










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista internacional de contaminación ambiental
versión impresa ISSN 0188-4999
Rev. Int. Contam. Ambient vol.29 no.1 Ciudad de México feb. 2013
Revisiones
Síntesis y biodegradación de polihidroxialcanoatos: plásticos de origen microbiano
Synthesis and biodegradation of polyhydroxialkanoates: Bacterially produced plastics
Yolanda GONZÁLEZ GARCÍA, Juan Carlos MEZA CONTRERAS, Orfil GONZÁLEZ REYNOSO y Jesús Antonio CÓRDOVA LÓPEZ
Departamento de Madera, Celulosa y Papel, Universidad de Guadalajara
Recibido diciembre 2011,
Aceptado noviembre 2012
RESUMEN
Los polihidroxialcanoatos (PHA) son biopoliésteres sintetizados intracelularmente por algunos microorganismos como reserva de carbono y energía que, una vez extraídos de la célula, presentan propiedades físicas similares a plásticos derivados del petróleo. A partir de la década de 1980 han sido estudiados intensivamente y actualmente siguen siendo un tema de investigación importante, sobre todo como sustitutos de los plásticos de origen petroquímico, ya que los PHA son completamente biodegradables y se producen a partir de fuentes de carbono renovables. También tienen una aplicación importante como materiales biocompatibles en el área biomédica y farmacéutica. En este trabajo se presenta una revisión bibliográfica extensa sobre las investigaciones realizadas respecto a la biosíntesis y la biodegradación de los PHA, así como el estado actual de su producción y comercialización a gran escala, y las perspectivas futuras en relación al estudio y aplicación de estos bioplásticos.
Palabras clave: Polihidroxialcanoatos, polihidroxibutirato, despolimerasas, biopoliésteres.
ABSTRACT
Polyhydroxyalkanoates (PHA) are biopolyesters synthesized by numerous microorganisms as an intracellular carbon and energy storage compound, and they have physical and chemical properties similar to those of petroleum-based plastics. They have been extensively studied since 1980, nowadays they are still an important research topic as substitutes of conventional plastics since PHA are biodegradable and synthesized from renewable resources. They also have an important application as biocompatible materials in the biomedical and pharmaceutical field. In this work is presented an extensive bibliographic review about scientific research on the biosynthesis and biodegradation of PHA, the status of its industrial production and commercialization, and the future perspectives regarding the study and application of these bioplastics.
Key words: Polyhydroxyalkanoates, polyhydroxybutyrate, depolymerase, biopolyesters.
INTRODUCCIÓN
Los productos plásticos tienen cualidades muy versátiles entre las que destacan su durabilidad y resistencia a la degradación, por lo que tienen un uso extendendido en la manufactura de una amplia variedad de productos, los cuales resultan imprescindibles para el estilo de vida actual (Khanna y Srivastava 2005). Estos productos son también parte esencial de todas las industrias y en muchos casos han reemplazado el uso de vidrio y papel como materiales de empaque. Su consumo anual se ha incrementado de 5 millones de toneladas, en 1950, a cerca de 200 millones de toneladas al año actualmente. Sin embargo, al ser resistentes a la degradación, su uso extensivo genera desechos que se acumulan en el ambiente a una tasa de 25 millones de toneladas al año, 40 % de las cuales son dispuestas en rellenos sanitarios, mientras que cientos de miles de toneladas son arrojadas a ambientes marinos. Este hecho ocasiona graves problemas, entre los que destacan el requerimiento de grandes espacios para su disposición, contaminación visual y muerte de animales que los ingieren accidentalmente, entre otros (Reddy et al. 2003, Salehizadeh y Van Loosdrecht 2004).
Se ha intentado atacar la problemática de la acumulación de plásticos en el ambiente a través de procesos de incineración, reciclaje, fotodegradación o reuso. Sin embargo estas medidas presentan desventajas por lo que no se consideran como soluciones 100% efectivas. Por ejemplo, durante el proceso de incineración se puede desprender ácido cianhídrico y ácido clorhídrico con potenciales daños para la salud. En el caso del reciclaje, este es complicado y consume mucho tiempo, además de que la presencia de muchos aditivos en los plásticos, como pigmentos y cubiertas, limita el proceso (Khanna y Srivastava 2005).
En vista de esta situación, la síntesis y uso de plásticos biodegradables en lugar de los derivados del petróleo se presenta como una solución efectiva. Actualmente ya existe la opción de utilizar un tipo de bioplásticos producidos por fermentación, llamados de manera genérica polihidroxialcanoatos (PHA), y particularmente el polihidroxibutirato (P3HB) y el polihidroxibutirato-cohidroxivalerato P(3HBco3HV), que se producen a escala comercial. Sin embargo el precio de los PHA (2 a 5 euros/ kg) es todavía alto en comparación con el de los plásticos derivados del petróleo, por lo que se están tomando medidas para hacerlos más accesibles a la población, entre las cuales figuran mejoras en el proceso de fermentación y extracción, así como aislar y desarrollar cepas microbianas más productivas y que puedan utilizar sustratos de bajo costo (Khanna y Srivastava 2005).
La investigación sobre la producción de PHA ha dado lugar a la obtención de cultivos con una concentración de polímero de más de 80 g/L y con una productividad de más de 2 g/L/h, usando sistemas por lote alimentado o continuo que potencialmente podrían producir 50 000 toneladas por año (Salehizadeh y Van Loosdrecht 2004).
Además de los problemas por disponibilidad de espacio y contaminación asociada a la producción y desecho de plásticos sintetizados químicamente, otra cuestión importante es la disponibilidad y el precio del petróleo, que es la materia prima para la producción de plásticos convencionales. Actualmente se utiliza un 5 % del petróleo disponible mundialmente para fabricar 200 millones de toneladas de plásticos por año. Sin embargo para el año 2100 se estima que la demanda de productos plásticos será de 2000 millones toneladas anuales, para lo cual será requerido usar el 50 % del petróleo disponible en ese momento. Tal situación tendrá un impacto en el precio de los plásticos, que se pronostica se incrementará drásticamente.
Considerando la problemática planteada anteriormente, la síntesis y uso de plásticos de origen microbiano podría ser una alternativa sustentable, ya que se producen a partir de fuentes de carbono renovables y tienen la ventaja de ser completamente biodegradables, es decir, mineralizados por la acción de microorganismos. Aunado a este beneficio ambiental, actualmente el uso de bioplásticos como materiales en el área médica y farmacéutica es de gran interés, por lo que se está trabajando intensamente en investigaciones científicas y tecnológicas para estas aplicaciones.
En esta investigación se presenta una revisión de los principales estudios que se han hecho alrededor del mundo, en relación con la biosíntesis y degradación de polihidroxialcanoatos, que son plásticos biodegradables de origen microbiano, similares a los derivados del petróleo pero producidos a partir de fuentes de carbono renovables, y que tienen un gran potencial de uso en aplicaciones que van desde la manufactura de productos desechables de uso común, hasta la de productos biomédicos y farmacéuticos de alto valor agregado. Se concluye con un análisis sobre el futuro de las investigaciones sobre este tema así como las aportaciones científicas hechas por investigadores latinoamericanos y las áreas de oportunidad para el estudio de estos biopoliésteres en México.
Definición, estructura y propiedades de los polihidroxialcanoatos
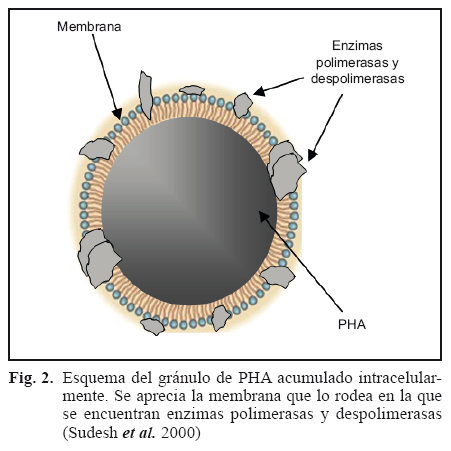
Los PHA son polímeros de ácidos hidroxialcanoicos que algunas bacterias, arqueas y microalgas acumulan intracelularmente como material de reserva, para usarlo posteriormente como fuente de carbono y energía. La estructura general y nomenclatura de estos compuestos se muestra en la figura 1. La polimerización de los ácidos hidroxialcanoicos, por acción de enzimas intracelulares, tiene lugar mediante condensación del grupo carboxilo de un monómero (ácido hidroxialcanoico), con el grupo hidroxilo del siguiente, formándose un enlace éster de allí que también se les conozca como biopoliésteres (Khanna y Srivastava 2005). Se acumulan como polímeros líquidos, móviles y amorfos en forma de gránulos que se alojan en el citoplasma microbiano rodeados de una monocapa de fosfolípidos que contiene enzimas polimerasas y despolimerasas (Fig. 2). Las investigaciones sobre el proceso de acumulación de PHA indican que el número de gránulos por célula se define en las primeras etapas de acumulación y que la producción del polímero cesa cuando su contenido alcanza cerca del 80 % del peso celular en base seca. Este fenómeno ha llevado a la conclusión de que existen restricciones físicas que impiden a la célula acumular más polímero, a pesar de la disponibilidad de sustrato y actividad de la enzima PHA polimerasa (Wang y Lee 1997). Estas inclusiones se observan bajo el microscopio como gránulos esféricos de diferentes tamaños (Braunegg et al. 1998) tal como se muestra en la figura 3.


Como se mencionó anteriormente, los PHA tienen características físicas similares a las de los plásticos derivados del petróleo, como el polipropileno y polietileno, pero tienen la ventaja de que pueden ser sintetizados a partir de fuentes de carbono renovables, son biodegradables (pueden ser asimilados por muchos microorganismos ya sea de suelos, mares, lagos o aguas residuales) y son biocompatibles (no causan efectos tóxicos). Estas propiedades les confieren una gran importancia como substitutos de los plásticos convencionales (Anderson y Dawes 1990).
En relación con las condiciones bajo las cuales ocurre la síntesis de los PHA, en la mayoría de los casos tiene lugar en respuesta a una limitación de N, P, S, Mg u oxígeno (Reddy et al. 2003), aunado a la presencia de un exceso de fuente de carbono en el medio de cultivo. Sin embargo, también existen bacterias que presentan una producción del polímero asociada al crecimiento, aunque este caso es menos frecuente. En el cuadro I se muestran los principales nutrientes cuya limitación da lugar a la síntesis de PHA en diferentes bacterias.

Respecto a su estructura monomérica, los PHA se pueden clasificar en varios tipos según la longitud de la cadena de los ácidos hidroxialcanoicos de los que están constituidos (Fig. 4). De esta manera, se definen como PHA de cadena corta (PHA-scl) aquellos polímeros compuestos por unidades monoméricas de 3 a 5 átomos de carbono, y como PHA de cadena media (PHA-mcl) a los que se componen por monómeros de 6 a 14 átomos de carbono. Sin embargo, también existen PHA mixtos, es decir que se componen de monómeros tanto de cadena corta como de cadena media. El tipo de PHA sintetizado depende del microorganismo en cuestión, por lo que la mayoría sólo produce PHA de cadena corta ó PHA de cadena media, y sólo una pequeña parte de ellos es capaz de producir PHA mixtos. Asimismo, los PHA se pueden presentar como homopolímeros o copolímeros, dependiendo también del tipo de microorganismo que se trate y las fuentes de carbono usados (Anderson y Dawes 1990). Hasta el momento se han descubierto más de 100 diferentes unidades monoméricas como constituyentes de los PHA (Lee 1996). En el cuadro II se muestra un ejemplo de la variedad de monómeros que se pueden presentar en un PHA. Es importante mencionar que dependiendo de su composición monomérica, el biopoliéster presentará diferentes propiedades físicas, las que se mencionarán posteriormente en este documento (Babel y Steinbüchel 2001).

Respecto a la diversidad de microorganismos productores de PHA, fuentes de carbono empleadas y porcentajes de acumulación, se han reportado más de 300 especies, principalmente bacterias, que utilizan sustratos que van desde carbohidratos, lípidos y proteínas, hasta compuesto aromáticos, residuos agroindustriales y gases, acumulando desde 1 hasta 80 % de polímero en relación al peso de su biomasa en base seca.
Desde que los PHA se descubrieron por primera vez , los investigadores de este tema se han referido a ellos como compuestos de naturaleza lipídica, y han dejado en claro que estos materiales son poliésteres insolubles en agua. La mayoría de los PHA son polímeros parcialmente cristalinos y por lo tanto sus propiedades térmicas y mecánicas usualmente se representan en términos de la temperatura de transición (Tg) o de la temperatura de fusión (Tm) (Anderson y Dawes 1990). Estos valores se presentan en el cuadro III para diferentes tipos de PHA.
El P3HB es una hélice levógira compacta que es ópticamente activa con el centro quiral de la unidad monomérica en la configuración R, esta estructura es muy similar al polipropileno. Las propiedades mecánicas como módulo de Young y la fuerza tensil de estos biopolímeros también están cercanas a las del polipropileno (Anderson y Dawes 1990).
Los PHA de cadena corta son típicamente polímeros termoplásticos, que pueden ser moldeables arriba de sus puntos de fusión. La temperatura de fusión es relativamente alta (180 °C) y su temperatura de transición está entre -5 y 20 °C. Este tipo de PHA comúnmente exhibe un grado de cristalinidad de 60 a 80 % el cual disminuye a 30 y 40 % a medida que el contenido de 3HV incrementa a 30 mol %. La incorporación de 3HV en el polímero también provoca que tanto la Tm como la Tg disminuyan significativamente, por lo que los copolímeros de cadena corta son materiales más versátiles que el homopolímero (Anderson y Dawes 1990).
En cuanto a los PHA de cadena media, estos son altamente amorfos con una Tg de entre -62 y -26 °C y Tm de 42 a 58 °C, por lo cual se clasifican como elastómeros.
Con respecto al peso molecular de los PHA, éste depende de las condiciones de producción y recuperación de estos compuestos. Los mecanismos que afectan y determinan el peso molecular de los PHA en las células bacterianas aún no son completamente entendidos, pero generalmente se atribuyen principalmente al tipo de microorganismo y la fuente de carbono usada (Anderson y Dawes 1990).
Por su parte, el método de recuperación del polímero puede influenciar el peso molecular significativamente. La extracción con solventes orgánicos da lugar a polímeros con peso molecular más alto, comparado con la extracción a base de hipoclorito de sodio u otros químicos. Debido a esto, existen PHA con una gran variedad de pesos moleculares, tal como se observa en el cuadro IV (Babel y Steinbüchel 2001). En general el peso molecular promedio de P3HB producido por bacterias silvestres está en el rango de 1 x 104 a 3 x 106 con una polidispersidad en el rango de 1.8 a 2.7.

Historia sobre la investigación de los PHA
La observación al microscopio de gránulos refráctiles en células bacterianas se remonta al año 1888. Sin embargo, la determinación de la composición de estos gránulos tuvo que esperar hasta 1925, como resultado de los estudios de Lemoigne en la bacteria Bacillus megaterium. En su investigación observó que al degradarse, este material desconocido liberaba ácido 3-hidroxibutírico y posteriormente describió dicho material como un homopoliéster de 3-hidroxibutirato, o poli-3-hidroxibutirato (P3HB), catalogándolo como un material de reserva (Braunegg et al. 1998).
Durante las siguientes tres décadas, el interés en el P3HB fue escaso y casi sólo se hicieron investigaciones con bacterias del género Bacillus. La propuesta de que el P3HB tenía un papel funcional se hizo en el año de 1958 por Macrae y Wilkinson. Estos científicos observaron que B. megaterium acumulaba el homopolímero cuando la relación carbono nitrógeno del medio era alta, y que la degradación del mismo ocurría rápidamente en la ausencia de una fuente de carbono y energía exógena. Concluyeron que el P3HB era un material de reserva de carbono y energía que retardaba la autolisis y muerte de las células (Braunegg et al. 1998).
En los siguientes quince años tuvo lugar un proceso de investigación extensa sobre la producción de este biopolímero en microorganismos fuera del género Bacillus incluyendo a Pseudomonas, Azotobacter, Hydrogenomonas, Chromatium, Cyanobacterium y muchas otras. También se iniciaron estudios sobre las propiedades químicas y físicas del P3HB, como peso molecular, temperatura de fusión, cristalinidad, morfología del gránulo, así como métodos de extracción, métodos de identificación y cuantificación, metabolismo, enzimología, degradación y función fisiológica (Babel y Steinbüchel 2001). En esta etapa fue de particular interés encontrar la relación entre la biosíntesis de P3HB y el ambiente extracelular. Las condiciones que favorecen la síntesis de P3HB fueron identificadas como aquellas que daban lugar a altas concentraciones de NAD(P)H y acetil-CoA, y baja concentración de coenzima-A libre. Se observó que estas condiciones variaban según el microorganismo, pero la biosíntesis del polímero siempre involucraba un factor de limitación de crecimiento como nitrógeno, fósforo, azufre, oxígeno o magnesio (Anderson y Dawes 1990).
El potencial del P3HB para ser usado en aplicaciones industriales fue reconocido por primera vez en la primera mitad de la década de 1960, cuando surgieron las primeras patentes relacionadas con su producción a través de procesos de fermentación, extracción, plastificación y mezclado con otros materiales. Sin embargo los plásticos derivados del petróleo eran mucho más baratos en ese momento y no había razón para pensar que los combustibles fósiles no fueran a permanecer a bajo precio, además de que las políticas de protección ambiental no estaban fuertemente establecidas en esa época (Braunegg et al. 1998).
La trigésima quinta conferencia de la organización de países exportadores de petróleo (OPEC) en Viena en septiembre de 1973, comenzó una cadena de eventos que cambiarían la situación con respecto a la investigación sobre producción de plásticos alternos. Los miembros de la OPEC decidieron incrementar los precios del petróleo en un 70 %. Este incremento fue seguido por otro más, anunciado en la conferencia de Teherán en diciembre del mismo año, en esa ocasión en un 130 %. En los años subsiguientes se dieron nuevos incrementos lo cual dio lugar a la conclusión de que las industrias de polímeros basados en petróleo no podrían sostenerse en un futuro, por lo que comenzó una intensa investigación para encontrar materiales plásticos alternos (Braunegg et al. 1998).
En 1976 la empresa Imperial Chemical Industries (ICI) de Inglaterra comenzó a investigar si el P3HB podría ser producido de manera económica por fermentación bacteriana a partir de carbohidratos procedentes de la agricultura. Como resultado de estos estudios, las propiedades de biodegradabilidad y biocompatibilidad de este biopolímero, así como la posibilidad de usarlo como fuente de moléculas ópticamente activas, fueron reconocidas como características de valor agregado. Estos resultados hicieron que ICI mantuviera el interés en la producción de P3HB aún después de terminada la crisis petrolera (Babel y Steinbüchel 2001).
En la década de 1980, Wallen y Davis reportaron por primera vez el aislamiento de un biopoliéster con propiedades físicas y químicas similares al P3HB pero con una composición diferente. Más tarde los análisis revelaron la presencia de 3-hidroxivalerato (3HV), 3-hidroxibutirato, 3-hidroxihexanoato (3HHx) y 3-hidroxiheptanoato (3HHp). Este fue el primer reporte de un heteropolímero, y debido a la diversidad de ácidos hidroxialcanoicos constituyentes estos biopolímeros (incluyendo el P3HB) comenzaron a llamarse de manera general polihidro-xialcanoatos (PHA). Muchos otros descubrimientos se hicieron en la década de 1980 en relación con la diversidad de monómeros constituyentes de los PHA, e ICI registró varias patentes para la producción, extracción y procesado de estos nuevos heteropolímeros (Braunegg et al. 1998).
Actualmente los PHA ya se producen comercialmente. Entre los más importantes está un copolímero de 3HB y 3HV, que tiene un porcentaje de 1 a 24 mol % de 3HV. Éste se produce con la bacteria Ralstonia eutropha y se comercializa con la marca Biopol manufacturado inicialmente por la empresa ICI (Zeneca), posteriormente vendida a Monsanto (1996) y finalmente obtenida por Metabolix (2001). La producción para el año 1988 fue del orden de las 800 toneladas. Biopol también se ha producido y vendido en EUA bajo el nombre de P3HBV y se ha vendido también en Japón. Otra empresa productora de estos polímeros es la austriaca Chemie Linz, la cual produjo comercialmente P3HB durante la década de 1980 y principio de la de 1990, usando la bacteria Alcaligenes latus, que es una cepa de rápido crecimiento que puede sintetizar P3HB durante crecimiento sin limitación de nutrientes (Braunegg et al. 1998). En Brasil existe otra empresa productora de P3HB llamada Biocycle. En esta industria se integra la producción de P3HB a la producción de etanol a partir de azúcar de caña. Muchas otras compañías lo han producido o producen actualmente y en el cuadro V se muestran las más importantes.
