










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista internacional de contaminación ambiental
versión impresa ISSN 0188-4999
Rev. Int. Contam. Ambient vol.26 no.1 Ciudad de México feb. 2010
Revisiones
Propiedades , aplicaciones y producción de biotensoactivos: una revisión
Biosurfactant properties, applications and production: a review
Donaji JIMÉNEZ ISLAS, Sergio A. MEDINA MORENO y Jorge Noel GRACIDA RODRÍGUEZ*
Departamento de Biotecnología, Universidad Politécnica de Pachuca. C. P. 43830, Ex–Hacienda de Santa Bárbara, Carretera Pachuca–Cd. Sahagún Km. 20, Zempoala, Hidalgo, México. *Correo electrónico: gracidaj@netscape.net
Recibido diciembre 2008
Aceptado marzo 2009
RESUMEN
Los tensoactivos son moléculas anfifílicas con una porción hidrofílica y otra hidrofóbica; se localizan preferentemente en la interfase de fluidos con diferentes grados de polaridad como son la de aceite–agua o aire–agua. Las propiedades de un tensoactivo son caracterizadas principalmente por cuatro parámetros: concentración micelar crítica (CMC), número de agregación (na), balance hidrofílico–lipofilico (HLB) y punto de nube (PN). Estos parámetros indican el potencial del tensoactivo como agente emulsificante y de superficie, determinando su aplicación en diversas áreas, principalmente a nivel industrial y ambiental. Los tensoactivos son sintetizados vía química o biológica, denominando a los últimos biotensoactivos. Aunque una amplia gama de microorganismos son capaces de sintetizar biotensoactivos, las bacterias son las principales productoras. Los biotensoactivos integran una serie de compuestos cuya estructura puede ser simple –como los ramnolípidos, formados por mono y disacáridos unidos por enlaces glicosídicos a ácidos grasos hidroxilados–, o compleja, como el emulsán, biotensoactivo polimérico complejo que contiene unidades monoméricas de heteropolisacáridos enlazados a lípidos. Los biotensoactivos muestran ventajas sobre los de síntesis química debido a su compatibilidad con el ambiente, baja toxicidad y biodegradabilidad. Sin embargo, su principal desventaja son los elevados costos asociados a su producción. En la presente revisión se describen las propiedades químicas y microbiológicas de tensoactivos y biotensoactivos, así como las diferentes aplicaciones de los últimos, principalmente en el campo de la biorremediación. Además, se analiza la influencia de las principales variables de proceso sobre la producción de biotenso–activos, en particular de los ramnolípidos provenientes de Pseudomonas aeruginosa.
Palabras clave: biotensoactivos, Pseudomonas aeruginosa, ramnolípidos, tensión superficial.
ABSTRACT
Surfactants are amphiphilic molecules containing both hydrophilic and hydrophobic parts; these substances are located at the fluid–fluid interface with different degrees of polarity from oil–water or air–water interfaces. The properties of surfactants are characterized principally by four parameters: critical micelle concentration (CMC), aggregation number (na), hydrophilic–lipophilic balance (HLB) and cloud point (CP). These parameters are indicative of the potential of the surfactant as an emulsifier and surface–active agent and determine their application, mainly in industry and the environment. Surfactants are synthesized chemically or biologically, in which case they are called biosurfactants. Although an extensive number of microorganisms are capable of synthesizing biosurfactants, they are mainly produced by bacteria. Biosurfactants make up a series of compounds whose structure may be relatively simple such as rhamnolipids, whose structures have mono– and disaccharides joined by glycosidic bonds to hydroxylated carboxylic acids, or more complex such as emulsán, a polymeric biosurfactant with monomeric units of heteropolysaccharides bound to lipids. Biosurfactants have advantages over chemical surfactants due to their compatibility with the environment, low toxicity and biodegradability. However, the principal disadvantage is their high production cost in some cases. In this paper the chemical and microbiological properties of surfactants and biosurfactants are described as well as biosurfactant applications, principally in the bioremediation field. The effect of the main process variables on the production of biosurfactants is also discussed, particularly of rhamnolipids from Pseudomonas aeruginosa.
