L., Liu M., Sun G., Lin Y., Luo D. and Zeng Z. (2008). Phthalate esters (PAEs): Emerging organic contaminants in agricultural soils in peri-urban areas around Guangzhou, China. Environ. Pollut. 156, 425-434. doi: http://dx.doi.org/10.1016/j.envpol.2008.01.045. [ Links ]
Zepeda-Gómez C, Lot-Helgueras A., Nemiga X.A. and Madrigal-Uribe D. (2012). Florística y diversidad de las ciénegas del río Lerma Estado de México, México. Acta Bot. Mex. 98, 23-49. [ Links ]