Aplicaciones de los PHA
Los PHA exhiben pesos moleculares relativamente altos, características termoplásticas o elastoméricas y otras propiedades físicas y mecánicas que los hacen candidatos para varias aplicaciones en la industria de empaques, medicina, farmacia, agricultura y alimentos, o como materias primas para la síntesis de químicos enantioméricamente puros y para la producción de pinturas (Babel y Steinbüchel 2001).
En el caso del P3HB, que es un termoplástico con propiedades similares al polipropileno, se usa principalmente para técnicas de moldeado. En cambio los copolímeros de P(3HBco3HV), tienen aplicaciones como material de empaque, productos higiénicos, agrícolas y biomédicos. La aplicación más conocida de estos biopolímeros de cadena corta es para la fabricación de botellas desechables para champú, contenedores para productos alimenticios, bolsas y otros productos desechables como pañales, servilletas, rastrillos, vasos y cubiertos (Anderson y Dawes 1990).
También es posible usarlos en la forma de látex acuoso para cubierta de materiales fibrosos como papel o cartón. Debido a su resistencia al agua, esta cubierta protege al papel o cartón contra el deterioro causado por la humedad. Otra aplicación de interés es su uso como material de empaque dado que estos PHA presentan una baja difusividad de oxígeno (Babel y Steinbüchel 2001).
En cuanto a las aplicaciones de los PHA de cadena corta en la agricultura, éstas consisten en macetas biodegradables, tubos de irrigación y matrices para la liberación controlada de factores de crecimiento, pesticidas y herbicidas. Una ventaja en este campo de aplicación es que no se requiere un grado de purificación muy alto del polímero, lo cual puede facilitar el proceso de extracción y hacerlo más económico (Babel y Steinbüchel 2001).
Con respecto a las aplicaciones biomédicas, éstas se basan en la propiedad de los PHA de ser biocompatibles con el tejido humano y reabsorbidos a una baja velocidad. Cuando este material se implanta en el cuerpo se hidroliza en metabolitos biocompatibles, por lo que se ha usado como hilo para sutura, sustitutos pericárdicos y sistemas de liberación de medicamentos. También se ha propuesto usarlos para fabricar jeringas desechables y lubricantes para guantes de cirugía. Dentro de las aplicaciones biomédicas, también está su uso como material osteosintético o de fijación para estimular la formación de hueso, así como el uso de sus productos de hidrólisis como materia prima para la industria farmacéutica en la que se requieren moléculas estero específicas (Babel y Steinbüchel 2001).
Dado que el P3HB y el P(3HBco3HV) han estado disponibles por varios años, el desarrollo de productos a partir de estos polímeros ha avanzado más que en el caso de los PHA de cadena media. Las investigaciones sobre estos últimos indican que podrían usarse para la fabricación de adhesivos, los cuales actualmente no son biodegradables y se hacen con base en polímeros sintéticos como, estireno y acetato de etilenvinil.
Otra aplicación de estos PHA se encuentra en el área de alimentos, específicamente como cubierta para quesos, que comúnmente son materiales no biodegradables con base en acetato de polivinilo, látex y ácido dibutil maleico (Babel y Steinbüchel 2001).
Los PHA de cadena media también se usan en la fabricación de hule biodegradable. Actualmente, los hules naturales y sintéticos como el de isopreno, estireno-butadieno, cloropreno y poliuretanos, se usan en aplicaciones como pañales, guantes, bandejas quirúrgicas, moldes y otros productos que son difíciles de reciclar o reusar. En investigaciones recientes se ha logrado fabricar hule biodegradable a partir de PHA de cadena media insaturados, por medio de enlaces entre los mismos. Esto se ha logrado por medio de reacciones químicas con sulfuros o peróxidos, o utilizando radiación UV. En general los PHA pueden ser manufacturados en diferentes materiales y formas, procesándolos con equipos convencionales para poliolefinas u otros plásticos, por ejemplo los de moldeado y extrusión. También pueden ser procesados en látex o en solución con diferentes solventes. Hasta el momento las aplicaciones comerciales han sido desarrolladas solamente para el producto Biopol. Este material ha sido procesado en botellas biodegradables para champú y para aceite para motor. Además, varios contenedores, rastrillos y bandejas para pescado y carne en la sección de refrigeradores en supermercados han sido manufacturados con Biopol y vendidos en Japón. En estos casos, este producto es usado por su imagen ambientalmente amigable para incrementar su venta y para otras aplicaciones que requieren biodegradabilidad por razones funcionales (Babel y Steinbüchel 2001).
En general, debido a su biodegradabilidad, resistencia al agua e impermeabilidad al oxígeno, los PHA pueden ser usados para todo tipo de material biodegradable. También se estima que su uso en artículos sanitarios desechables es económicamente factible. Otros mercados que se observan como prometedores, son de los de materiales de pesca, construcción e industria agrícola (Babel y Steinbüchel 2001).
En cuanto al potencial para aplicaciones biomédicas, éste es alentador, dado que el valor agregado de estos productos especiales es muy alto y técnica y económicamente parecen ser muy factibles.
Recientemente el desarrollo de pinturas y recubrimientos ambientalmente amigables basadas en PHA de cadena media, indica que tienen un potencial en países en donde las pinturas con base en solventes orgánicos han empezado a ser restringidas por sus gobiernos (Babel y Steinbüchel 2001).
Técnicamente, las perspectivas de los PHA son muy prometedoras, si el precio de estos materiales pudiera ser reducido, su aplicación sería muy atractiva. En este momento existe un esfuerzo mundial para producir PHA a partir de plantas como maíz y papa; así como la búsqueda de tecnologías baratas para su producción y uso en la manufactura de productos de uso diario. En este contexto, se está recurriendo a la búsqueda de cepas microbianas que por un lado sean capaces de producir PHA, y por otro, puedan emplear como sustratos residuos de las industrias agrícolas y pecuarias.
Rutas de biosíntesis de PHA en microorganismos
Como se ha mencionado previamente, el PHA más estudiado es el P3HB producido por la bacteria Ralstonia eutropha por lo que la ruta de biosíntesis en este microorganismo a partir de carbohidratos ha sido descrita a detalle. En general el proceso metabólico para la producción de PHA de cadena corta comienza a partir de acetil-CoA, en una secuencia de tres reacciones catalizadas por las enzimas 3-cetotiolasa (acetil-CoA acetiltransferasa, EC 2.3.1.9), acetoacetil-CoA reductasa (hidroxibutiril-CoA deshidrogenasa, EC 1.1.1.36) y la poli(3-hidroxibutirato) sintetasa (Fig. 5) (Anderson y Dawes 1990). Las rutas metabólicas para la biosíntesis de P3HB también han sido estudiadas en otras bacterias y las enzimas involucradas en el proceso también han sido caracterizadas. La enzima 3-cetotiolasa de bacterias como Azotobacter beijerinckii, Ralstonia eutropha, Zooglea ramigera y Rhizobium japonicum ha sido purificada y estudiada. Esta enzima ha mostrado ser la que controla la biosíntesis de P3HB en R. eutropha, con la CoA como el metabolito clave. Por su parte la enzima acetoacetil-CoA reductasa ha sido investigada en las bacterias Azotobacter beijerinckii, Rhodopseudomonas spheroides, Rhodomicrobium vannielii y Streptomyces coelicolor.

Con respecto a la P3HB sintetasa, se ha descubierto que en la mayoría de los casos está asociada con los gránulos de P3HB (Fig. 2), como en el caso de R. eutropha, R. rubrum, B. megaterium, A. beijerinckii y Z. ramigera. (Anderson y Dawes 1990). También se ha encontrado que existen cuatro clases principales de PHA sintetasas. Las clases I y II son enzimas que consisten en un solo tipo de subunidad (PhaC). Según su especificidad de sustrato, tanto in vivo como in vitro, la clase I utiliza preferentemente tioésteres-CoA de (R)-3-hidroxiácidos grasos de 3 a 5 átomos de carbono; mientras que las sintetasas clase II usan tioésteres-CoA de (R)-3-hidroxiácidos grasos de entre 6 y 14 átomos de carbono. La clase III de PHA sintetasas son enzimas que tienen dos tipos diferentes de subunidades, la PhaC y la PhaE. Las PHA sintetasas de la clase IV se parecen a las de la clase III pero en lugar de la subunidad PhaE tienen una subunidad PhaR. La polimerasa tipo III existe en Chromatium vinosum, Thiocystis violacea, Thiocapsa pfennigii y Synechocystis sp. PCC 6803. Este tipo de enzima polimeriza principalmente hidroxiácidos de cadena corta, pero también puede polimerizar a los de cadena media (Rehm 2003).
Uno de los estudios más detallados sobre la regulación de la síntesis de P3HB es el realizado en A. beijerinckii. Esta bacteria acumula P3HB bajo condiciones de limitación por oxígeno a partir de glucosa. La clave del control de la síntesis en este microorganismo es el destino de la acetil-CoA, la cual puede ser oxidada vía el ciclo de los ácidos tricarboxílicos (CAT) o puede servir como sustrato para la síntesis de P3HB. La ruta que tome la acetil-CoA depende de las condiciones ambientales, especialmente limitación por oxígeno cuando la relación NADH/NAD aumenta. La citrato sintetasa y la isocitrato deshidrogenasa se inhiben por el NADH y en consecuencia la acetil-CoA no entra al CAT en la misma cantidad, por lo que es convertida a acetoacetil-CoA por la 3-cetotiolasa la que a su vez es inhibida por CoA. Cuando la cantidad de oxígeno es adecuada, la concentración de CoA es alta y la acetil-CoA entra al ciclo CAT mientras que la 3-cetotiolasa es inhibida y consecuentemente la síntesis de P3HB cesa (Babel y Steinbüchel 2001).
Senior y Dawes (1971) propusieron que el P3HB servía no solamente como reserva de carbono y energía, sino que es un resumidero de poder reductor, considerado como un regulador redox dentro de la célula. Bajo condiciones de crecimiento limitadas por otros nutrientes se deja de sintetizar proteína y un proceso endergónico acoplado a la generación de ATP, vía la cadena de transferencia de electrones, podría dar lugar al incremento de la concentración intracelular de NAD(P)H propiciando un proceso similar al ya comentado anteriormente para A. beijerincki
En cuanto a la degradación interna del gránulo de P3HB, es también es un mecanismo complejo que requiere de la acción de varias despolimerasas (Luengo et al. 2003) el cual se describirá más adelante en este documento.
Generalmente las bacterias que sintetizan P3HB no producen PHA de cadena media, sin embargo con microorganismos recombinantes se pueden lograr mezclar las rutas metabólicas y producir polímeros o copolímeros con monómeros de cadena corta, media o ambas (Babel y Steinbüchel 2001).
Con respecto a la ruta metabólica para la biosíntesis de PHA de cadena media, la más estudiada es la que presentan las bacterias del género Pseudomonas. Por ejemplo, en Pseudomonas oleovorans los genes para la biosíntesis de PHA están presentes en el operón phaC1zC2D que codifica para dos polimerasas, PhaC1 y PhaC2, una despolimerasa (Paz) y la proteína PhaD. Las dos polimerasas catalizan la condensación en PHA de varios derivados de 3-hidroxiacil-CoA con cadenas de entre 6 y 14 carbonos. La mayoría de los intermediarios para la síntesis de PHA de cadena media se obtienen a partir de la β-oxidación (Fig. 6), aunque algunos otros monómeros pueden obtenerse de otras rutas (Luengo et al. 2003).

En el proceso de producción de PHA la provisión de 3-hidroxiacil-CoA tioésteres a la ruta metabólica, es decir la concentración de sustrato para la PHA sintetasa, es uno de los principales factores que determinan la masa molecular y la composición del biopoliéster, aunque también influye la existencia de despolimerasas, esterasas o lipasas inespecíficas que hidrolicen el polímero ya acumulado. Otro factor es el nivel de expresión de la proteína de PHA sintetasa activa (Rehm 2003). Por su parte la composición del PHA depende estrictamente de la especificidad de sustrato de la PHA sintetasa y del potencial metabólico del microorganismo para proveer (R)-3-hidroxiacil-CoA tioésteres a partir de la fuente de carbono empleada (Rehm 2003).
Técnicas analíticas para la detección, cuantificación y caracterización de PHA
Detección
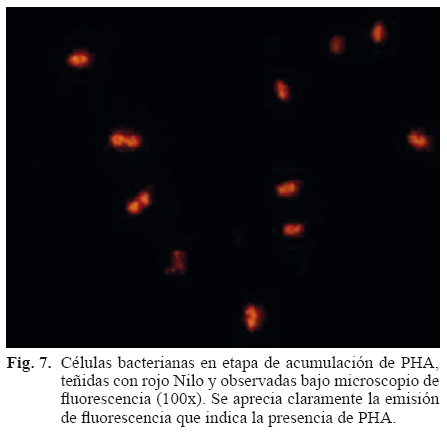
Tradicionalmente la detección de gránulos de PHA en células microbianas se ha llevado a cabo por medio de la tinción con el colorante lipofílico negro Sudán B, debido a la naturaleza lipídica de los PHA. Sin embargo actualmente existen otros colorantes que tienen mayor afinidad y especificidad por compuestos de naturaleza lipídica entre los que destaca el azul Nilo A (cloruro de 5-amino-9-dietilamino-benzo [alfa] fe nazonio), que pertenece a la familia de las oxazinas y que presenta una florescencia naranja a una longitud de onda de 460 nm (Ostle y Holt 1992). El rojo Nilo (7-diethilamino-3,4-benzofenoxazina-2-ona) es también un colorante excelente para la detección de inclusiones intracelulares de este tipo (Fig. 7). Los gránulos de PHA una vez teñidos presentan una fluorescencia que va del amarillo-dorado (excitación de 450-500 nm) al rosa-rojo (excitación de 515-560 nm) (Greenspan y Fowler 1985).
Por otro lado, la detección de PHA intracelular también puede llevarse a cabo mediante espectroscopia de infrarrojo (FT-IR). Los espectros de PHA con esta técnica exhiben un pico alrededor de los 1730 nanómetros correspondiente al enlace éster carbonilo característico de los PHA (Fig. 8) (Hong et al. 1999).
Extracción
La metodología reportada más frecuentemente para la extracción de PHA de la biomasa microbiana ha sido el uso de hidrocarburos clorados, especialmente la técnica utilizando reflujo con cloroformo (Fuchtenbusch et al. 1996). La solución de PHA resultante se filtra para remover restos de células, luego se concentra y el PHA se precipita en metanol o etanol (Fig. 9). Con esta técnica los lípidos de bajo peso molecular se queden en solución y no interfieren con la determinación. El uso de solventes clorados es más utilizado para la extracción de PHA de cadena corta como el P3HB, sin embargo los PHA se cadena media son solubles en un rango de solventes más amplio. Debido a que el uso a gran escala de los solventes mencionados anteriormente puede ser costoso, también se han desarrollado otros procesos de extracción con base en el uso de etilen y propilen carbonato (Fiorese et al. 2009), así como metodologías que se basan en la liberación del polímero de las células mediante la ruptura de las mismas usando soluciones de hipoclorito de sodio, ácidos o bases.

Cuantificación y análisis fisicoquímico
Con respecto a la cuantificación de PHA uno de los métodos que se utilizaron inicialmente fue el gravimétrico, que consiste en usar cloroformo como solvente para extraer el polímero de la biomasa liofilizada, posteriormente precipitarlo con dietil éter o acetona, y finalmente secarlo y pesarlo (Grothe et al. 1999). Otra metodología para hacer la cuantificación gravimétrica se basa en el hecho de que, bajo condiciones controladas de tiempo y temperatura, el material celular (excepto los gránulos de P3HB) se disuelve en una solución alcalina de hipoclorito de sodio de manera que el bioplástico se libera y es recuperado por centrifugación y posteriormente secado y pesado (Hahn et al. 1994).
Por su parte, la técnica espectrofotométrica para la cuantificación de P3HB se basa en la conversión del polímero a ácido crotónico mediante el uso de ácido sulfúrico concentrado y la aplicación de calor. El ácido crotónico producido se cuantifica por espectrofotometría UV leyendo absorbancia a 230 nm y correlacionando los valores con una curva estándar del ácido (Law y Sleepecky 1961).
Respecto a la técnica cromatografía de gases (CG), la más utilizada se fundamenta en la metodología desarrollada por Braunegg et al. (1978) que consiste en someter a las células liofilizadas a una metanólisis ácida o alcalina. Con este proceso se generan metal ésteres de los monómeros que componen el PHA y posteriormente se analizan por CG con detector de ionización de flama. Mediante el uso de un CG acoplado a un detector de masas, esta técnica no sólo funciona para cuantificar PHA sino también para determinar su composición monomérica (Mittendorf et al. 1998).
La determinación de PHA en células intactas utilizando espectroscopia de fluorescencia bidimensional y citometría de flujo, también ha sido propuesta. En este procedimiento las células teñidas con azul Nilo muestran un máximo de fluorescencia entre 570 y 605 nm cuando son excitadas entre 540 y 560 nm, y se ha encontrado una buena correlación entre la intensidad de fluorescencia y la concentración de PHA (Vidal-Mas 2001).
Otro método de cuantificación es la cromatografía iónica que se basa en la conversión de los monómeros de PHA a ácidos alcanoicos mediante propanólis ácida seguida de una hidrólisis alcalina con Ca(OH)2 o ácida con H2SO4 concentrado. La muestra se inyecta en una columna aniónica con detector de conductividad (Hesselmann et al. 1999).
También se ha establecido un método de cuantificación enzimática de PHA a partir del equipo desarrollado por Roche Molecular Biochemicals, de los EUA (No. 127833). La técnica implica la oxidación de 3HB y la reoxidación del NADH producido a partir de NAD dando lugar a la producción de formazan, que se lee espectrofotométricamente a 492 nm.
En cuanto a las técnicas para determinar la composición monomérica, peso molecular y propiedades térmicas de los PHA destacan: la cromatografía de gases acoplada a masas y la resonancia magnética nuclear (RMN); la cromatografía de filtración en gel; y la calorimetría diferencial de barrido, respectivamente. En el caso de las técnicas por NMR éstas se han aplicado exitosamente en el estudio de la estructura de los PHA. El análisis del espectro H RMN permite determinar la composición del polímero y la distribución de las unidades monoméricas puede deducirse de las secuencias de diadas y triadas por análisis espectral de 13C RMN. Jacob et al. (1986) demostraron que era posible también utilizar espectro 13C RMN de polarización cruzada de células liofilizadas para monitorear directamente, y de manera no destructiva, PHA acumulado intracelularmente. En relación con la metodología utilizada para la determinación de peso molecular, la cromatografía de exclusión molecular o cromatografía de filtración en gel utilizando como solvente cloroformo a 30 °C, un conjunto de columnas de exclusión molecular y estándares de poliestireno de baja polidispersidad (Lee et al. 1995) es la que se aplica con mayor frecuencia. Para la determinación de las propiedades térmicas se utiliza la calorimetría diferencial de barrido debido a que los PHA son polímeros parcialmente cristalinos, y la definición de sus propiedades térmicas y mecánicas se expresa en términos de las temperaturas de transición de la fase amorfa y de la temperatura de fusión de la fase cristalina (Zagar y Krzan 2004).
Procesos de fermentación para producir PHA
Muchas bacterias se han investigado para la producción de PHA, principalmente de cadena corta. Con base en esto se han desarrollado procesos y sistemas de cultivo muy variados, y se ha estudiado la influencia de diferentes parámetros sobre la síntesis de estos biopoliésteres a partir de una gran diversidad de sustratos simples y complejos. En la figura 9 se presenta un esquema general de producción de estos biopolímeros. Los rendimientos y productividades alcanzadas por varios microorganismos cultivados en diferentes sistemas de cultivo, a partir de diversos sustratos, se pueden apreciar en el cuadro VI.
De manera general las bacterias usadas para la producción de PHA pueden ser divididas en dos grupos, según las condiciones de cultivo que requieren para la síntesis del polímero. El primer grupo de bacterias requiere de la limitación de un nutriente esencial (nitrógeno, fósforo, azufre, magnesio, oxígeno), para sintetizar PHA a partir de un exceso de fuente de carbono. Este grupo incluye a Ralstonia eutropha, Pseudomonas extorquens y Pseudomonas oleovorans, entre otras (Khanna y Srivastava 2005).
El segundo grupo de bacterias no requiere de la limitación de nutrientes para la síntesis de PHA y pueden acumular polímero en grandes cantidades durante la fase de crecimiento. Este grupo incluye a Alcaligenes latus, Azotobacter vinelandii recombinante y E. coli recombinante, entre otras, que acumulan grandes cantidades de polímero durante la fase de crecimiento exponencial (> 50 %), a diferencia del otro grupo de bacterias que durante esta fase acumulan muy bajas cantidades de polímero (Braunegg et al. 1998).
Con respecto a las estrategias de cultivo para la producción de PHA, se ha visto que se puede alcanzar una productividad alta mediante cultivo por lote alimentado o cultivo continuo. Para cultivo por lote alimentado de bacterias que requieren de la limitación por algún nutriente para sintetizar PHA, se utiliza un sistema en dos etapas. En la primera se obtiene una concentración de biomasa deseada sin limitación de nutrientes. En la segunda etapa se promueve la síntesis de PHA manteniendo en concentraciones limitantes uno de los nutrientes esenciales. En esta segunda etapa la concentración biomasa sin-PHA permanece casi constante y el incremento en biomasa se debe a la acumulación intracelular del polímero. R. eutropha acumula una gran cantidad de PHA (hasta 80 % de su peso en base seca) cuando la fuente de nitrógeno o fósforo es totalmente consumida y aún hay glucosa disponible en el medio. Por otro lado, otras bacterias de este mismo tipo producen PHA más eficientemente cuando uno de los nutrientes está limitado, pero no totalmente consumido (Khanna y Srivastava 2005).
Para el cultivo de bacterias con producción de PHA asociada al crecimiento, se pueden alcanzar productividades altas en un sistema de lote alimentado en una sola etapa. La estrategia de alimentación es muy importante para obtener una densidad celular alta y no tiene que ver con la limitación de nutrientes, por lo que se pueden usar fuentes de nitrógeno complejas para mejorar el crecimiento (Khanna y Srivastava 2005).
En cuanto al desarrollo de estrategias para la producción de PHA, existe una mayor cantidad de estudios sobre el proceso utilizando microorganismos que polimerizan monómeros de cadena corta (scl-PHA). Se ha llegado a la conclusión de que algunos metilotrofos y metanotrofos son interesantes para la producción de P3HB. El metanol es un sustrato barato y existe mucha información sobre el cultivo de estas bacterias ya que muchas han sido consideradas para la producción industrial de proteína celular a gran escala. Dentro de este grupo se incluyen a Protomonas extorquens, Paracoccus denitrificans y Methylobacterium extorquens (Suzuki et al. 1986). Esta última sintetiza un copoliéster de P(3HBco3HV) cuando se proporciona simultáneamente alcohol amílico y metanol en un medio limitado por fuente de nitrógeno (Ueda et al. 1992, Yamane et al. 1996). Paracoccus sp.12-A es también capaz de acumular P(3HBco3HV) cuando crece a partir de ácido fórmico (Mineki et al. 1997) y Methylobacterium organophilum usando metanol como sustrato (Kim et al. 1996). De forma interesante la capacidad de los metanotrofos de crecer a partir de hidrocarburos clorados puede ser también explotada para la producción de P3HB. Por ejemplo Methylosinus trichosporium OB3b es capaz de degradar tricloroetileno que es un contaminante común del agua subterránea para producir P3HB (Shah et al. 1996). Azotobacter vinelandii UWD es otra cepa que vale la pena mencionar (Page y Knosp. 1989)siendo capaz de producir P3HB a partir de fuentes de carbono baratas. Sintetiza P3HB durante el crecimiento exponencial usando jarabe de maíz, melazas de caña, melaza de remolacha y extracto de malta con rendimientos altos (Page 1989). Incluso, se pueden obtener polímeros con pesos moleculares extraordinariamente altos (por arriba de 4 millones de Daltones), cuando esta cepa crece en un medio de melazas. De forma notable, la adición de valerato como cosustrato permite la producción de un copolímero (Page et al. 1992).
La producción de P3HB también es una característica común entre las bacterias fototrópicas anoxigénicas. Algunas de las bacterias de este grupo que han sido estudiadas para la producción de P3HB son de los géneros Chromatium, Thiocystis, Thiocapsa, Rhodococcus, Rhodobacter y Rhodospirillum, que pertenecen a las bacterias púrpura (las tres primeras) y no púrpuras (las tres últimas). Creciendo sobre acetato, la mayoría de estas bacterias produce solamente un homopolímero de P3HB. Chromobacterium violaceum es una bacteria heterótrofa que es capaz de producir homopoliésteres de hidroxivalerato, cuando se cultiva en ácido valérico como fuente de carbono (Steinbüchel et al. 1993).
R. eutropha es de hecho una bacteria autótrofa oxidadora de hidrógeno que puede producir P3HB a partir de CO2 y H2. El factor crítico en este proceso autótrofo es evitar posibles explosiones de gas. Por lo tanto ha sido desarrollado un circuito cerrado para reciclar gas dentro de los márgenes de seguridad (Tanaka et al. 1995). Por otro lado, las cianobacterias como Synechococcus sp. MA19 también son capaces de producir P3HB usando CO2 como fuente de carbono (Miyake et al. 1996).
También se ha descrito un nuevo proceso para lo foto conversión de desperdicios orgánicos en biopoliésteres. En este proceso la biomasa seca se gasifica térmicamente y resulta una mezcla de gases compuesta de CO2 y H2. Las bacterias fotosintéticas asimilan los componentes del gas en nueva masa celular (Maness y Weaver 1994).
Con el objeto de producir P3HB a partir de sustratos baratos, también han sido estudiados métodos de cultivo por lote alimentado en dos etapas, usando dos diferentes microorganismos, el primero transforma la fuente de cultivo barata en sustratos asimilables para que el segundo microorganismo los use para producir PHA (Tanaka et al. 1993).
Por otro lado, como se mencionó anteriormente, los PHA son casi siempre 3-hidroxi ésteres; sin embargo, también hay ciertas variaciones. La más común es la ocurrencia de 4-hidroxi ésteres. Entre muchas diferentes cepas, R. eutropha, A. latus y C. acidovorans son capaces de sintetizar copolímeros de 3- hidroxibutirato y 4-hidroxibutirato cuando crecen en medios con ácido 4-hidroxibutírico, 1,4-butanodiol, butirolactona o ácido 4-clorobutírico (Renner et al. 1996). También R. eutropha produce un copolímero de 3HB y 4HB a partir de mezclas de fructosa y butirolactona (Doi et al. 1990). Se pueden producir homopolímeros de 4HB si se usa 1,4 butanodiol o ácido 4-butírico como única fuente de carbono (Saito y Doi 1994).
Más aún, las condiciones de cultivo también pueden influenciar la composición del polímero. Se ha demostrado que la adición de 2 % de polietilenglicol da como resultado una mayor incorporación de 4-hidroxibutirato (Shi et al. 1996). Se han encontrado polímeros que contienen monómeros de 4- hidroxivalerato en bacterias del género Ralstonia y de la cepa de Pseudomonas oxalaticus (Valentin et al. 1992). También han sido descritos poliésteres conteniendo 2-hidroxi, 5-hidroxi, o 6-hidroxi monómeros (Williams 1999).
R. eutropha produce un terpolímero de 3-hidro-xibutirato, 3- hidroxivalerato y 5-hidroxivalerato al alimentarla con 5-clorovalerato o 5-hidroxivalerato y ácido valérico (Doi et al. 1987).
Análisis con RMN de PHA aislados de Pseudomonas aeruginosa 44T1 cultivadas en aceite de euforbia y castor, sugieren la presencia de 3,6-dihidroxidodecanoato, 6-hidroxi-3-dodecenoato y 4-hidroxidecanoato (Eggink et al. 1995). Mediante el uso de bacterias recombinantes también se han producido PHA con monómeros de 5-hidroxihexanoato, 4-hidroxiheptanoato y 4-hidroxioctanoato (Valentin et al. 1996).
Otra variante de la estructura de los PHA es la presencia de grupos metilo en el carbono 2. R. eutropha y Burkholderia cepacia, por ejemplo, acumulan un terpoliéster de 3-hidroxibutirato, 3-hidroxivalerato y 2-metil-3-hidroxibutirato (Fü-chtenbusch et al. 1996). Al crecer en medio mineral con glucosa y 3-hidroxipivalato se produce un ácido alcanoico con dos grupos metilos sustituyentes en el átomo de carbono alfa por Rhodococcus ruber y bacterias relacionadas.
Por otro lado, en lodos anaerobios es común encontrar PHA conteniendo monómeros de 2-metil-3-hidroxivalerato. El proceso de usar aguas residuales (cultivos mixtos) para la producción de PHA, también ha sido investigado profundamente ya que el uso de sustratos de bajo costo es parte esencial de la investigación sobre la producción de PHA.
En términos generales, la rentabilidad de una bacteria para producir PHA a escala industrial depende de varios factores como son: la estabilidad e inocuidad del organismo, la velocidad de acumulación del polímero, la velocidad de crecimiento, la densidad celular que se puede alcanzar, el contenido de polímero, la facilidad de extracción, el peso molecular del PHA, el rango de fuentes de carbono utilizables, el costo del medio de cultivo y la generación de subproductos.
La empresa ICI (Inglaterra) fue la primera en interesarse por la producción a gran escala de P3HB, para lo cual evaluó a tres microorganismos: Azotobacter sp. , Methylobacterium sp. y Ralstonia eutropha. El microorganismo que decidieron usar para el proceso a nivel industrial fue una cepa de R. eutropha con la cual han producido P3HB y un copoliéster de 3HB y 3HV (Byrom 1993). El equipo que esta empresa ha usado para la producción de estos biopolímeros es un fermentador air-lift de 35 000 litros y varios tanques de agitación de 200 000 L (Schlegel y Gottschalk 1965). La fermentación se lleva a cabo en un sistema por lote alimentado en dos etapas, en el que se utiliza la limitación por fosfato como factor para estimular la síntesis de P3HB. Durante la primera etapa las células crecen en un medio mineral con glucosa como fuente de carbono y energía y con la cantidad de fosfato necesaria para producir cierta cantidad de biomasa. A medida que crece la bacteria, el fosfato se consume del medio y durante la segunda etapa, cuando el fosfato está en concentraciones limitantes, las células comienzan a acumular el polímero. Se proporciona glucosa al cultivo y la fermentación continua hasta que el contenido de polímero deseado se alcanza. Cada fase dura aproximadamente 48 h y se alcanza una concentración de biomasa en base seca de 100 g/L (Byrom 1992).
Se ha observado que durante las primeras 60 h, la acumulación de P3HB es poca, pero después de que inicia la alimentación con glucosa se logra que el polímero se acumule hasta en un 75 % del peso de la biomasa en base seca.
El copolímero de 3HB y 3HV se produce agregando una mezcla de glucosa y ácido propiónico en la fase de acumulación. El contenido de hidroxivalerato en el copolímero se regula ajustando la relación de los dos sustratos, cuidando que el propionato se mantenga en una concentración menor a 1 g/L para evitar efectos tóxicos (Byrom 1992).
La recuperación del polímero se hace mediante un proceso de reflujo con metanol caliente para remover lípidos y fosfolípidos de las células, seguido de extracción del P3HB con cloroformo o cloruro de metileno. La solución se filtra para remover restos celulares, se enfría y se precipita para luego ser secada a vacío.
Para reducir costos, ICI también ha usado una estrategia en la que las células se desintegran por shock térmico, luego son tratadas con detergentes y enzimas para disolver los componentes celulares que no sean P3HB, posteriormente el polímero se lava y flocula y finalmente se recupera como un polvo blanco del cual se hacen hojuelas.
Generalmente se requiere alrededor de 2.1 g de glucosa para producir 1 g de P3HB, pero como también se se necesita de una fuente de carbono y energía para el crecimiento, en la práctica se ha observado que se precisan 3 g de glucosa para producir 1g de P3HB.
En 1996, este proceso de producción de biopolímeros fue adquirido por Monsanto, quienes detuvieron la producción de P3HB y P(3HBco3HV) a finales de 1998.
Un proceso industrial diferente se desarrolló en la empresa Biotechnologische Forschungsgesellschaft en Linz (Austria) para la producción de P3HB (Hánggi 1990, Hrabak 1992). Este proceso se basó en el uso de Alcaligenes latus DSM1124, el cual puede acumular P3HB hasta en un 80 % de su peso en base seca durante crecimiento balanceado. Gracias a esto, la fermentación se da en un solo paso usando un cultivo por lote alimentado en un medio de sales minerales con sacarosa como fuente de carbono. Aunque se alcanzó una biomasa celular de 60g/L, la compañía detuvo la producción en 1993 (Hrabak 1992).
En el caso de la empresa brasileña Biocycle, dentro de la planta productora de azúcar y etanol, está instalada una planta productora de P3HB a partir de los residuos de la extracción del azúcar de caña. Esto permite tener un ciclo cerrado en el que los desperdicios de un proceso se utilizan como insumos para el otro, como el caso del agua residual de la fermentación para producir P3HB que se utiliza para regar los sembradíos de caña, y el uso del bagazo de caña residual para generar energía para el funcionamiento del complejo de producción (Nonato et al. 2001).
En general, el acoplar el proceso de producción de P3HB en una fábrica de azúcar y etanol, hace posible que se reduzca el precio por kilogramo de P3HB de 4 a 5 veces en comparación con otros procesos para producir el mismo polímero (Nonato et al. 2001). Por su parte los PHA de cadena media no han sido producidos en escala comercial. Consecuentemente, los procesos para producción y recuperación de PHA de cadena corta han recibido mucha más atención (Wang y Lee 1997). Cuando se comparan cultivos por lote alimentado, la única diferencia entre PHA de cadena corta y PHA de cadena media, es el bajo contenido celular de polímero que se encuentra en estos últimos, lo cual afecta el proceso y lo hace menos rentable en comparación al P3HB, ya que el costo de recuperación de los de cadena media es más elevado (Choi y Lee 1999).
Biodegradación de PHA
La composición monomérica de los PHA como ya fue mencionado es altamente variable, lo que determina las características físicas y químicas de los mismos y por consecuencia su susceptibilidad a ser biodegradados. El gran tamaño de los polímeros individuales les impide ser transportados a través de la membrana celular, por lo tanto el microorganismo debe de tener la capacidad de hidrolizarlos en sus correspondientes monómeros (hidroxiácidos) (Williams y Peoples 1996). Los PHA pueden ser biodegradados por una amplia variedad de microorganismos ubicuos en muchos ecosistemas hasta dióxido de carbono o metano, tanto en condiciones aerobias como anaerobias, sin la formación de productos tóxicos.
Chowdhury reportó por primera vez la degradación de P3HB por microorganismos de los géneros de Bacillus, Pseudomonas y Streptomyces (Chowdhury 1963). Estos microorganismos han sido aislados del suelo (Pseudomonas lemoigne, Comamonas sp., Acidovoraxfaecalis, Aspergillus fumigatus y Variovorax paradoxus), lodos activados y anaerobios (Alcaligenes faecalis, Pseudomonas sp., Illyobacter delafieldi), agua dulce (lagos) y salada (mar) (Comamonas testosterone, Pseudomonas stutzeri) (Lee 1996). El porcentaje de microorganismos degradadores de P3HB en el ambiente fue estimada entre 0.5-9.6 % del total de colonias (Suyama et al. 1998).
Se han realizado estudios ecológicos y taxonómicos sobre la abundancia y diversidad de microorganismos degradadores de P3HB y P(3HBco3HV) en suelo, agua y compostas, tanto in situ como bajo condiciones de laboratorio. Las principales variables consideradas fueron el tiempo, el tipo de ambiente, la temperatura de incubación y el % de hidroxivalerato (3HV) (Mergaert et al. 1992). Un total de 330 microorganismos degradadores fueron aislados e identificados: 154 bacterias, 77 estreptomicetos y 99 hongos. Las bacterias degradadoras de PHA están divididas en once grupos con base en la especificidad de las despolimerasas por su sustrato (Jendrossek et al. 1996) y los hongos en al menos 95 géneros conocidos como degradadores de PHA. La mayoría de las bacterias caracterizadas presentan una especificidad dependiente de la longitud de la cadena del polímero (corta o media), aunque algunas manifiestan la capacidad de utilizar una variedad importante de polímeros (Jendrossek et al. 1996) (Cuadro VII). Se han identificado muchos hongos degradadores de P3HB (Penicillium, Aspergillus y Paecilomyces) (Mergaert et al. 1993, Oda et al. 1997, Han et al. 1998, Gonda et al. 2000, Miyazaki et al. 2000). La capacidad de los hongos para degradar P3HB ha sido evaluada en función de su afiliación sistemática, encontrando que miembros de basidiomicetos, deuteriomicetos, ascomicetos, zigomicetos, chitridiomicetos y mixomicetos son capaces de degradar P3HB (Matavulj y Molitoris 1992). Sin embargo, no se conoce cuál de los dos grupos de microorganismos contribuye más en la degradación de P3HB en el ambiente. El aislamiento de estos microorganismos ha permitido realizar estudios de biodegradación in vitro de diferentes tipos de PHA (Cuadro VII). La degradación de P3HB ha sido reportada principalmente a temperaturas mesofílicas, aunque Tokiwa et al. enfatizan que el compostaje a altas temperaturas es una de las tecnologías más prometedoras para el reciclado de plásticos biodegradables (Abe y Doi 1999). La velocidad de degradación de PHA, aún bajo condiciones ambientales controladas es difícil de predecir. Normalmente altas temperaturas permiten una mejor degradación, probablemente debido al incremento en la actividad microbiana. Es importante notar que a temperaturas elevadas se mejora de manera importante la biodisponibilidad y solubilidad de compuestos poliméricos orgánicos y de contaminantes ambientales biodegradables, permitiendo su eficiente biorremediación (Müller et al. 1998). A altas temperaturas, el P3HB es amorfo y más sensible al ataque enzimático permitiendo su rápida degradación (Takeda et al. 1998). Mientras existen numerosos reportes sobre bacterias degradadoras de polímeros en condiciones mesofílicas, sólo recientemente se han publicado trabajos de bacterias termofílicas y de enzimas termoestables para dicho propósito (Elbanna et al. 2004). Los microorganismos termofílicos capaces de degradar varias clases de poliésteres a altas temperaturas son de interés y entre éstos los actinomicetos han demostrado ser los más importantes. Estos han sido aislados de diferentes ecosistemas, encontrando que de 341 cepas, 31 aisladas fueron capaces de degradar P3HB, PCL (policaprolactona) y polietilen succinato (PES). Estas fueron identificadas como miembros de los géneros: Actinomadura, Microbispora, Streptomyces, Thermoactinomyces y Saccharomonospora (Yamashita et al. 2001). Una cepa termofílica de Streptomyces sp. aislada de suelo ha sido capaz de degradar P3HB y también polietilen succinato (PES), polibutilen succinato (PBS) y poli (oligo(tetrametilen succinato)-co-tetrametilen carbonato)) (PBS/C). Este actinomiceto ha presentado una actividad degradadora de P3HB superior a la de cepas termotolerantes y termofílicas procedentes de colecciones de cultivo (Kasuya et al. 1999). Recientemente, en las bacterias termófilas Anoxybacillus gonensis G2 (Colak et al. 2005) y Thermus thermophilus HB8 (Papaneophytou et al. 2009) se ha reportado una despolimerasa extracelular degradadora de P3HB. La degradación de P3HB ha sido también reportada en una cepa termotolerante de Aspergillus sp. alcanzándose una degradación del 90 % de una película de P3HB después de 5 días de cultivo a 50 °C (Handrick et al. 2001). El aislamiento de estos microorganismos ha permitido realizar estudios de biodegradación in vitro de diferentes tipos de PHA. En contraste, comparativamente han sido aislados pocos microorganismos degradadores de PHA de cadena media (mcl-PHA). Alrededor de 26 microorganismos degradadores de mcl-PHA han sido aislados de suelo incluyendo numerosos del género Pseudomonas, una cepa de Comamonas y de Alcaligenes (Schirmer et al. 1993, Foster et al. 1995). Seis géneros relativamente no muy comunes de microorganismos degradadores de PHA de cadena media fueron aislados de suelos contaminados con hidrocarburos composteados (Ramsay et al. 1994).
La velocidad de biodegradación del polímero depende de varios factores, incluyendo el área superficial del polímero, actividad microbiana, pH, temperatura, humedad y la presencia de otros nutrientes (Lee 1996). Los parámetros del polímero que afectan tal velocidad incluyen la composición monomérica del polímero, el nivel de cristalinidad o de regiones amorfas (a mayor cristalinidad menor biodegradación) (Nishida y Tokiwa 1995, Spyros y Kimmich 1997) y el peso molecular (polímeros de bajo PM se degradan más rápidamente que los de alto PM) (Jendrossek et al. 1996). Los microorganismos colonizan la superficie de los polímeros, secretando despolimerasas, las que hidrolizan los enlaces éster de los PHA generando oligómeros, dímeros que subsecuentemente son llevados a sus formas monoméricas (p.e. P3HB a hidroxibutirato, P3HV a hidroxivalerato, etc.) por hidrolasas de oligómeros (Shirakura et al. 1983). Estos monómeros son suficientemente pequeños para pasar a través de la membrana microbiana semipermeable para ser metabolizados por P-oxidación y el ciclo de los ácidos tricarboxílicos en dióxido de carbono y agua en condiciones aeróbicas (Scott 1990), mientras que en condiciones anaerobias también es producido metano (Luzier 1992).
La biodegradación de PHA puede ser claramente distinguida entre la realizada al interior y al exterior de la célula a partir del hecho que dicho polímero puede estar presente en dos conformaciones biofísicas. En la biodegradación intracelular los gránulos amorfos de PHA se acumulan in vivo y presentan una capa superficial intacta compuesta de proteínas y fosfolípidos, y en la extracelular la pérdida de dicha capa (después de la lisis celular o de la extracción del polímero) provoca la coalescencia de los gránulos y la formación de cristales de la estructura amorfa previa del polímero (Merrik et al. 1965, Jendrossek 2001). La mayoría de las enzimas que hidrolizan P3HB son específicas para las formas nativa y desnaturalizada de P3HB (Handrick et al. 2004). La degradación enzimática de PHA está influenciada por la composición monomérica, estructura estereoquímica, distribución de la secuencia, área superficial y morfología (Scherer et al. 2010).
La capacidad de utilizar extracelularmente PHA como fuente de carbono y energía está ampliamente distribuida entre bacterias (Gram + y -), hongos y actinomicetos, sin estar limitada únicamente a organismos productores de PHA. La degradación extracelular se presenta en microorganismos no necesariamente acumuladores de PHA, que les permite utilizar como fuente de carbono exógena estos polímeros extracelulares liberados por células muertas acumuladoras de PHA. Las enzimas clave que participan en ambos procesos de degradación son denominadas de manera genérica como despolimerasas (intra y extracelulares) específicas de PHA (despPHA), las cuales son carboxiesterasas (EC 3.1.1) que hidrolizan los polímeros insolubles en agua en monómeros y oligómeros solubles. La mayoría de las bacterias producen despPHA que son específicas para polímeros de cadena corta o media, aunque existen algunas excepciones (Schirmer et al. 1995).
La degradación microbiana y enzimática de PHA puede ser abordada desde dos puntos de vista: uno basado en las características microbianas (enzimática) y otro sobre las correspondientes de los bioplásticos. Los PHA pueden ser utilizados como fuente de carbono por una amplia variedad de bacterias y de eucariontes. Estos microorganismos emplean varias despolimerasas específicas de estos polímeros (Uefuji et al. 1997, Schóber et al. 2000), las cuales son carboxiesterasas (EC 3.1.1.75 y EC 3.1.1.76) (Jendrossek y Handrick 2002). La degradación enzimática de polímeros por hidrólisis es un proceso de dos pasos en la interface sólido-líquido: primero, la enzima enlaza al polímero-sustrato y susecuentemente cataliza el rompimiento hidrolítico.
Las propiedades de las despolimerasas de P3HB (despPHB) han sido ampliamente estudiadas, teniendo en común varias características bioquímicas tales como: peso molecular relativamente pequeño (<100 kDa); están conformadas por una sola cadena polipeptídica; no se enlazan a intercambiadores aniónicos como la dietilaminoetil-celulosa, aunque tienen una fuerte afinidad a materiales hidrofóbicos tales como el butilo o fenilo Toyopearl; el pH óptimo está entre 7.5-9.8, a excepción de la polimerasa de Pseudomonas picketti y de Penicillium funiculosum que se encuentra entre 5.5 y 7; alta estabilidad en un amplio rango de pH, temperatura y fuerza iónica. La mayoría de las despolimerasas bacterianas no son glicosiladas aunque las de eucariontes y de Pseudomona lemoignei sí lo son. Esta característica no es esencial para la actividad, pero podría ayudar en la estabilidad de la enzima. Todas las despolimerasas son específicas para la configuración (R) aunque los polímeros (R-S)-3HB son degradados en oligómeros más largos puesto que al parecer no son capaces de hidrolizar los enlaces éster entre monómeros de la configuración (S). La mayoría de estas enzimas parecen ser serina hidrolasas, las que son específicas para PHA de cadena corta o media; son inhibidas por inhibidores de serina esterasas (diisopropil-fluoril fosfato o compuestos acil sulfonilo), los que se enlazan covalentemente con la serina del sitio activo de las serina hidrolasas (Jendrossek et al. 1996, 1998).
La hidrólisis enzimática de PHA se lleva a cabo primero en la región amorfa y subsecuentemente en la cristalina. Para películas de poli-3-hidroxibutirato con grados similares de cristalinidad, la velocidad de hidrólisis enzimática depende del tamaño y de la estructura de las esferulitas (cristalinas).
Varias despolimerasas de PHA han sido aisladas y purificadas de microorganismos pertenecientes a los géneros Alcaligenes (Shirakura et al. 1986, Bachmann y Seebach 1999), Comomonas (Jendrossek et al. 1993, Kasuya et al. 1994) y Pseudomonas (Mukai et al. 1993, Mukai et al. 1994, Schóber et al. 2000). En la última década más de 20 diferentes despolimerasas extracelulares de P3HB han sido caracterizadas (Jendrossek y Handrick 2002). Estas han sido analizadas a nivel de sus genes estructurales, encontrando que mientras la mayoría de las bacterias degradadoras de PHA contiene únicamente una despolimerasa, aunque Paucimonas lemoignei presenta siete despolimerasas con diferente especificidad hacia los diferentes sustratos (Jendrossek y Handrick 2002). Las despolimerasas son secretadas al ambiente y catalizan la degradación endo y exo de las cadenas de PHA. El ataque exo ocurre estrictamente en la parte terminal o extremo del polímero, frecuentemente con una preferencia por uno de éstos (p.e. el hidroxilo final del hidroxiácido del poliéster es atacado más que el ácido carboxílico final del otro extremo), produciendo exclusivamente pequeños oligómeros o monómeros metabolizables. En el caso del ataque endo, éste ocurre en cualquier lugar a lo largo de la cadena del polímero, generando una mezcla de oligómeros y una reducción rápida del peso molecular (Lenz 1993). Las despolimerasas también pueden combinar sinérgicamente los dos tipos de actividad. La secreción de las despolimerasas ocurre principalmente al final de la fase de crecimiento exponencial y es usualmente reprimida en presencia de una fuente de carbono soluble, mientras que el agotamiento de nutrientes promueve la síntesis de éstas (Jendrossek et al. 1996). El metabolismo de 3HB suprime la producción de enzimas y causa una desaparición de la enzima secretada. La despolimerasa extracelular podría representar al menos el 25 % o más del total de la proteína extracelular. También el 3HB parece inhibir la excreción de la despolimerasa, lo cual indica que su producción podría estar sujeta a represión catabólica. Es posible que esta enzima sea reabsorbida por las células o que sea destruida en la superficie de la célula (Delafield et al. 1965). La estructura de estas enzimas secretadas al medio incluye varios dominios y un péptido señal. Estas proteínas despolimerasas de PHA (PhaZ) tienen un amplio dominio catalítico (320-350 aminoácidos) en el extremo N-terminal, un dominio de enlace al sustrato (40-50 aminoácidos) en el extremo C-terminal y un dominio de unión (50-100 aminoácidos) conectando los dominios (Jendrossek y Handrick 2002). El dominio de enlace es el responsable de la adsorción de la enzima a la superficie del polímero insoluble en agua (PHA sólido), lo que permite al dominio catalítico interactuar con la cadena del polímero (Fukui et al. 1998). El sitio activo está conformado por el dominio catalítico constituido por tres aminoácidos altamente conservados semejante al de lipasas: serina, aspartato e histidina (Brucato y Wong 1991) conocido como caja lipasa (lipase box) y un grupo de aminoácidos que estabilizan el oxianión. La serina que ataca nucleofílicamente el enlace éster es parte del pentapéptido (Gly-X-Ser-X-Gly) que forma la caja lipasa, la cual ha sido encontrada en todas las hidrolasas conocidas tales como lipasas, esterasas y proteasas séricas (Jaeger et al. 1994). Recientemente se han clasificado las despPHA en tres tipos en función de la estructura de su dominio de enlace del anclaje al sustrato o de la posición de la caja lipasa en el dominio catalítico (Jendrossek et al. 1995, Klingbeil et al. 1996). Entre las enzimas, de tipo I se encuentran la PhaZ4 de Pseudomonas lemoignei (Jendrossek et al. 1995b) y la de Alcaligenes faecalis T1 (actualmente Ralstonia pickettii T1) (Shiraki et al. 1995), mientras que las de tipo II son producidas por Comamonas testoteroni (Jendrossek et al. 1995), Streptomyces exfoliatus (Klingbeil et al. 1996) y Comamonas acidovorans YM1609 (Kasuya et al. 1997). Las despolimerasas del tipo I y II (Fig. 10) tienen una fibronectina tipo III (Fn3) como dominio de enlace al sustrato, mientras que en las del tipo III, como la P. lemoignei PhaZ5 (isoenzima A) (Jendrossek et al. 1995b) dicho dominio está conformado por una región rica en treonina. Las enzimas de tipo I y II se distinguen por la posición de la caja lipasa en el dominio catalítico; en las del tipo I tal secuencia se sitúa alrededor del centro del dominio catalítico, mientras que en el tipo II se encuentra cercana al extremo N-terminal. Adicionalmente en las del tipo I, el orden de la secuencia de aminoácidos activos es histidina (sitio oxianión)-serina-aspartato-histidina del N terminal al C terminal. En las del tipo II, el orden es serina-aspartato-histidina-histidina (sitio oxianión). Recientemente, se ha propuesto que el dominio de enlace al sustrato tiene una función adicional sobre la modificación de la estructura del polímero (Murase et al. 2002a, b). El dominio de unión podría funcionar como un espaciador que confiere flexibilidad a los dominios de enlace al sustrato y al catalítico, incrementando la eficiencia hidrolítica del dominio catalítico (Nojiri y Saito 1997).

Las despPHA no actúan como lipasas puesto que éstas no pueden enlazarse a las largas cadenas de triacilglicerol e hidrolizarlas. Además éstas presentan como residuo X1 en su sitio activo leucina, mientras que en las lipasas bacterianas este aminoácido es substituido por histidina. Esto sugiere que las lipasas de P. alcaligenes y P. aeruginosa y las despolimerasas de PHA comparten un mecanismo similar de hidrólisis de sustrato (Jaeger et al. 1995). Sin embargo, algunas lipasas pueden degradar películas de P3HB, P4HB, P5HV y P6HHx. Las lipasas de procariontes hidrolizan exclusivamente películas de P3HP mientras que las de eucariontes tienen una amplia especificidad de sustrato y pueden degradar al menos entre 2 a 4 tipos de películas de PHA (Mukai et al. 1993). También existen despP3HB como la de P. funiculosum que no presentan un dominio de enlace de sustrato, que no son específicas y con una débil capacidad de enlace a la superficie del polímero. Este tipo degrada el P3HB, dímeros y trímeros de 3HB. Los residuos catalíticos Ser39, Asp121 y His155 están localizados en una hendidura formada en la superficie de la estructura de la enzima. El interior de esta hendidura sirve como un sitio de enlace al sustrato, el cual es suficientemente amplio para permitir la incorporación de una sola cadena del polímero. El ambiente hidrofóbico al interior de la hendidura enzimática favorece el enlace de las cadenas del polímero, permitiéndole degradar el 100 % de P3HV (Miyazaki et al. 2000). Las interacciones hidrofóbicas desestabilizan las interacciones intra e inter cadena de las cadenas de P3HB, lo cual podría generar un daño mecánico a las regiones ordenadas de las cadenas del polímero, incrementando la movilidad de éstas y creando una región susceptible para la hidrólisis enzimática, resultando en la degradación eficiente de P3HB. El rearreglo espacial de los residuos catalíticos indica que el mecanismo de despolimerización podría ser similar al de una lipasa/serina esterasa (Yamashita et al. 2006). La Ser39 juega un papel central en la reacción catalítica, participando en un ataque nucleofílico sobre el átomo de carbono del carbonilo de la cadena de P3HB, mientras que los residuos His155-Asp121 forman un sistema de puentes de hidrógeno con la cadena de P3HB, mejorando la nucleofilicidad del grupo hidróxilo de la Ser39. El incremento del tamaño del monómero dentro de la cadena del PHA incrementa la longitud de la parte hidrofóbica de la cadena, lo cual podría cubrir preferencialmente o bloquear la hendidura hidrofóbica de la enzima evitando o inhibiendo la degradación. La mayoría de las despolimerasas purificadas son específicas para P3HB y para PHA de cadena corta (desp-scl-PHA). Los microorganismos productores de despolimerasas extracelulares de cadena media (desp-mcl-PHA) son relativamente raros y la mayoría pertenece al género Pseudomonas (Ramsay et al. 1994). Una desp-scl-PHA fue obtenida de P. fluorescens GK123 la que fue capaz de descomponer el 95 y 82 % de poli-3-hidroxioctanoato (P3HO)
y poli-3-hidroxidecanoato-co-3-hidroxioctanoato (P3HDco3HO) después de 38 h y 5 días de cultivo en condiciones aerobias, respectivamente. Esta despolimerasa está constituida por dos cadenas polipeptídicas idénticas con un PM alrededor de 50 kDa con un rango de pH activo de 6.3 a 9.3 (Schirmer et al. 1993). Otra desp-mcl-PHA fue producida recientemente por una nueva bacteria (Pseudomonas sp. RY-1) capaz de crecer en poli-3-hidroxiheptanoato, octanoato, nonanoato y conteniendo otros grupos funcionales (Kim et al. 2000a, b, c). A diferencia de la anterior despolimerasa, ésta tiene cuatro subunidades idénticas, un PM de 115 kDa, un pI de 5.9, una actividad superior al 80% a un rango de pH de 7.0 a 10 y una actividad máxima a pH de 8.5 a una temperatura de 35 °C. Esta despolimerasa fue parcialmente inactivada por EDTA pero no inhibida por los inhibidores usuales de desp-scl-PHA (ditiotreitol y fluoruro de fenilmetil-sulfonilo) (Shiraki et al. 1995, Müller y Jendrossek 1993). Estos resultados sugieren que los residuos de serina, grupos tiol reducidos o puentes disulfuros no son esenciales en el sitio activo de la enzima, en analogía con la despolimerasa de P3HO de P. fluorescens GK13 y deXantomonas sp. JS02 (Kim et al. 2000a, b, c). La importancia de los residuos de serina en la hidrólisis de PHA de cadena media ha sido evidenciado en despolimerasas aisladas y purificadas de Pseudomonas luteola M13-4 (Rhee et al. 2006) y P. alcaligenes LB19 (Kim et al. 2002). En contraste con otras despolimerasas, la correspondiente de P. luteola M13-4 presentó un bajo peso molecular (28 kDa), pI de 6, actividad óptima a pH de 10 a una temperatura de 40 °C. Esta enzima degradó totalmente P3HO mientras que sólo parcialmente el poli-3-hidroxihexanoato y un copolímero de P(3HB-co-60%3HV). La hidrólisis enzimática de PHA primero ocurre en la fase amorfa, seguida por la zona cristalina del polímero. La velocidad de degradación de películas de P3HB y de sus copolímeros ha sido evaluada, encontrando que el incremento de la cristalinidad, el tamaño del cristal y espesor de la película tienen un efecto marcado sobre dicho parámetro (Kasuya et al. 1999). El dominio hidrofóbico de enlace de sustratos de las despP3HB permite la adsorción a la superficie del cristal o película del polímero, incrementando la movilidad y el desorden de las cadenas empaquetadas de P3HB. Esto es necesario para iniciar el ataque enzimático sobre las cadenas del polímero por el dominio catalítico (Sudesh et al. 2000). La velocidad global de degradación se incrementa rápidamente con la concentración y velocidad de ataque de la despP3HB hasta el valor máximo, seguida por una disminución gradual y una variación en la pérdida de peso de la película del polímero, sin un cambio en la cristalinidad total durante la hidrólisis. La degradación enzimática de cristales de P3HB con despP3HB de P. lemoignei (Nobes et al. 1996) y R. picketti T1 (Iwata et al. 1997b, Murase et al. 2001a) fue observada por microscopia de fuerza atómica y de transmisión de electrones, encontrando que la hidrólisis enzimática progresa de las partes terminales cortas de los cristales formando grietas a lo largo del eje longitudinal, mientras que la hidrólisis lateral de los mismos difícilmente ocurre. La tripsina o activador modifica la estructura de la superficie de los gránulos del polímero eliminando proteínas de ésta, permitiéndole a la despolimerasa encontrar y enlazarse a la superficie del polímero. El tercer componente del sistema biocatalítico de PHA es una hidrolasa dimérica responsable de la degradación de los productos de hidrólisis primaria de P3HB (p.e. dímeros y oligómeros de 3HB) a 3HB.
La degradación intracelular es una respuesta endógena de la bacteria que le permite emplear (hidrolizar) sus reservas de carbono acumuladas en forma de PHA a monómeros mediante la participación de despolimerasas intracelulares de PHA (despintPHA).
En el caso del P3HB, éste es hidrolizado a 3HB y a su vez oxidado a acetoacetato por una deshidrogenasa dependiente de NAD. El acetoacetato es convertido a acetoacetil-CoA por la succinato deshidrogenasa. Sin embargo, existe otro mecanismo en Zoogloea ramigera de formación de acetoacetato-CoA mediante una reacción de esterificación catalizada por una CoA-sintetasa usando ATP (Dawes 1988). Las despintPHA son específicas para la forma nativa y amorfa del PHA intracelular. Al respecto, pocos estudios sobre la degradación intracelular de P3HB han sido publicados, por lo que se sabe poco sobre las despolimerasas involucradas. La estrecha asociación de estas enzimas con la superficie de los gránulos conteniendo este polímero y la dependencia de la actividad enzimática con la integridad estructural de los gránulos han dificultado adicionalmente el estudio de las mismas. Las despintPHA han sido menos estudiadas que sus contrapartes extracelulares, permaneciendo algunos mecanismos como el de su regulación aún no muy bien entendidos. Merrick y Doudoroff (1964) fueron los primeros en reportar el sistema intracelular de despolimerasa de P3HB de Rhodospirillum rubrum, el cual fue usado para degradar gránulos nativos de P3HB aislados de Bacillus megaterium. Este sistema está compuesto de un activador termoestable (tripsina) que actúa sobre las proteínas de la capa superficial de los gránulos nativos de P3HB para ser hidrolizados posteriormente por una despolimerasa termolábil. Los sistemas de degradación intracelular de P3HB son frecuentemente muy complejos, como el de Ralstonia eutropha (Uchino et al. 2008), que podría tener hasta nueve despP3HB clasificadas en cuatro clases (PhaZa, b, c y d) (Pohlmann et al. 2006). La PhaZa1 está ampliamente distribuida entre bacterias (York et al. 2003), no contiene el pentapéptido de la caja lipasa (Glu-X1-Ser-X2-Gly) como todas las despolimerasas extracelulares de P3HB (despextP3HB). En esta clase de enzimas la cisteína substituye a la serina en el sitio activo (Kobayashi y Saito 2003). La PhaZb y PhaZc presentan la secuencia de la caja lipasa en sus sitios activos. Estas diferencias estructurales entre los diferentes grupos de despolimerasas determinan en gran medida la especificidad de las mismas por diferentes sustratos. La enzima PhaZa hidroliza P3HB pero no oligómeros lineales o cíclicos de 3HB a diferencia de la PhaZb que lo realiza preferencialmente de manera endo y exo (Scherer et al. 2000). Este tipo de enzima se ha encontrado en el género Wautersia (especies: eutropha y metallidurans). Los principales productos generados por la PhaZ1 a partir de P3HB son oligómeros de 3HB de diferentes longitudes, sobre los cuales actúa la PhaZb, que presenta una amplia especificidad por este tipo de sustratos produciendo los monómeros correspondientes. Estas enzimas actúan sinérgicamente y la localización de ambas en los gránulos de P3HB garantiza una rápida degradación de P3HB in vivo. La primera secuencia de una despolimerasa intracelular (PhaZ1Rue; Ralstonia eutropha H16) fue reportada por Saegusa et al. (2001), la cual pudo degradar P3HB amorfo pero no cristalino. Recientemente, una despintPHA de R. eutropha fue eficientemente purificada, manifestando una actividad hidrolasa del tipo exo y endo sobre oligómeros lineales y cíclicos de 3HB, respectivamente. Las propiedades cinéticas de esta enzima fueron muy similares a las de una oligómero-hidrolasa extracelular de 3HB de R. picketii T1, reportada como una hidrolasa del tipo exo (Kobayashi et al. 2003). La enzima PhaZc tiene un bajo PM (30 kDa) con una alta actividad hidrolasa de oligómeros de 3HB comparada con otras hidrolasas de oligómeros de 3HB. La enzima PhaZc no es ni lipasa ni una esterasa no específica que se ha encontrado en gránulos nativos de P3HB y que degrada P3HB amorfo artificial. La síntesis y degradación de P3HB parece ocurrir simultáneamente (Kawaguchi y Doi 1992), sin embargo, este recambio desde el punto de vista de economía energética parece ser una desventaja para la bacteria. La degradación in vivo de P3HB se inicia con la acción de la despolimerasa PhaZa1 o PhaZb que provoca varias incisiones en las cadenas de las moléculas amorfas de P3HB, dando como resultados oligómeros de 3HB de longitud mediana que permanecen unidos a los gránulos y que mantienen la hidrofobicidad de los mismos. De manera consecuente se origina una descompactación de las mismas cadenas de P3HB y una cantidad pequeña de monómeros de 3HB y oligómeros de 3HB de cadena corta que difunden de los gránulos. La PhaZb degrada de manera exo los oligómeros y cadenas descompactadas de 3HB sobre los gránulos de P3HB a 3HB. Finalmente, la PhaZb localizada en el citosol hidroliza los oligómeros de 3HB difundidos. En contraste con otras despintPHA conocidas, las PhaZ de B. thuringiensis y B. megaterium generan una mayor cantidad de monómeros de 3HB de los gránulos nativos de P3HB, correspondiendo aproximadamente a un 42 y 34 % del total, respectivamente. Estos valores en el caso de la despolimerasa PhaZ7 de Paucimonas lemoignei se encuentran apenas entre el 0.5-2.5 % del total de equivalentes de 3HB presentes en los gránulos de P3HB (Handrick et al. 2001). Se evaluó la degradación intracelular de varias inclusiones de PHA en Hydrogenophaga pseudoflava, incluyendo copolímeros de 3HB, 4HB y de 3HV y de homopolímeros de estos monómeros acumulados separadamente en la célula (Yoon et al. 1995, Yoon y Choi 1999). El copolímero de 3HB/4HB fue degradado cuando el polímero contenía un nivel mínimo de 3HB, mientras que en el caso de los homopolímeros de 3HB (P3HB) ó 4HB (P4HB), únicamente el P3HB fue degradado. Esto muestra la inactividad total de la despolimerasa intracelular contra el P4HB. Estos resultados, aunados a los obtenidos mediante resonancia magnética nuclear de 13C de la degradación de copolímeros conformados con diferentes relaciones molares de 3HB, 4HB y 3HV, han demostrado que la degradación intracelular de PHA es dependiente de la secuencia local del polímero y que el paso de despolimerización es uno de los limitantes en la velocidad de degradación intracelular de PHA. La ve locidad de despolimerización está gobernada por la especificidad de la enzima hacia la secuencia de monómeros, tales como dímeros, trímeros y tetrámeros. La despintPHA de H. pseudoflava no es una enzima que rompa la cadena del polímero de manera secuencial. Además, su incapacidad de degradar P(4HB) y de copolímeros ricos en 4HB, permite concluir que la presencia de un carbono quiral en el grupo éster y la posición gama al carbonilo susceptible a la oxidación, son esenciales para la reacción de hidrólisis por la despolimerasa. Para el caso de P3HB se reportó una velocidad de degradación de primer orden y la comparación de las constantes de velocidad de los diferentes PHA determinaron que la especificidad relativa hacia los sustratos de la despolimerasa es en el orden: 3HB>3HV>4HB. Recientemente, una desp-mcl-PHA de T. thermophilus HB8, diferente a las previamente reportadas en bacterias, ha sido purificada y caracterizada cinéticamente (Papaneophytoua et al. 2011). Este es de los pocos microorganismos capaces de secretar diferentes despolimerasas que degradan P3HB de cadena corta y mediana, dependiendo de las condiciones de crecimiento. La desp-mcl-PHA es secretada en presencia de alcanoatos de cadena media pero no en presencia de glucosa o de alcanoatos de cadena corta. A pesar de que contiene los tres dominios característicos en su estructura, no presenta la secuencia típica del pentapéptido situado en la caja lipasa, puesto que en lugar de tener isoleucina (X1) y Ser (X2) contiene glicina y tirosina, respectivamente. Adicionalmente en el sitio de formación del oxianión durante la hidrólisis del PHA, la histidina es remplazada por asparagina. Esta metaloproteína de bajo PM (28 kDa) tiene un pH óptimo de actividad a 8.5 mientras que su temperatura óptima (70 °C) es superior a las reportadas previamente para despolimerasas de Xantomonas sp. JS02 (60 °C), Streptomyces sp. KJ-72 (50 °C) y P. alcaligenes LB19 (35 °C), debido probablemente a su gran número de aminoácidos cargados (64), que promueven una mayor interacción electrostática entre éstos (puentes de hidrógeno y salinos) estabilizando la estructura de la enzima a altas temperaturas.
Estudios en Pseudomonas oleovorans sobre la síntesis y degradación intracelular de P3HO, demostraron que antes de que la bacteria complete la utilización de la fuente de carbono del medio, la célula disminuye la concentración de polimerasa (de 20 a 12%) mientras que incrementa la producción de la despolimerasa (de 10 al 15%) (Lenz y Marchessault 2005). Este comportamiento parecería representar la respuesta de la célula a un ambiente metabólico cada vez menos permisible (Stuart et al. 1996), preparándose las células a movilizar las reservas intracelulares de carbono para su supervivencia. En Alcaligenes eutropha, la síntesis y degradación de poliésteres puede proceder simultáneamente (Doi et al. 1990). La velocidad de degradación in vitro del ácido polihidroxinonanoico (PHN) en P. oleovorans a pH 9 y 30 °C, ha sido determinada en 0.92 mg/h, siendo muy similar a las reportadas previamente para cuerpos de inclusión de P3HO y poli-3-hidroxi-5-fenilvalerato (PHPV) (0.98 mg/h). Sin embargo, en inclusiones conteniendo mezclas de PHN/PHPV la velocidad de degradación fue muy baja (0.30 mg/h), posiblemente por la distribución inusual de los polímeros dentro de los cuerpos de inclusión. La actividad de despolimerasas intracelulares en estudios in vitro contrasta con aquellas obtenidas en pruebas de degradación in vivo de inclusiones de P3HN, PHPV y P3HV/ PHPV. En estos casos, la degradación in vivo de P3HN procedió significativamente más rápido que la de su contraparte para PHPV. Además de que, la degradación de P3HN se lleva a cabo preferencialmente cuando se encuentra en las inclusiones con PHPV. Entre las razones reportadas para explicar la diferencia entre las velocidades de degradación in vitro e in vivo, se encuentran, primero que la velocidad de degradación in vivo pudiera ser limitada respecto a la in vitro debido a la variedad de enzimas o a la falta de especificidad por el sustrato para la utilización del polímero que se puede presentar in vivo, comparado con una sola despolimerasa que se utiliza en los ensayos in vitro. Una segunda razón, podría ser que los estudios in vitro se realizan a las condiciones óptimas determinadas y requeridas por la(s) enzima(s), lo cual no necesariamente ocurre in vivo. En este sentido, estudios in vitro demostraron que la despolimerasa intracelular de P. oleovorans fácilmente degradó P3HN y PHPV, mientras que in vivo, el P3HN fue degradado preferencialmente. Esto sugiere que en este microorganismo otros procesos en el metabolismo de PHA limitan la velocidad de degradación de PHPV in vivo. La presencia del grupo aromático en las unidades del polímero posiblemente es el factor limitante, tal y como lo han sugerido otros estudios empleando PHPV y despolimerasas extracelulares de P. maculicola (Foster et al. 1995). El sistema de despolimerización intracelular de PHA de cadena mediana aún no ha sido bien elucidado en comparación con la información disponible de los genes que codifican para las enzimas intracelulares que degradan los PHA de cadena corta. En los últimos años se ha caracterizado el gene de la despolimerasa phaZ de P. putida KT2442, que es capaz de hidrolizar específicamente PHA de cadena media conteniendo monómeros alifáticos y aromáticos. Esta despolimerasa, localizada en los gránulos de PHA, se comporta como una serina hidrolasa y degrada tales polímeros de manera endo y exo (de Eugenio et al. 2007, 2008). La degradación intracelular de PHA acumulado con cierta heterogeneidad estructural en P. putida y P. citronellolis (Chung et al. 2001) fue catalogada como una cinética de primer orden, con dos diferentes constantes de velocidad (k1) una de 0.087 y otra de 0.015 h-1 durante las primeras 20 h de degradación. En la última etapa de degradación, la velocidad de desaparición de las unidades de 3HO fue estimada en 0.020 h-1 y comparable a la de HPV (de 0.015 h-1). Inicialmente una más alta desaparición de 3HO se observa debido a la degradación de P3HO, mientras que en la última fase ésta fue más lenta como consecuencia de la degradación de las unidades de 3HO incorporadas con las de HPV en las cadenas del copolímero. En contraste, los valores de k1 para el PHA de P. citronellolis fueron de 0.035 y 0.029 h-1 para las unidades de 3HO y HPV, respectivamente; indicando esta similitud de velocidades la naturaleza al azar del copolímero del PHA de P. citronellolis. Adicionalmente, la velocidad de degradación de P3HO respecto a la de PHPV por la despolimerasa intracelular de P. putida indica que ésta es más específica para el PHA alifático que para el aromático. La evaluación de la cinética de adsorción e hidrólisis de la despextP3HB de A. faecalis sobre la superficie de cinco tipos de películas de PHA, permitió poner en evidencia que las películas de P3HB, P3HP y P4HB fueron hidrolizadas por la enzima, en tanto que las correspondientes de P(S-2-hidroxipropionato) y P6HHx no lo fueron. La despP3HB se adsorbió sobre todas las películas siguiendo una cinética descrita por la isoterma de Langmuir (Kasuya et al. 1996). Esto evidencia que el dominio de enlace de la despolimerasa no es específico para las películas de PHA mientras que el sitio activo es específico para la hidrólisis de PHA. Debido a la escasa información de las estructuras terciarias de despP3HB, no se conoce específicamente cuales aminoácidos del dominio de enlace de la enzima están involucrados en la adsorción a la superficie del polímero y como éstos contribuyen en tal proceso. Últimamente, mediante estudios de mutagénesis aleatoria y de microscopía de fuerza atómica se pudo identificar los aminoácidos involucrados en la reacción, así como la fuerza de adhesión (100 pN) implicada en la adsorción al P3HB de la despP3HB (PhaZRpiT1) de Ralstonia pickettii T1. Entre los aminoácidos del dominio de enlace de la enzima, la Ser, Val, Leu, Ala y Tyr probab lemente participan en la adsorción de la enzima a la superficie del P3HB mientras que la Leu441, Tyr443 y Ser445 podrían estar directamente implicadas en la adsorción (Hirashi et al. 2010). El análisis sugiere que la alta hidrofobicidad de estas tres posiciones mejora la capacidad de adsorción de la enzima.
En general, los PHA son insolubles en agua y pueden ser degradados a una velocidad moderada (3-9 meses) por muchos microorganismos. La evaluación y monitoreo de la biodegradación de PHA incluye: observación visual (rugosidad de la superficie, formación de orificios o grietas, fragmentación, cambios en color o formación de biopelículas sobre la superficie); microscopía electrónica de barrido (SEM) o de fuerza atómica (AFM); cambios en las propiedades mecánicas y masa molar (Erlandsson et al. 1997); medición de la pérdida de peso y el empleo de la técnica de formación de zona clara (Nishida y Tokiwa 1993, Abou-Zeid 2001). La mayoría de las bacterias y hongos degradadores de polímeros en cultivos puros hasta ahora son inicialmente aislados sobre cajas Petri con esta técnica. La respirometría (producción o evolución de CO2/ consumo de O2) (Püchner et al. 1995) o exclusivamente la formación de CO2 (prueba de Sturm) es el método más frecuentemente empleado para medir la biodegradación en pruebas de laboratorio, obteniendo información directa sobre la bioconversión del esqueleto carbonado del polímero al producto final metabólico (Pagga et al. 2001). Una manera rápida de evaluar la capacidad de degradación de PHA es mediante el empleo de medios sólidos a base de agar y gránulos desnaturalizados del polímero (método de zona clara) o considerando una película delgada de dicho material asociada con un colorante (p.e. rojo Sudán) para el caso de poner en evidencia despolimerasas de PHA de cadena corta o de cadena media, respectivamente. De manera paralela se han desarrollado emulsiones de PHA y surfactantes, estables y resistentes al calor para la rápida identificación y aislamiento de microorganismos degradadores de PHA (Ramsay et al. 1994, Horowitz et al. 1994 y 1995, Marchessault et al. 1995, Schirmer et al. 1995). Las bacterias con rápido crecimiento y altas velocidades de hidrólisis de los polímeros pueden ser diferenciadas de aquellas con bajas capacidades de hidrólisis de PHA por la medición del diámetro de las colonias y de las zonas claras (halos) generadas alrededor de las mismas.
En la actualidad, los polímeros residuales tales como los biopoliésteres han tenido mucha atención como fuentes concentradas de monómeros, lo que ha permitido la reducción de costos de producción y de la emisión de dióxido de carbono (Tokiwa y Jarerat 2004). El reciclado químico de estos materiales es un proceso rápido que convencionalmente involucra degradaciones térmicas e hidrolíticas como primer paso para la generación de sustratos monoméricos u oligoméricos a partir de poliésteres. Sin embargo, estos métodos producen subproductos indeseables, tales como racematos y compuestos tipo crotonato (Fan et al. 2003, Saeki et al. 2005, Abe 2006). Una posible solución a este problema es la aplicación de enzimas, tales como las despP3HB para el reciclado de P3HB, debido a las ventajas que presentan. Entre éstas podemos destacar, su alta especificidad que evita la formación de productos indeseables, su capacidad de funcionar bajo condiciones moderadas y su formación a partir de fuentes renovables (Schulze y Wubbolts 1999, Ran et al. 2008).
CONCLUSIONES
Los PHA son plásticos microbianos biodegradables y biocompatibles que se obtienen a partir de fuentes renovables. Actualmente se producen a escala industrial usando principalmente bacterias silvestres, aunque también se está estudiando su obtención con microorganismos recombinantes, y por vía enzimática. Sus propiedades físicas dependen en gran medida de su estructura química, siendo los PHA de cadena corta los más estudiados, en especial el P3HB, mientras que la investigación de los PHA de cadena media aún representa un gran campo de de estudio. Con base en las propiedades físicas de los PHA que se conocen hasta el momento, estos podrían utilizarse como sustitutos de algunos plásticos derivados del petróleo, como el polipropileno y el polietileno y su uso ha sido enfocado como materia prima para fabricar productos desechables biodegradables. Sin embargo, la principal desventaja para utilizarlos extensivamente es su precio, el cual resulta superior al de los plásticos derivados del petróleo. No obstante, en los últimos años se han ido ampliando y popularizando sus aplicaciones en áreas más especializadas, como la biomédica y la farmacéutica, en donde la biocompatibilidad y funcionalidad de estos biopolímeros justifican su costo.
Las tendencias actuales en cuanto a la producción de PHA por microorganismos se enfocan en dos puntos centrales:
a) Lograr que el proceso de producción sea competitivo económicamente y sea ambientalmente amigable. En este sentido, los trabajos de investigación se han focalizado en la optimización de procesos de fermentación para aumentar su productividad, obteniendo cultivos de alta densidad y con alto contenido de polímero; también se ha buscado mejorar los procesos de extracción y purificación del PHA para alcanzar mayores eficiencias de recuperación, minimizando el uso de solventes tóxicos. Asimismo, se ha investigado sobre el uso de fuentes de carbono económicas, renovables y altamente disponibles, con el fin de disminuir los costos de producción. En este sentido el glicerol derivado de la producción de biodiesel, los hidrolizados de residuos celulósicos y diversos efluentes agroindustriales, son sustratos con gran potencial para este fin. En México, en particular sería interesante utilizar como fuente de carbono residuos de la industria tequilera y cervecera, así como subproductos y residuos del cultivo de maíz y de los procesos industriales basados en este grano. De manera paralela se están buscando nuevos microorganismos, capaces de acumular grandes cantidades de polímero, que presenten un crecimiento rápido bajo diversas condiciones ambientales y utilicen fuentes de carbono económicas; así como el estudio del metabolismo de microorganismos ya conocidos, para llevar a cabo modificaciones genéticas exitosas que den lugar a cepas superproductoras.
b) Obtener PHA con propiedades físicas nuevas o mejoradas que permitan ampliar su uso en aplicaciones de alto valor agregado, mediante la búsqueda nuevos microorganismos silvestres o modificados genéticamente, capaces de sintetizar copolímeros con estructura y peso molecular deseable. Una segunda estrategia considerada es la modificación química o enzimática del polímero previamente producido y extraído, con el fin de lograr las características deseables del mismo.
En cuanto a la biodegradación de estos materiales, aún faltan estudios que permitan comprender mejor los mecanismos y la importancia de los microorganismos implicados en el proceso de degradación, especialmente de los PHA producidos mediante la incorporación de algunos precursores complejos, modificados químicamente o mezclados con otros compuestos, ya que estos factores podrían afectar su biodegradabilidad. Adicionalmente, una línea emergente de investigación con un amplio potencial en esta área es el estudio y caracterización de las enzimas despolimerasas para aplicarlas en la degradación controlada de PHA, con el fin de obtener oligómeros y monómeros con potencial de uso en aplicaciones especializadas.
En Latinoamérica el único país que produce comercialmente estos bioplásticos es Brasil, específicamente la empresa Biocycle que utiliza como fuente de carbono azúcar de caña. En el caso de México, la investigación sobre PHA por los centros de investigación y universidades existentes es escaso. Sin embargo en nuestro país existe una gran área de oportunidad en el estudio de diferentes residuos agroindustriales como materia prima para la biosíntesis de PHA. Por otro lado, los países latinoamericanos y en especial México, tienen una enorme diversidad biológica la cual no ha sido explotada para la búsqueda de nuevos microorganismos, que produzcan PHA más eficientemente, o que sinteticen polímeros con propiedades novedosas, lo cual es también un área de oportunidad para la investigación científica.
REFERENCIAS
Abe H. (2006). Thermal degradation of environmentally degradable poly(hydroxyalkanoic acid)s. Macromol. Biosci. 6, 469-486. [ Links ]
Abou-Zeid D.M. (2000). Anaerobic Biodegradation of Natural and Synthetic Polyesters. PhD thesis. Technical University Braunschweig. Alemania, 127 pp. [ Links ]
Amirul A.A., Yahya A.R.M., Sudesh K., Azizan M. y Majid M.I.A. (2008). Biosynthesis of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) copolymer by Cupriavidus sp. USMAA1020 isolated from Lake Kulim, Malaysia. Bioresource Technol. 99,4903-4909. [ Links ]
Anderson A.J. y Dawes E.A. (1990). Occurrence, metabolism, metabolic role, and industrial uses of bacterial polyhydroxyalkanoates. Microbiol. Rev. 54,450-472. [ Links ]
Andreessen B.y Steinbüchel A. (2010). Biosynthesis and biodegradation of 3-hydroxypropionate- containing polyesters. Appl. Environ. Microbiol. 76, 4919-4925. [ Links ]
Aneja P. y Charles T. C. (1999). Poly-3-hydroxybutyrate degradation in Rhizobium (Sinorhizobium) meliloti: isolation and characterization of a gene encoding 3-hydroxybutyrate dehydrogenase, J. Bacteriol. 181, 849-857. [ Links ]
Asano Y. y Watanabe S. (2001). Isolation of poly (3-hydroxybutyrate) (P3HB)-degrading microorganisms and characterization of P3HBdepolymerase from Arthrobacter sp. strain W6. Biosci. Biotechnol. Biochem. 65, 1191-1194. [ Links ]
Atlić A., Koller M., Scherzer D., Kutschera C., Grillo-Fernandes E., Horvat P., Chiellini E. y Braunegg G. (2011). Continuous production of poly((R)-3-hydroxybutyrate) by Cupriavidus necator in a multistage bioreactor cascade. Appl. Microbiol. Biotechnol. 91,295-304. [ Links ]
Babel W. y Steinbüchel A. (2001) Biopolyesters. Special issue of advances in Biochem. Eng. Biotechnology. 1st. ed. Springer-Verlag. Berlin, Alemania, 342 pp. [ Links ]
Bachmann B.y Seebach D. (1999). Investigation of the enzymatic cleavage of diastereomeric oligo(3-hy-droxybutanoates) containing two to eight HB units. A model for the stereoselectivity of P3HB depolymerase from Alcaligenes faecalis T1. Macromolecules 32, 1777-1784. [ Links ]
Bengtsson S., Werker A. y Welander T. (2008). Production of polyhydroxyalkanoates by glycogen accumulating organisms treating a paper mill wastewater. Wat. Sci. Technol. 58, 323-330. [ Links ]
Bertrand J.L., Ramsay B.A., Ramsay J.A. y Chavarie C. (1990). Biosynthesis of poly-B-hydroxyalkanotes from pentoses by Pseudomonas pseudoflava. Appl. Environ. Microbiol. 56, 3133-3138. [ Links ]
Borah B., Thakur P.S. y Nigam J.N. (2002). The influence of nutritional and environmental conditions on the accumulation of poly-beta-hydroxybutyrate in Bacillus mycoides RLJ B-017. J. Appl. Microbiol. 92, 776-783. [ Links ]
Bormann E.J. y Roth M. (1999). the Production of polyhydroxybutyrate by Methylobacterium rhodesianum and Ralstonia eutropha in media containing glycerol and casein hydrolysates. Biotechnol. Lett. 21, 1059-1063. [ Links ]
Bramer C.O., Vandamme P., da Silva L.F., Gomez J.G.C. y Steinbüchel A. (2001). Burkholderia sacchari sp nov., a polyhydroxyalkanoate-accumulating bacterium isolated from soil of a sugar-cane plantation in Brazil. Int. J. Sys. Evol. Micr. 51, 1709-1713. [ Links ]
Brandhl H., Bachofen R., Mayer J. y Wintermantel E. (1995). Degradation and applications of polyhydroxyalkanoates. Can. J. Microbiol. 41, 143-153. [ Links ]
Brandl H., Gross R.A., Lenz R.W. y Fuller R.C. (1988). Pseudomonas oleovorans as a source of poly(beta-Hydroxyalkanoates) for potential applications as biodegradable polyesters. Appl. Environ. Microbiol. 54, 1977-1982. [ Links ]
Braunegg G., Lefebvre G. y Genser K.F. (1998). Polyhydroxyalkanoates, Biopolyesters from renewable resources: Physiological and engineering aspects. J. Biotechnol. 65, 127-161. [ Links ]
Brucato C.L. y Wong S.S. (1991). Extracellular poly(3-hydroxybutyrate) depolymerase from Penicillium funiculosum: general characteristics and active site studies. Arch. Biochem. Biophys. 290, 497-502. [ Links ]
Byrom D. (1992). Production of poly-beta-hydroxybutyrate - poly-beta-hydroxyvalerate copolymers. Fems Microbiol. Rev. 103, 247-250. [ Links ]
Byrom D. (1993). The synthesis and biodegradation of polyhydroxyalkanoates from bacteria. Int. Bioterio. Biodegrad. 31,199-208. [ Links ]
Cai M., Chua H., Wong A., Yu H., Sin N., Ren J., He D. y Zhao Q. (2008). Polyhydroxyalkanoates microbiological synthesis from food wastes. Huan Jing Ke Xue 29, 2643-2648. [ Links ]
Calabia B.P.y Tokiwa Y.(2004). Microbial degradation of poly(D-3hydroxybutyrate) by a new thermophilic Streptomyces isolate. Biotechnol. Lett. 26, 15-19. [ Links ]
Cao A., Arai Y., Yoshie N., Kasuya K.I., Doi Y. y Inoue Y. (1999). Solid structure and biodegradation of the compositionally fractionated poly(3hydroxybutyric acid- co-3-hydroxypropionic acid)s. Polymer 40, 6821-6830. [ Links ]
Chen G. (2010). Plastics from Bacteria: Natural Functions and Applications. 1a. ed. Springer-Verlag. 450 pp. [ Links ]
Chen J., Zhang L., Chen J. y Chen G. (2007). Biosynthesis and characterization of polyhydroxyalkanoate copolyesters in Ralstonia eutropha P3HB—4 harboring a low-substrate-specificity PHA synthase PhaC2Ps from Pseudomonas stutzeri 1317. Chin. J. Chem. Eng. 15, 391-396 . [ Links ]
Choi J.I. y Lee S.Y. (1999). Efficient and economical recovery of poly(3-hydroxybutyrate) from recombinant Escherichia coli by simple digestion with chemicals. 62, 546-553. [ Links ]
Chung D.M., Choi M.H., Song J.J., Yoon S.C., Kang I.K. y Huh N.E. (2001). Intracellular degradation of two structurally different polyhydroxyalkanoic acids accumulated in Pseudomonas putida and Pseudomonas citronellolis from mixtures of octanoic acid and 5-phenylvaleric acid. Int. J. Biol. Macromol. 29, 243-250. [ Links ]
Çolak A., Şişik D., Saglam N., Güner S., Çanakçi S. y Beldüz A.O. (2005). Characterization of a thermoalka-lophilic esterase from a novelnthermophilic bacterium, Anoxybacillusgonensis G2. Bioresource Technol. 96, 625-631. [ Links ]
Costa S.G., Lépine F., Milot S., Déziel E., Nitschke M. y Contiero J. (2009). Cassava wastewater as a substrate for the simultaneous production of rhamnolipids and polyhydroxyalkanoates by Pseudomonas aeruginosa. J. Ind. Microbiol. Biotechnol. 3, 1063-1072. [ Links ]
da Cruz Pradella J.G., Taciro M.K. y Pataquiva Mateus A.Y. (2010). High-cell-density poly (3-hydroxybutyrate) production from sucrose using Burkholderia sacchari culture in airlift bioreactor. Bioresource Technol. 101, 8355-8360 . [ Links ]
Dawes E.A. (1988). Polyhydroxybutyrate: an intriguing biopolymer. Biosci. Rep. 8, 537-547. [ Links ]
de Eugenio L.I., García J.L., García P., Prieto M.A. y Sanz J.M. (2008). Comparative analysis of the physiological and structural properties of a medium chain length polyhydroxyalkanoate depolymerase from Pseudomonas putida KT2442. Eng. Life. Sci. 8, 260-267. [ Links ]
de Eugenio L.I., García P., Luengo J.M., Sanz J.M., San Roman J., García J.L. y Prieto M.A. (2007). Biochemical evidence that phaZ gene encodes a specific intracellular medium chain length polyhydroxyalkanoate depolymerase in Pseudomonas putida KT2442. J. Biol. Chem. 282, 4951-4962. [ Links ]
Delafield F.P., Doudoroff M., Palleroni N.J., Lusty C.J. y Contopoulos R. (1965). Decomposition of poly-b-hydroxybutyrate by pseudomonads. J. Bacteriol. 90, 1455-1466. [ Links ]
Doi Y., Segawa A. y Kunioka M. (1990). Biosynthesis and characterization of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) in Alcaligenes eutrophus. Int. J. Biol. Macr. 12, 106-11. [ Links ]
Doi Y., Tamaki A., Kunioka M. y Soga K. (1987). Biosynthesis of terpolyesters of 3-hydroxybutyrate, 3-hydroxyvalerate, and 5-hydroxyvalerate in Alcaligenes eutrophus from 5-chloropentanoic and pentanoic acids. Makromol. Chem. Rap. Com. 8,631-635. [ Links ]
Eggink G., Dewaard P. y Huijberts G.N.M. (1995. Formation of novel poly(hydroxyalkanoates) from long-chain fatty-acids. Can. J. Microbiol. 41, 14-21. [ Links ]
Elbanna K., Lütke-Eversloh T., Jendrossek D., Luft-mann H. y Steinbüchel A. (2004). Studies on the biodegradability of polythioester copolymers and homopolymers by polyhydroxyalkanoate (PHA)-degrading bacteria and PHA depolymerases. Arch. Microbiol. 182, 212-225. [ Links ]
Erlandsson B., Karlsson S. y Albertsson A.C. (1997). The mode of action of corn starch and prooxidant system in LDPE: influence of themooxidation and UV-irradiation on the molecular weight changes. Poly. Degrad. Stab. 55, 237-245. [ Links ]
Fan Y., Nishida H., Shirai Y. y Endo T. (2003). Control of racemization for feedstock recycling of PLLA. Green Chem. 5, 575-579. [ Links ]
Fiorese M.L., Freitas F., Pais J., Ramos A.M., Aragao G. y Reis M. (2009). Recovery of polyhydroxybutyrate (P3HB) from Cupriavidus necator biomass by solvent extraction with 1,2-propylene carbonate. Eng. Life. Sci. 9,454-461. [ Links ]
Foster L.J.R., Zervas S.J., Lenz R.W. y Fuller R.C. (1995). The biodegradation of poly-3-hydroxyalkanoates, PHAs, with long alkyl substituents by Pseudomonas maculicola. Biodegradation 6, 67-73. [ Links ]
Fuchtenbusch B., Fabritius D. y Steinbüchel A. (1996). Incorporation of 2-methyl-3-hydroxybutyric acid into polyhydroxyalkanoic acids by axenic cultures in defined media. Fems. Microbiol. Lett. 138, 153-160. [ Links ]
Fuchtenbusch B. y Steinbüchel A. (1999). Biosynthesis of polyhydroxyalkanoates from low-rank coal liquefaction products by Pseudomonas oleovorans and Rhodococcus ruber. Appl. Microbiol. Biotechnol. 52, 91-95. [ Links ]
Fukui T., Narikawa T., Miwa T., Shirakawa Y., Saito T. y Tomita K. (1998). Effect of limited tryptic modification of a bacterial poly(3-hydroxybutyrate) depolymerase on its catalytic activity. Biochem. Biophys. Acta 952, 164-171. [ Links ]
Garcia-Lillo J. y Rodriguez-Varela F. (1990). Effects of culture conditions on poly(β-hydroxybutyric acid) production by Haloferax mediterranei. Appl. Env. Microbiol. 56, 2517-2521. [ Links ]
Gonda K.E., Jendrossek D. y Molitoris H.P. (2000). Fungal degradation of thermoplastic polymers under simulated deep-sea conditions. Hydrobiologia 426, 173-183. [ Links ]
González-García Y., Nungaray J., Córdova J., González-Reynoso O,. Koller M., Atlic A. y Braunegg G. (2008). Biosynthesis and characterization of polyhydroxyalkanoates in the polysaccharide-degrading marine bacterium Saccharophagus degradans ATCC 43961. J. Ind. Microbiol. Biotechnol. 35, 629-633. [ Links ]
González-López J., Pozo C., Martínez-Toledo M.V., Rodelas B. y Salmeron V. (1996). Production of polyhydroxyalkanoates by Azotobacter chroococcum H23 in wastewater from olive oil mills (alpechín). Int. Biodet. Biodegrad. 38, 271-276. [ Links ]
Greenspan P. y Fowler S.D. ( 1985 ). Spectrofluorometric studies of the lipid probe, nile red. J. Lip. Res. 26, 781-789. [ Links ]
Grothe E., Moo-Young M. y Chisti Y. (1999). Fermentation optimization for the production of poly(beta-hydroxybutyric acid) microbial thermoplastic. Enzyme Microb. Tech. 25, 132-141 [ Links ]
Guerin P., Renard E.y Langlois V. (2010). Degradation of natural and artificial poly((R)-3-hydroxyalkanoate)s: from biodegradation to hydrolysis. En: Plastics from Bacteria: Natural Functions and Applications (G. G.-Q. Chen, Ed.). Microbiol. Monogr. Springer-Verlag, p. 283-321. [ Links ]
Hafuka A., Sakaida K., Satoh H., Takahashi M., Watanabe Y. y Okabe S. (2011). Effect of feeding regimens on polyhydroxybutyrate production from food wastes by Cupriavidus necator. Bioresource Technol. 102, 3551-3553. [ Links ]
Hahn S.K., Chang Y.K., Kim B.S. y Chang H.N. (1994). Optimization of microbial poly(3-hydroxybutyrate) recovery using dispersions of sodium-hypochlorite solution and chloroform. Biotechnol. Bioeng. 44, 256-261. [ Links ]
Han R.J., Son Y.J., Chang C.S. y Kim M.N. (1998). Purification and properties of extracellular poly (3-hydroxybutyrate) depolymerase produced by Penicillium pinophilum. The Journal of Microbiology 36, 67-73. [ Links ]
Handrick R., Reichart S., Schultheiss D., Reichart T., Schüler D., Jendrossek V. y Jendrossek D. (2004). Unraveling the function of the Rhodospirillum rubrum activator of polyhydroxybutyrate (P3HB) degradation, the activator is a P3HB-granule-bound protein (pha-sin). J. Bacteriol. 186, 2466-2475. [ Links ]
Handrick R., Reinhardt S., Focarete M.L., Scandola M., Adamus G., Kowalczuk M. y Jendrossek D. (2001). A new type of thermoalkalophilic hydrolase of Paucimonas lemoignei with high specificity for amorphous polyesters of short chain-length hydroxyalkanoic acids. J. Biol. Chem. 276, 36215-36224. [ Links ]
Handrick R., Technow U., Reichart T., Reinhardt S., Sander T. y Jendrossek D. (2004b). The activator of the Rhodospirillum rubrum P3HB depolymerase is a polypeptide that is extremely resistant to high temperature (121°C) and other physical or chemical stresses. FEMS Microbiol. Lett. 230, 265-274. [ Links ]
Hesselmann R.P., Fleischmann T., Hany R. y Zehnder A.J. (1999). Determination of polyhydroxyalkanoates in activated sludge by ion chromatographic and enzymatic methods. J. Microbiol. Meth. 35, 111-119. [ Links ]
Hiraishi T., Komiya N. y Maeda M. (2010). Y443F mutation in the substrate-binding domain of extracellular P3HB depolymerase enhances its P3HB adsorption and disruption abilities. Poly. Degrad. Stab. 95, 1370-1374. [ Links ]
Hong K., Sun S., Tian W., Chen G.Q. y Huang W. (1999). A rapid method for detecting bacterial polyhydroxyalkanoates in intact cells by Fourier transform infrared spectroscopy. Appl. Microbiol. Biotechnol. 51, 523-526. [ Links ]
Horowitz D. M. y Sanders J. K. M. (1995). Biomimetic amorphous granules of polyhydroxyalkanoates: composition, mobility, and stabilization in vitro by proteins. Can. J. Microbiol. 41, 115-123. [ Links ]
Horowitz D. M. y Sanders J. K. M. (1994). Amorphous, biomimetic granules of polyhydroxybutyrate: preparation, characterization, and biological implications. J. Am. Chem. Soc. 116, 2695-2702. [ Links ]
Hrabak O. (1992). Industrial-production of poly-beta-hydroxybutyrate. Fems Microbiol. Rev. 103, 251-255. [ Links ]
Huang D.H., Lu X.Z., Xiong S.J. y Hao L.P. (2006). Microorganism screening for PHAMCL accumulation from activated sludge and its characteristics. J. Agro-Environ. Sci. 25, 517-521 (Chinese). [ Links ]
Huang T.Y., Duan K.J., Huang S.Y. y Chen C.W. (2006). Production of polyhydroxyalkanoates from inexpensive extruded rice bran and starch by Haloferax mediterranei. J. Ind. Microbiol. Biotechnol. 33, 701-706. [ Links ]
Isemori Y., Tajima K., Tanaka S., Yu F., Ishida K. y Inoue Y. (2006). Effects of pH of fermentation medium on biosynthesis of poly((3-hydroxybutyrate)-co-(3-mercaptopropionate)) by Wautersia eutropha. Macromol. Biosci. 6, 818-826. [ Links ]
Iwata T., Doi Y., Tanaka T., Akehata T., Shiromo M. y Teramachi S. (1997). Enzymatic degradation and adsorption on poly((R)-3-hydroxybutyrate) single crystals with two types of extracellular P3HB depolymerases from Comamonas acidovorans YM1609 and Alcaligenes faecalis T1. Macromolecules 30, 5290-5296. [ Links ]
Jacob G., Garbow J.R. y Schaeferg J. (1986). Direct Measurement of poly(beta-hydroxybutyrate) in a pseudomonad by solid-state 13C NMR. J. Biol. Chem. 261, 16785-16787. [ Links ]
Jaeger K.E., Ransac S., Dijkstra B.W., Colson C., van Heuvel M. y Misset O. (1994). Bacterial lipases. FEMS Microbiol. Rev. 15, 29-63. [ Links ]
Jansen P.H. y Harfoot C.G. (1990). Ilyobacter delafieldii sp. nov., a metabolically restricted anaerobic bacterium fermenting P3HB. Arch. Microbiol. 154, 253-259. [ Links ]
Jendrossek D., Knoke I., Habibian R.B., Steinbüchel A.y Schlegel H.G. (1993). Degradation of poly(3-hydroxybutyrate), P3HB, by bacteria and purification of a novel P3HB depolymerase from Comamonas sp. J. Environ. Polymer Degrad. 1 , 53-63. [ Links ]
Jendrossek, D. (2001). Microbial degradation of polyesters. En: Biopolyesters (Steinbüchel A. y Babel W. Eds.). Adv. Biochem. Eng. Biot. 71, 293-325. [ Links ]
Jendrossek D. y Handrick R. (2002). Microbial degradation of polyhydroxyalkanoates. Annu. Rev. Microbiol. 56, 403-432. [ Links ]
Jendrossek D., Schirmer A. y Schlege H.G. (1996). Biodegradation of polyhydroxyalkanoic acids. Appl. Microbiol. Biotechnol. 46, 451-463. [ Links ]
Jendrossek D., Backhaus M. y Andermann M. (1995). Characterization of the extracellular poly(3-hydroxy-butyrate) depolymerase of Comamonas sp. and of its structural gene. Can. J. Microbiol. 41, 160-169. [ Links ]
Jendrossek D., Frisse A., Behrends A., Andermann M., Kratzin H.D., Stanislawski T. y Schlegel H. G. (1995b) Biochemical and molecular characterization of the Pseudomonas lemoignei polyhydroxyalkanoate depolymerase system. J. Bacteriol. 177, 596-607. [ Links ]
Kasuya K., Doi Y. y Yao T. (1994). Enzymatic degradation of poly((R)-3hydroxybutyrate) by Comamonas testosteroni ATSU of soil bacterium. Polym. Degrad. Stab. 45, 379-386. [ Links ]
Kasuya K., Ohura T., Masuda K. y Doi Y. (1999). Substrate and binding specificities of bacterial polyhydroxybutyrate depolymerases. Int. J. Biol. Macromol. 24, 329-336. [ Links ]
Kasuya, K., Inoue Y., Tanaka T., Akehata T., Iwata T., Fukui T. y Doi Y. (1997). Biochemical and molecular characterization of the polyhydroxybutyrate depolymerase of Comamonas acidovorans YM1609, isolated from freshwater. Appl. Envir. Microbiol. 63, 4844-4852. [ Links ]
Kasuya K., Inoue Y. y Doi Y. (1996). Adsorption kinetics of bacterial P3HB depolym surface of polyhydroxyalkanoate films. Int. J . Biol. Macromol. 19, 35-40. [ Links ]
Kawaguchi Y. y Doi Y. (1992). Kinetic and mechanism of synthesis and degradation of poly(3hydroxybutyrate) in Alcaligenes eutrophus. Macromolecules 25, 2324-2329. [ Links ]
Keenan T.M., Nakas J.P. y Tanenbaum S.W. (2006). Polyhydroxyalkanoate copolymers from forest biomass. J. Ind. Microbiol. Biotechnol. 33, 616-626. [ Links ]
Khanna S. y Srivastava A.K. (2005). A simple structured mathematical model for biopolymer (P3HB) production. Biotechnol. Prog. 21, 830-838. [ Links ]
Khanna S. y Srivastava A.K. (2005b). Recent advances in microbial polyhydroxyalkanoates. Process Biochem. 40, 607-619. [ Links ]
Khardenavis A.A., Kumar M.S., Mudliar S.N. y Chakrabarti (2007). Biotechnological conversion of agro-industrial wastewaters into biodegradable plastic, poly β-hydroxybutyrate Bioresource Technol. 98, 3579-3584. [ Links ]
Kim B.S. y Chang H.N. (1995). Control of glucose feeding using exit gas data and its application to the production of P3HB from tapioca hydrolysate by Alcaligenes eutrophus. Biotechnol. Tech. 9, 311-314. [ Links ]
Kim B.S. y Chang H.N. (1998). Production of poly(3-hydroxybutyrate) from starch by Azotobacter chroococcum. Biotechnol. Lett. 20, 109-112. [ Links ]
Kim B.S., Lee S.C., Lee S.Y., Chang H.N., Chang Y.K. y Woo S.I. (1994). Production of polyhydroxybutyrate by fed batch with glucose concentration control in Ralstonia eutropha. Biotechnol. Bioeng. 43, 892-898. [ Links ]
Kim B.S., Lee S.C., Lee S.Y., Chang H.N., Chang Y.K. y Woo S.I. (1994b). Production of poly(3-hydroxybutyric-co-3-hydroxyvaleric acid) by fed-batch culture of Alcaligenes eutrophus with substrate control using on-line glucose analyzer. Enzyme Microbiol. Technol. 16, 556-561. [ Links ]
Kim D.Y., Nam J.S., Rhee Y.H. (2002). Characterization of an extracellular medium-chain-length poly(3-hydroxyalkanoate) depolymerase from Pseudomonas alcaligenes LB19. Biomacromolecules 3, 291-296. [ Links ]
Kim D.Y., Shin D.S., Nam J.S., Rhee Y.H. (2002b). Molecular cloning of the gene encoding medium-chain-length poly(3-hydroxyalkanoate) depolymerase from Pseudomonas alcaligenes LB19. Proc. Int. Meet. Fed. Korean Microbiol. Soc. p.206. [ Links ]
Kim D.Y., Yun J.H., Kim H.W., Bae K.S., Rhee Y.H. (2002c). Purification and characterization of poly(3-hydroxybutyrate) depolymerase from a fungal isolate, Emericellopsis minima W2. J. Microbiol. 40, 129-133. [ Links ]
Kim H., Ju H.S. y Kim J. (2000). Characterization of an extracellular poly(3-hydroxy-5-phenylvalerate) depolymerase from Xanthomonas sp. JS02. Appl. Microbiol. Biotechnol. 53, 323-327. [ Links ]
Kim H.J., Kim D.Y., Nam J.S., Bae K.S. y Rhee Y.H. (2003). Characterization of an extracellular medium-chain-length poly(3-hydroxyalkanoate) depolymerase from Streptomyces sp. KJ-72. A. Van Leeuwen. J. 83, 183-189. [ Links ]
Kim H.J., Ryu K.E., Bae K.S. y Rhee Y.H. (2000). Purification and characterization of extracellular medium-chain-length polyhydroxyalkanoate depolymerise from Pseudomonas sp. RY-1. J. Biosci. Bioeng. 89, 196-198. [ Links ]
Kim S.W., Kim P., Lee H.S. y Kim J.H. (1996). High production of poly-beta-hydroxybutyrate (P3HB) from Methylobacterium organophilum under potassium limitation. Biotechnol. Lett. 18, 25-30. [ Links ]
Kim B.S.y Chang H.N. (1998). Production of poly (3-hydroxybutyrate) from starch by Azotobacter chroococcum. Biotechnol. Lett. 2, 109-112. [ Links ]
Kita K., Mashiba S., Nagita M., Ishimaru K., Okamoto K. y Yanase H. (1997). Cloning of poly(3-hydroxybutyrate) depolymerase from a marine bacterium, Alcaligenes faecalis AE122, and characterization of its gene product. Biochim. Biophys. 1352, 113-122. [ Links ]
Klingbeil B., Kroppenstedt R.M. y Jendrossek D. (1996). Taxonomic identification of Streptomyces exfoliatus K10 and characterization of its poly (3-hydroxybutyrate) depolymerase gene. FEMS Microbiol. Lett.142, 215-221. [ Links ]
Kobayashi T. y Saito T. (2003). Catalytic triad of intracellular poly(3-hydroxybutyrate) depolymerase (PhaZ1) in Ralstonia eutropha H16. J. Biosci. Bioeng. 96, 487-492. [ Links ]
Kobayashi T., Shiraki M., Abe T., Sugiyama A. y Saito T. (2003b). Purification and properties of an intracellular 3-hydroxybutyrate-oligomer hydrolase (PhaZ2) in Ralstonia eutropha H16 and its identification as a novel intracellular poly (3- hydroxybutyrate) depolymerase. J. Bacteriol. 185, 3485-3490. [ Links ]
Koller M., Bona R., Chiellini E., Fernandes E.G., Horvat P., Kutschera C., Hesse P. y Braunegg G. (2008). Polyhydroxyalkanoate production from whey by Pseudomonas hydrogenovora. Bioresource Technol. 99, 4854-63. [ Links ]
Korotkova N. y Lidstrom M.E. (2001). Connection between poly-beta-hydroxybutyrate biosynthesis and growth on C-1 and C-2 compounds in the methylotroph Methylobacterium extorquens AM1. J. Bacteriol. 183, 1038-1046. [ Links ]
Kulkarni S.O., Kanekar P.P., Nilegaonkar S.S. y Jog J.P. (2010). Production and characterization of a biodegradable poly (hydroxybutyrate-co-hydroxyvalerate) (P3HB-co-PHV) copolymer by moderately haloalkali-tolerant Halomonas campisalis MCM B-1027 isolated from Lonar Lake, India. Bioresource Technol. 101, 9765-9771. [ Links ]
Labuzek S. y Radecka I. (2001). Biosynthesis of P3HB tercopolymer by Bacillus cereus UW85. J. Appl. Microbiol. 90, 353-357. [ Links ]
Law J.H. y Slepecky R. (1961). Assay of poly-beta-hydroxybutyric acid. J. Bacteriol. 82, 33-36. [ Links ]
Lee E.Y., Kang S.H. y Choi C.Y. (1995). Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by newly isolated Agrobacterium Sp Sh-1 and gw-014 from structurally unrelated single carbon substrates. J. Ferm. Bioeng. 79, 328-334. [ Links ]
Lee S.Y. y Chang H.N. (1995). Production of poly-(hydroxyalkanoic acid). Adv. Biochem. Eng./ Biotechnol. 52, 27-58. [ Links ]
Lee S.Y. (1996). Bacterial polyhydrohyalkanoates. Biotechnol. Bioeng. 49, 1-14. [ Links ]
Lee W.H., Azizan M.N.M. y Sudesh K. (2004). Effects of culture conditions on the composition of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) synthesized by Comamonas acidovorans. Poly. Degrad. Stab. 84, 129-134. [ Links ]
Lenz R.W. y Marchessault R.H. (2005). Bacterial polyesters: biosynthesis, biodegradable plastics and biotechnology. Biomacromolecules 6, 1-8. [ Links ]
Lenz R.W. (1993). Biodegradable polymers. Adv. Polym. Sci. 107, 1-40. [ Links ]
Loo C.Y. y Sudesh K. (2007). Biosynthesis and native granule characteristics of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) in Delftia acidovorans. Int. J. Biol. Macromol. 40, 466-471. [ Links ]
López-Cuellar M.R., Alba-Flores J., Gracida-Rodríguez J. y Pérez- Guevara F. (2011). Production of polyhydroxyalkanoates (PHAs) with canola oil as carbon source. Int. J. Biol. Macromol. 48, 74-80. [ Links ]
Luengo J.M., Garcia B., Sandoval A., Naharro G. y Olivera E.R. (2003). Bioplastics from microorganisms. Curr. Opin. Microbiol. 6, 251-260. [ Links ]
Lugg H., Sammons R.L., Marquis P.M., Hewitt C.J., Yong P., Paterson-Beedle M., Redwood M.D., Stamboulis A., Kashani M., Jenkins M. y Macaskie L.E. (2008). Polyhydroxybutyrate accumulation by a Serratia sp. Biotechnol. Lett. 30, 481-491. [ Links ]
Luzier W.D. (1992). Materials derived from biomass/ biodegradable materials. Proc. Natl. Acad. Sci. USA 89, 839-842. [ Links ]
Maness P.C. y Weaver P.F. (1994). Production of poly-3-hydroxyalkanoates from co and h-2 by a novel photosynthetic bacterium. Appl. Biochem. Biotechnol. 45, 395-406. [ Links ]
Marchessault R.H., Morin F.G., Wong S. y Saracovan I. (1995). Artificial granule suspensions of long side-chain poly(3-hydroxyalkanoate). Can. J. Microbiol. 41, 138-142. [ Links ]
Matavulj M. y Molitoris H.P. (1992). Fungal degradation of polyhydroxyalkanoates and a semiquantitative assay for screening their degradation by terrestrial fungi. FEMS Microbiol. Rev. 103, 323-332. [ Links ]
McLellan D.W. y Halling P.J. (1988). Acid-tolerant poly(3-hydroxybutyrate) hydrolases from moulds. FEMS Microbiol. Lett . 52, 215-218. [ Links ]
Mergaert J., Anderson C., Wouters A. y Swings J. (1994). Microbial degradation of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) in compost. J. Environ. Polym. Degrad. 2, 177-183. [ Links ]
Mergaert J., Anderson C., Wouters A., Swings J. y Kersters K. (1992). Biodegradation of poly(hydroxyalkanoates). FEMS Microbiol. Rev. 103, 317-322. [ Links ]
Mergaert J., Webb A., Anderson C., Wouters A. y Swings J. (1993). Microbial degradation of poly(3-hydroxybutyrate) and poly(3hydroxybutyrate-co-3-hydroxyvalerate) in soils. Appl. Environ. Microbiol. 59, 3233-3238. [ Links ]
Mergaert J., Wouters A., Anderson C. y Swings J. (1995). In situ biodegradation of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) in natural waters. Can. J. Microbiol. 41, 154-159. [ Links ]
Mergaert J. y Swings J. (1996). Biodiversity of microorganisms that degrade bacterial and synthetic polymers. J. Ind. Microbiol. 17, 463-469. [ Links ]
Merrick J.M., Lundgren D.G. y Pfister R.M. (1965). Morphological changes in poly-beta-hydroxybutyrate granules associated with decreased susceptibility to enzymatic hydrolysis. J. Bacteriol. 89, 234-239. [ Links ]
Merrick J.M. y Doudoroff M. (1964). Depolymerization of poly-β-hydroxybutyrate by an intracellular enzyme system. J. Bacteriol. 88, 60-71. [ Links ]
Mineki S., Fukutome N., Oinuma N., Nagashima H. y Iida M. (1997). Synthesis and characterization of polyesters produced by Paracoccus sp. 12-a from formic acid. Macromolecules 30, 319-321. [ Links ]
Mittendorf V., Robertson E., Leech R., Kruger N., Steinbüchel A. y Poirier Y. (1998). Synthesis of medium-chain-length polyhydroxyalkanoates in Arabidopsis thaliana using intermediates of peroxisomal fatty acid b-oxidation. Appl. Biol. Sci. 99, 3397-13402. [ Links ]
Miyake M., Erata M. y Asada Y. (1996). A thermophilic Cyanobacterium, Synechococcus Sp. MA19, capable of accumulating poly-beta-hydroxybutyrate. J. Ferm. Bioeng. 82, 512-514. [ Links ]
Miyazaki S., Takahashi K., Shiraki M., Saito T., Tezuka Y. y Kasuya K. (2000). Properties of a poly(3hydroxybutyrate) depolymerase from Penicillium funiculosum. J. Polym. Environ. 8, 175-182. [ Links ]
Mohammad H., Ibrahim A. y Steinbüchel A. (2009). poly(3-hydroxybutyrate) production from glycerol by Zobellella denitrificans MW1 via high-cell-density fed-batch fermentation and simplified solvent extraction. Appl. Env. Microbiol. 75, 6222-6231. [ Links ]
Mothes C., Schnorpfeil C. y Ackermann J.U. (2007). Production of P3HB from crude glycerol. Eng. Life Sci. 7, 475-479 [ Links ]
Mukai K., Yamada K. y Doi Y. (1994). Efficient hydrolysis of polyhydroxyalkanoates by Pseudomonas stutzeri YM1414 isolated from lake water. Polym. Degrad. Stab. 43, 319-327. [ Links ]
Mukai K., Yamada K. y Doi Y. (1993a). Enzymatic degradation of poly(hydroxyalkanoates) by a marine bacterium. Polym. Degrad. Stab. 41, 85-91. [ Links ]
Mukai K., Doi Y., Sema Y. y Tomita K. (1993b). Substrate specificities in hydrolysis of polyhydroxyalkanoates by microbial esterases. Biotechnol. Lett., 15, 601-604. [ Links ]
Müller B. y Jendrossek D. (1993). Purification and properties of poly(3-hydroxyvaleric acid) depolymerase from Pseudomonas lemoignei. Appl. Microbiol. Biotechnol. 38, 487-492. [ Links ]
Murase T., Iwata T. y Doi T. (2001a). Direct observation of enzymatic degradation behavior of poly((R)-3-hydroxybutyrate) lamellar single crystals by atomic force microscopy. Macromolecules 34, 5848-5853. [ Links ]
Murase T., Suzuki Y., Doi Y. y Iwata T. (2002). Non hydrolytic fragmentation of a poly((R)-3-hydroxybutyrate) single crystal revealed by use of a mutant of polyhydroxybutyrate depolymerase. Biomacromolecules 3, 312-317. [ Links ]
Ng K.S., Ooi W.Y., Goh L.K., Shenbagarathai R. y Sudesh K. (2010). Evaluation ofjatropha oil to produce poly(3-hydroxybutyrate) by Cupriavidus necator H16. Poly. Degrad. Stab. 45, 1365-1369. [ Links ]
Nishida H. y Tokiwa Y. (1995). Confirmation of colonization of degrading bacterium strain Sc-17 on poly(3-hydroxybutyrate) cast film. J. Environ. Polym. Degr. 3, 187-197. [ Links ]
Nishida H. y Tokiwa Y. (1993). Distribution of P3HB and PCL aerobic degrading microorganisms in different environments. J. Environ. Polym. Degrad. 1, 227-233. [ Links ]
Nojima S., Mineki S. y Iida M. (1996). Purification and characterization of extracellular poly(3-hydroxybutyrate) depolymerase produced by Agrobacterium sp. K-03. J. Ferm. Bioeng. 81, 72-75. [ Links ]
Nojiri M. y Saito T. (1997). Structure and function of poly(3-hydroxybutyrate) depolymerase from Alcaligenes faeccalis T1. J. Bacteriol. 179, 6965-6970. [ Links ]
Nonato R.V., Mantelatto P.E. y Rossell C.E.V. (2001). Integrated production of biodegradable plastic, sugar and ethanol. Appl. Microbiol. Biotechnol. 57, 1-5. [ Links ]
Obruca S., Marova I., Snajdar O., Mravcova L. y Svoboda Z. (2010). Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) by Cupriavidus necator from waste rapeseed oil using propanol as a precursor of 3-hydroxyvalerate. Biotechnol. Lett. 32, 1925-1932. [ Links ]
Oda Y., Asari H., Urakami T. y Tonomura K. (1995). Microbial degradation of poly(3-hydroxybutyrate) and polycaprolactone by filamentous fungi. J. Ferment. Bioeng. 80, 265-269. [ Links ]
Oda Y., Osaka H., Urakami T. y Tonomura K. (1997). Purification and properties of poly(3-hydroxybutyrate) depolymerase from the fungus Paecilomyces lilacinus D218. Curr. Microbiol. 34, 230-232. [ Links ]
Omar S., Rayes A., Eqaab A., Voss I. y Steinbüchel A. (2001). Optimization of cell growth and poly(3-hydroxybutyrate) accumulation on date syrup by a Bacillus megaterium strain. Biotechnol. Lett. 23, 1119-1123. [ Links ]
Ostle A. y Holt J.G. (1982). Nile Blue A as a Fluorescent Stain for Poly-3-Hydroxybutyrate. Appl. Env. Microbiol. 44, 238-241. [ Links ]
Ouyang S.P., Luo R.C., Chen S.S., Liu Q., Chung A., Wu Q. y Chen G.Q. (2007). Production of polyhydroxyalkanoates with high 3-hydroxydodecanoate monomer content by fadB and fadA knockout mutant of Pseudomonas putida KT2442. Biomacromolecules. 8, 2504-2511. [ Links ]
Page W.J. (1989). Production of poly-beta-hydroxybutyrate by Azotobacter vinelandii strain UWD During growth on molasses and other complex carbon-sources. Appl. Microbiol. Biotechnol. 31, 329-333. [ Links ]
Page W.J. (1992). Production of polyhydroxyalkanoates by Azotobacter vinelandii UWD in beet molasses culture. FEMS Microbiol. Rev. 103, 149-157. [ Links ]
Page W.J., Bhanthumnavin N., Manchak J. y Ruman M. (1997). Production of poly(beta-hydroxybutyrate-beta-hydroxyvalerate) copolymer from sugars by Azotobacter salinestris. Appl. Microbiol. Biotechnol. 48 ,88-93. [ Links ]
Page W.J. y Cornish A. (1993). Growth of Azotobacter vinelandii UWD in fish peptone medium and simplified extraction of poly-beta-hydroxybutyrate. Appl. Environ. Microbiol. 59, 4236-4244. [ Links ]
Page W.J. y Knosp O. (1989). Hyperproduction of poly-beta-hydroxybutyrate during exponential-growth of Azotobacter vinelandii UWD. Appl. Env. Microbiol. 55, 1334-1339. [ Links ]
Pagga U., Schafer A., Müller R-J. y Pantke M. (2001). Determination of the aerobic biodegradability of polymeric material in aquatic batch tests. Chemosphere 42, 319-331. [ Links ]
Pal S., Manna A. y Paul A.K. (1999). Production of poly(beta-hydroxybutyric acid) and exopolysaccharide by Azotobacter beijerinckii WDN-01. World J. Microb. Biot. 15, 15-21. [ Links ]
Panda B., Jain P., Sharma L. y Mallick N. (2006). Optimization of cultural and nutritional conditions for accumulation of poly-beta-hydroxybutyrate in Synechocys-tis sp PCC 6803. Bioresour. Technol. 97, 1296-1301. [ Links ]
Pandian S.R., Deepak V., Kalishwaralal K., Rameshkumar N., Jeyaraj M. y Gurunathan S. (2010). Optimization and fed-batch production of P3HB utilizing dairy waste and sea water as nutrient sources by Bacillus megaterium SRKP-3. Bioresour. Technol. 101, 705-711. [ Links ]
Papaneophytou C.P., Pantazaki A.A. y Kyriakidis D.A. (2009). An extracellular polyhydroxybutyrate depolymerase in Thermus thermophilus HB8. Appl. Microbiol. Biotechnol. 83, 659-668. [ Links ]
Papaneophytoua C.P., Velalia E.E. y Pantazakin A. A. (2011). Purification and characterization of an extracellular medium-chain length polyhydroxyalkanoate depolymerase from Thermus thermophilus HB8. Poly. Degrad. Stab. 96, 670-678. [ Links ]
Park D.H. y Kim B.S. (2011). Production of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-4-hydroxybutyrate) by Ralstonia eutropha from soybean oil. N. Biotechnol. 28, 719-724. [ Links ]
Patwardhana P. y Srivastava A.K. (2008). Fed-batch cultivation of Wautersia eutropha. Bioresour. Technol. 99, 1787-1792. [ Links ]
Pohlmann A., Fricke W.F., Reinecke F., Kusian B., Liesegang H., Cramm H., Eitinger T., Ewering C., Potter M., Schwart S. A., Voss I., Gootschalk G., Steinbüchel A. y Bowien F. B. (2006). Genome sequence of the bioplastic-producing "Knallglas" bacterium Ralstonia eutropha H16. Nat. Biotechnol. 24, 1227-1229. [ Links ]
Porwal S., Kumara T., Lal S., Rani A., Kumar S., Cheema S., Purohitd H., Sharma R., Patel S. y Kalia V.C. (2008). Hydrogen and polyhydroxybutyrate producing abilities of microbes from diverse habitats by dark fermentative process. Bioresour. Technol. 13, 5444-5451 . [ Links ]
Pozo C., Martinez-Toledo M.V., Rodelas B. y Gonzalez-Lopez J. (2002). Effects of culture conditions on the production of polyhydroxyalkanoates by Azotobacter chroococcum H23 in media containing a high concentration of alpechin (wastewater from olive oil mills) as primary carbon source. J. Biotechnol. 97, 125-131. [ Links ]
Püchner P., Müller W.R. y Bartke D. (1995). Assessing the biodegradation potential of polymers in screening and long term test systems. J. Env. Poly. Deg. 3, 133-143. [ Links ]
Qin L.F., Gao X., Liu Q., Wu Q. y Chen G.C. (2007). Biosynthesis of polyhydroxyalkanoate copolyesters by Aeromonas hydrophila mutant expressing a low-substrate-specificity PHA synthase PhaC2Ps. Biochem. Eng. J. 37, 144-150. [ Links ]
Qingsheng Q. y Rehm B. (2001). Polyhydroxybutyrate biosynthesis in Caulobacter crescentus, molecular characterization of the polyhydroxybutyrate synthase. Microbiology. 147, 3353-3358. [ Links ]
Quillaguaman J., Hashim S., Bento F., Mattiasson B. y Hatti-Kaul R. (2005). Poly(beta-hydroxybutyrate) production by a moderate halophile, Halomonas boliviensis LC1 using starch hydrolysate as substrate. J. Appl. Microbiol. 99, 151-157. [ Links ]
Quillaguaman J., Munoz M., Mattiasson B. y Hatti-Kaul R.(2007). Optimizing conditions for poly(beta-hydroxybutyrate) production by Halomonas Boliviensis LC1 in batch culture with sucrose as carbon source. Appl. Microbiol. Biotechnol. 74, 981-986. [ Links ]
Ramadas N., Kumar S., Soccol C. y Pandey A. (2009). Polyhydroxybutyrate production using agro-industrial residue as substrate by Bacillus sphaericus NCIM 5149. Braz. Arch. Biol. Technol. 52, 17-23. [ Links ]
Ramsay B.A., Lomaliza K., Chavarie C., Dube B., Bataille P. y Ramsay J.A. (1990). Production of poly-(beta-hydroxybutyric-co-beta-hydroxyvaleric) acids. Appl. Env. Microbiol. 56, 2093-2098. [ Links ]
Ramsay B.A., Saracovan I., Ramsay J.A y Marchessault R.H. (1994). A method for the isolation of microorganism producing extracellular long-side-chain poly(b-hydroxyalkanoate) depolymerase). J. Environ. Polym. Degrad. 2, 1-7. [ Links ]
Ran N., Zhao L., Chen Z. y Tao J. (2008). Recent applications of biocatalysis in developing green chemistry for chemical synthesis at the industrial scale. Green Chem. 10, 361-372. [ Links ]
Reddy C. S. K., Ghai R., Rashmi y Kalia V. C. (2003). Polyhydroxyalkanoates: an overview. Bioresour. Technol. 87, 137-146. [ Links ]
Rehm B.H.A. (2003). Polyester Synthases: Natural Catalysts for Plastics. Biochem. J. 376, 15-33. [ Links ]
Renner G., Pongratz K. y Braunegg G. (1996). Production of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) by Comamonas testosteronii A3. Food Technol. Biotech- nol. 34, 91-95. [ Links ]
Renstad R., Karlsson S. y Albertsson A-C. (1999). The influence of processing induced differences in molecular structure on the biological and non-biological degradation of poly(3-hydroxybutyrate-co-3-hydroxyvalerate), P(3-HB-co-3-HV). Polym. Degrad. Stab. 63, 201-211. [ Links ]
Rhee Y.A., Kim Y.H. y Shin K.S. (2006). Characterization of an extracellular poly(3-hydroxyoctanoate) depoly-merase from the marine isolate Pseudomonas luteola M13-4. Enzyme Microb. Technol. 38, 529-535. [ Links ]
Romen F., Reinhardt S. y Jendrossek D. (2004). Thermo-tolerant poly(3hydroxybutyrate)-degrading bacteria from hot compost and characterization of the P3HB depolymerase of Schlegellella sp. KB1a. Arch. Microbiol. 182, 157-164. [ Links ]
Ryu H.W., Cho K.S., Lee E.G. y Chang Y.K. (2000). Recovery of Poly(3-Hydroxybutyrate) from Coagulated Ralstonia eutropha using a chemical digestion method. Biotechnol. Prog.16, 676-679. [ Links ]
Saegusa H., Shiraki M., Kanai C. y Saito T. (2001). Cloning of an intracellular poly(D()-3-hydroxybutyrate) depolymerase gene from Ralstonia eutropha H16 and characterization of the gene product. J. Bacteriol. 183, 94-100. [ Links ]
Saeki T., Tsukegi T., Tsuji H., Daimon H. yFujie K. (2005). Hydrolytic degradation of poly ((R)-3-hydroxybutyric acid) in the melt. Polymer. 46, 2157-2162. [ Links ]
Saito Y. y Doi Y.(1994). Microbial synthesis and properties of poly(3-hydroxybutyrate-co-4-hydroxybutyrate) in Comamonas acidovorans. Int. J. Biol. Macromol. 16, 99-104. [ Links ]
Saito T., Saegusa H., Miyata Y. y Fukui T. (1992). Intracellular degradation of poly(3-hydroxybutyrate) granules of Zoogloea ramigera I-16-M, FEMS Microbiol. Rev. 103, 333-338. [ Links ]
Salehizadeh H. y Van Loosdrecht M.C.M. (2004). Production of polyhydroxyalkanoates by mixed culture: recent trends and biotechnological importance. Biotechnol. Adv. 22, 261-279. [ Links ]
Samrot A.V., Avinesh R., Sukeetha S. y Senthilkumar P. (2011). Accumulation of Poly((R)-3-hydroxyalkanoates) in Enterobacter cloacae SU-1 during growth with two different carbon sources in batch culture. Appl. Biochem. Biotechnol. 163, 195-203. [ Links ]
Sanchez J.G., Tsuchii A. y Tokiwa Y. (2000). Degradation of polycaprolactone at 50 °C by a thermotolerant Aspergillus sp. Biotechnol. Lett. 22, 849-853. [ Links ]
Sang B-I., Hori K., Tanji Y. y Unno H. (2002). Fungal contribution to in situ biodegradation of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) film in soil. Appl. Microbiol. Biotechnol. 58, 241-247. [ Links ]
Sangkharak K. y Prasertsan P. (2007). Optimization of polyhydroxybutyrate production from a wild type and two mutant strains of Rhodobacter sphaeroides using statistical method. J. Biotechnol. 132, 331-340. [ Links ]
Scherer T., Fuller M.R.C., Goodwin S. y Lenz R.W. (2000a). Enzymatic hydrolysis of oligomeric models of poly-3-hydroxybutyrate. Biomacromolecules 1, 577-583. [ Links ]
Scherer T., Rothermich M.M., Quinteros R., Poch M.T., Lenz R.W. y S. Goodwin. (2000b). Broad-based screening of polymer biodegradability in polymers from renewable resources. En: Symposium Series, Biopolyesters and biocatalysis (C. Scholz y R.A. Gross, Eds.). 764 ACS, 254-280, Oxford University Press, Washington, DC, EUA. [ Links ]
Scherer T.M., Fuller R.C., Lenz R.W. y Goodwin S. (1999). Hydrolysis activity of an extracellular depolymerase from Aspergillus fumigatus with bacterial and synthetic polyesters. Polym. Degrad. Stab. 64, 267-275. [ Links ]
Schirmer A., Jendrossek D. y Schlegel H. (1993). Degradation of poly(3-hydroxyoctanoic acid) (P(3HO)) by bacteria: purification and properties of a P(3HO) depolymerase from Pseudomonas fluorescens GK13. Appl. Environ. Microbiol. 59, 1220-1227. [ Links ]
Schirmer A., Matz C. y Jendrossek D. (1995). Substrate specificities of poly(hydroxyalkanoate)-degrading bacteria and active site studies on the extracellular poly(3-hydroxyoctanoic acid) depolymerase of Pseudomonas fluorescens GK13. Can. J. Microbiol.41 (Suppl. 1), 170-179. [ Links ]
Schober U., Thiel C. y Jendrossek D. (2000). Poly (3-Hydroxyvalerate) depolymerase of Pseudomonas lemoignei. Appl. Env. Microbiol. 66, 1385-1392. [ Links ]
Schulze B. y Wubbolts M.G. (1999). Biocatalysis for industrial production of fine chemicals. Curr. Opin. Biotechnol. 10, 609-615. [ Links ]
Scott G. (1990). Photo-biodegradable plastics: their role in the protection of the environment. Poly. Degrad. Stab. 29, 135-154. [ Links ]
Senior P.J. y Dawes E.A. (1971). Poly-beta-hydroxy-butyrate biosynthesis and regulation of glucose metabolism in Azotobacter beijerinckii. Biochem. J. 125, 55-66. [ Links ]
Shah N.N., Hanna M.L. y Taylor R.T. (1996). Batch Cultivation of Methylosinus trichosporium OB3b .5. characterization of poly-beta-hydroxybutyrate production under methane-dependent growth conditions. Biotechnol. Bioeng. 49, 161-171. [ Links ]
Shang L., Jiang M., Yun Z., Yan H.Q. y Chang H.N. (2008). Mass production of medium-chain-length poly(3-hydroxyalkanoates) from hydrolyzed corn oil by fed-batch culture of Pseudomonasputida. World J. Microbiol. Biot. 24, 2783-2787. [ Links ]
Shi H.P., Lee C.M. y Ma W.H. (2007). Influence of electron acceptor, carbon, nitrogen, and phosphorus on polyhydroxyalkanoate (PHA) production by Brachymonas sp P12. World J. Microb. Biot. 23, 625-632. [ Links ]
Shiraki M., Shimada T., Tatsumchi M. y Saito T. (1995). Purification and characterization of extracellular poly(3-hydroxybutyrate) depolymerases. J. Environ. Polym. Degrad. 3, 13-21. [ Links ]
Shirakura Y., Fukui T., Saito T., Okamoto Y., Narikawa T., Koide K., Tomita K., Takemasa T. y Masamune S. (1986). Degradation of poly(3-hydroxybutyrate) by poly(3-hydroxybutyrate) depolymerase from Alcaligenes faecalis T1. Biochim. Biophys. 880, 46-53. [ Links ]
Silva L.F., Taciro M.K., Ramos M.E.M., Carter J.M., Pradella J.G. y Gomez J.G.(2004) Poly-3-hydroxy-butyrate (P3HB) production by bacteria from xylose, glucose and sugarcane bagasse hydrolysate. J. Ind. Microbiol. Biotechnol. 31, 245-254. [ Links ]
Simon-Colin C., Alain K., Colin S., Cozien J., Costa B., Guezennec J. y Raguénés G. (2008) . A novel mcl PHA-producing bacterium, Pseudomonas guezennei sp. nov., isolated from a 'kopara' mat located in Rangiroa, an atoll of French Polynesia. J. Appl. Microbiol. 104, 581-586. [ Links ]
Spyros A. y Kimmich R. (1997). H NMR Imaging study of enzymatic degradation in poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3hydroxyvalerate). Evidence for Preferential degradation of the amorphous phase by P3HB depolymerase B from Pseudomonas lemoignei. Macromolecules 30, 8218-8225. [ Links ]
Steinbüchel A. (2001). Perspectives for biotechnological production and utilization of biopolymers: Metabolic engineering of polyhydroxyalkanoate biosynthesis pathways as a successful example. Macromol. Biosci. 1, 1-24. [ Links ]
Steinbüchel A., Debzi E.M., Marchessault R.H. y Timm A. (1993). Synthesis and production of poly(3-hydroxyvaleric acid) homopolyester by Chromobacterium violaceum. Appl. Microbiol. Biotechnol. 39, 443-449. [ Links ]
Steinbüchel A. y Valentin H. (1995). Diversity of bacterial polyhydroxyalkanoic acids FEMS Microbiol. Lett. 128, 219-228. [ Links ]
Stockdale H., Ribbons D.W. y Dawes E.A. (1968). Occurrence of poly-beta-hydroxybutyrate in the Azoto-bacteriaceae. J. Bacteriol. 95, 1798-1803 [ Links ]
Stuart E.S., Foster L.J.R., Lenz R.W. y Fuller R.C. (1996). Intracellular depolymerase functionality and location in Pseudomonas oleovorans inclusions containing polyhydroxyoctanoate. Int. J. Biol. Macromol. 19, 171-176. [ Links ]
Sudesh K., Abe H. y Doi Y. (2000). Synthesis, structure and properties of polyhydroxyalkanoates biological polyesters. Prog. Polym. Sci. 25, 1503-1555. [ Links ]
Sun W.Q., Cao J.G., Teng K. y Meighen E.A. (1994). Biosynthesis of poly-3-hydroxybutyrate in the luminescent bacterium, Vibrio harveyi, and regulation by the lux autoinducer, n-(3-hydroxybutanoyl)homoserine lactone. J. Biol. Chem. 269, 20785-20790. [ Links ]
Suzuki T., Yamane T. y Shimizu S. (1986). Kinetics and effect of nitrogen-source feeding on production of poly-beta-hydroxybutyric acid by fed-batch culture. Appl. Microbiol. Biotechnol. 24, 366-369. [ Links ]
Takaku H., Kimoto A., Kodaira S., Nashimoto M. y Takagi M. (2006). Isolation of a Grampositive poly(3-hydroxybutyrate) (P3HB)-degrading bacterium from compost, and cloning and characterization of a gene encoding P3HB depolymerase of Bacillus megaterium N-18-25-9. FEMS Microbiol. Lett. 264, 152-159. [ Links ]
Takeda M., Kamagata Y., Ghirose W.C., Hanada S. y Koizumi J. (2002). Caldimonas manganoxidans gen. nov., sp nov., a poly(3hydroxybutyrate)-degrading, manganese-oxidizing thermophile. Int. J. Syst. Evol. Microbiol. 52, 895-900. [ Links ]
Takeda M., Koizumi J., Yabe K. y Adachi K. (1998). Thermostable poly(3-hydroxybutyrate) depolymerase of a thermophilic strain of Leptothrix sp. isolated from hot spring. J. Ferment. Bioeng. 85, 375-380. [ Links ]
Tan D., Xue Y., Aibaidul G. y Chen G.Q. (2011). Unsterile and continuous production of polyhydroxybutyrate by Halomonas TD01. Bioresour. Technol. 102, 8130-8136. [ Links ]
Tanaka K., Ishizaki A., Kanamaru T. y Kawano T. (1995). Production of poly(d-3-hydroxybutyrate) from CO2: H2: and O2 by high cell-density autotrophic cultivation of Alcaligenes eutrophus. Biotechnol. Bioeng. 45, 268-275 [ Links ]
Tanaka K., Katamune K. y Ishizaki A. (1993). Fermentative production of poly-beta-hydroxy-butyric acid from xylose by a 2-stage culture method employing Lactococcus lactis Io-1 and Alcaligenes eutrophus. Biotechnol. Lett. 15, 1217-1222. [ Links ]
Tansengco M.L. y Tokiwa Y. (1998b). Thermophilic microbial degradation of polyethylene succinate. World J. Microbiol. Biot. 14, 133-138. [ Links ]
Thakor N.S., Patel M.A., Trivedi U.B. y Patel K.C. (2003). Production of poly(beta-hydroxybutyrate) by Comamonas testosteroni during growth on naphthalene. World. J. Microb. Biot.19, 185-189. [ Links ]
Tian J., Sinskey A. y Stubbe J. (2005). Kinetic Studies of Polyhydroxybutyrate Granule Formation in Wautersia eutropha H16 by Transmission Electron Microscopy J. Bacteriol. 187, 3814-3824. [ Links ]
Tokiwa Y., Ando T. y Suzuki T. (1976). Degradation of polycaprolactone by a fungus. J. Ferment. Tehcnol. 54 ,603-608 [ Links ]
Tokiwa Y. y Calabia B.P. (2004a). Degradation of microbial polyesters. Biotechnol. Lett. 26, 1181-1189. [ Links ]
Tokiwa Y. y Jarerat A. (2004b). Biodegradation of poly( L-lactide). Biotechnol. Lett. 26, 771-777. [ Links ]
Uchino K., Saito T. y Jendrossek D. (2008). Poly(3-hydroxybutyrate) (P3HB) depolymerase PhaZa1 is involved in mobilization of accumumated P3HB in Ralstonia eutropha H 16. Appl. Environ. Microbiol. 74, 1058-1063. [ Links ]
Ueda S., Matsumoto S., Takagi A. y Yamane T. (1992). Synthesis of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) from methanol and n-amyl alcohol by the methylotrophic bacteria Paracoccus denitrificans and Methylobacterium extorquens. Appl. Env. Microbiol. 58, 3574-3579. [ Links ]
Uefuji M., Kasuya K. y Doi Y. (1997). Enzymatic degradation of poly((R)-3-hydroxybutyrate): secretion and properties of P3HB depolymerase from Pseudomonas stutzeri. Polym. Degrad. Stab. 58, 275-281. [ Links ]
Valappil S.P., Boccaccini A.R., Bucke C. y Roy I. (2007). Polyhydroxyalkanoates in Gram-positive bacteria: insights from the genera Bacillus and Streptomyces. A. Van Leeuwen. J. 91, 1-17. [ Links ]
Valentin H.E., Schonebaum A. y Steinbüchel A. (1992). Identification of 4-hydroxyvaleric acid as a constituent of biosynthetic polyhydroxyalkanoic acids from bacteria. Appl. Microbiol. Biotechnol. 36, 507-514. [ Links ]
Valentin H.E., Schonebaum A. y Steinbüchel A. (1996). Identification of 5-hydroxyhexanoic acid, 4-hydroxy-heptanoic acid and 4-hydroxyoctanoic acid as new constituents of bacterial polyhydroxyalkanoic acids. Appl. Microbiol. Biotechnol. 46, 261-267. [ Links ]
Vidal-Mas J., Resina-Pelfort O., Haba E., Comas J., Manresa A. y Vives-Rego J. (2001). Rapid flow cytometry - Nile red assessment of PHA cellular content and heterogeneity in cultures of Pseudomonas aeruginosa 47T2 (NCIB 40044) grown in waste frying oil. A. Van Leeuwen. J. 80, 57-63 [ Links ]
Volova T.G., Boyandin A.N., Vasiliev A.D., Karpov VA., Prudnikova S.V., Boyarskikh U.A., Filipenko M.L., Rudnev VP., Büi Bá X., VuViet D. y Gitelson I.I. (2010). Biodegradation of polyhydroxyalkanoates (PHAs) in tropical coastal waters and identification of PHA-de-grading bacteria. Polym. Degrad. Stab. 95, 2350-2359. [ Links ]
Wang F.L. y Lee S.Y. (1997). Production of poly(3-hydroxybutyrate) by fed-batch culture of filamentation-suppressed recombinant Escherichia coli. Appl. Env. Microbiol. 63, 4765-4769. [ Links ]
Wang J., Yue Z., Sheng G. y Yu H. (2010). Kinetic analysis on the production of polyhydroxyalkanoates from volatile fatty acids by Cupriavidus necator with a consideration of substrate inhibition, cell growth, maintenance, and product formation. Biochem. Eng. J. 49, 422-428 [ Links ]
Williams S.F., Martin D.P., Horowitz D.M. y Peoples O.P. (1999). PHA applications: addressing the price performance issue i. tissue engineering. Int. J. Biol. Macromol. 25, 111-121 [ Links ]
Williams S. F. y Peoples O. P. (1996). Biodegradable plastics from plants. Chemtech. 26, 38-44. [ Links ]
Xie W.P. y Chen G.Q. (2007). Production and characterization of terpolyesterpoly (3-hydroxybutyrate-co-4-hydroxybutyrate-co-3- hydroxyhexanoate) by recombinant Aeromonas hydrophila 4AK4 harboring genes phaPCJ (J). Biochem. Eng. J. 38, 384-389. [ Links ]
Yamane T., Chen X.F. y Ueda S. (1996). Growth-associated production of poly(3-hydroxyvalerate) from n-pentanol by a methylotrophic bacterium, Paracoccus denitrificans. Appl. Env. Microbiol. 62, 380-384 [ Links ]
Yamashita K., Yamada M., Numata K. y Taguchi S. (2006). Nonspecific hydrophobic interactions of a repressor protein, PhaR, with poly((R)-3-hydroxybutyrate) film studied with a quartz crystal microbalance. Macromolecules 7, 2449-2454. [ Links ]
Yang J.S. y Huang J.X. (2006). Mathematical modeling of Zoogloea sp. batch fermentation for PHAs synthesis. J. Chem. Ind. Eng. 57, 1655-1658. [ Links ]
Yezza A., Halasz A., Levadoux W. y Hawari J. (2007). Production of poly-beta-hydroxybutyrate (P3HB) by Alcaligenes latus from maple sap. Appl. Microbiol. Biot. 77, 269-274. [ Links ]
Yoon S.C. y Choi M.H. (1999). Local sequence dependence of polyhydroxyalkanoic acid degradation in Hydrogenophaga pseudoflava. J. Biol. Chem. 274, 37800-37808 [ Links ]
York G.M., Lupberger J., Tian J., Lawrence A.G., Stubbe J. y Sinskey A.J. (2003). Ralstonia eutropha H16 encodes two and possibly three intracellular poly(D-(-)-3-hydroxybutyrate) depolymerase genes. J. Bacteriol. 185, 3788-3794. [ Links ]
Yu J. y Si Y.T. (2001). A dynamic study and modeling of the formation of polyhydroxyalkanoates combined with treatment of high strength wastewater. Environ. Sci. Technol. 35, 3584-3588. [ Links ]
Yu J. y Stahl H. (2008). Microbial utilization and biopolyester synthesis of bagasse hydrolysates. Bioresour. Technol. 99, 8042-8048. [ Links ]
Zagar E, Krzan A. (2004). SEC-MALS Characterization of Microbial Polyhydroxyalkanoates. Biomacromolecules. 5, 628-636. [ Links ]
Zakaria M., Suraini A., Ariffin H. y Yee P. (2010). Comamonas sp. EB172 isolated from digester treating palm oil mill effluent as potential polyhydroxyalkanoate (PHA) producer. African J. Biotechnol.7, 4118-4121. [ Links ]
Zhao W. y Chen G.Q. (2007). Production and characterization of terpolyester poly(3-hydroxybutyrate-co-3-hydroxyvalerate-co-3-hydroxyhexanoate) by recombinant Aeromonas hydrophila 4AK4 harboring genes phaAB. Process Biochem. 42, 1342-1347. [ Links ]
Zheng Z., Li M., Xue X., Tian H., Li Z. y Chen G. (2006). Mutation on N-terminus of polyhydroxybutyrate synthase of Ralsonia eutropha enhanced P3HB accumulation. Appl. Microbiol. Biotechnol. 72, 896-905. [ Links ]