Key words: biosurfactants, Pseudomonas aeruginosa, rhamnolipids, surface tension.
INTRODUCCIÓN
Los tensoactivos son moléculas anfifílicas que reducen la tensión en la interfase (Gautam y Tyagi 2006). Debido a esta propiedad, las moléculas de los tensoactivos se ubican en la interfase de fluidos como son los sistemas aceite–agua y aire–agua (Stoyanov et al. 2003). Además, la presencia de tensoactivos en un medio acuoso incrementa la solubilidad en agua y disponibilidad de compuestos orgánicos, siendo ésta una de sus principales propiedades para su aplicación en el área de la biotecnología ambiental para la remoción y biodegradación de contaminantes (Liu et al. 1991).
La industria de extracción y transformación del petróleo emplea tensoactivos en la limpieza de los tanques de almacenamiento para incrementar la solubilidad de hidrocarburos no miscibles en fases acuosas, tales como pentano, hexano, ciclohexano, octano, dodecano, hexadecano, tolueno y naftaleno (Perfumo et al. 2006).
Los tensoactivos son producidos vía química o biológica. Los de síntesis química están siendo remplazados por los de origen biológico (biotensoactivos) debido a que estos últimos poseen baja toxicidad (Cuadro I), son biodegradables y tienen una mejor compatibilidad con el ambiente (Nitschke et al. 2005b, Gautam y Tiagy 2006, Mukherjee et al. 2006). Los biotensoactivos son moléculas complejas producidas y secretadas por diferentes tipos de microorganismos, están integrados por estructuras diversas como péptidos, glicolípidos, glicopéptidos, ácidos grasos y fosfolípidos (Gautam y Tyagi 2006).

El interés en los biotensoactivos se ha incrementado considerablemente en años recientes por sus aplicaciones en la industria y el ambiente. Por ello, el registro en el número de patentes internacionales para la aplicación de biotensoactivos en los sectores de alimentos, limpieza doméstica, cosméticos, recuperación de crudo, remediación de sitios contaminados y aplicaciones en agricultura, crece de manera significativa año con año (Hung y Shreve 2001). No obstante, uno de los factores que limita la comercialización de biotensoactivos en algunos casos, son los elevados costos asociados a su producción en gran escala. Por esta razón, las investigaciones recientes se han enfocado tanto a la optimización de los procesos de producción como a la búsqueda de sustratos económicos, con la finalidad de disminuir los costos. En la presente revisión se plantea un panorama general de las propiedades y aplicación de los biotensoactivos, principalmente los de origen bacteriano. También se discute el efecto de algunas de las variables implicadas en la producción de biotensoactivos por diferentes microorganismos, en particular de los ramnolípidos a partir de Pseudo–monas aeruginosa.
Propiedades y clasificación de tensoactivos y biotensoactivos
La actividad de los tensoactivos comúnmente se relaciona con cambios de la tensión superficial de los líquidos. Los gradientes de tensión superficial, debido a la presencia de moléculas de un tensoactivo en las interfases aire–agua o aceite–agua, son determinados mediante tensómetros (Makkar y Cameotra 1998).
La concentración de tensoactivos en la que se alcanza la mínima tensión superficial es denominada concentración micelar crítica (CMC); es también la concentración de tensoactivos por debajo de la cual las micelas se forman espontáneamente. La CMC es un parámetro característico para cada tensoactivo. Para valores inferiores a esta concentración, todo el tensoactivo presente en solución se encuentra en forma de monómeros ubicados en la interfase agua–aire, mientras que a valores más altos, algunas moléculas de tensoactivo se encuentran en forma micelar (Fig. 1). En el cuadro II se muestran los valores de la CMC para diferentes tipos de tensoactivos.
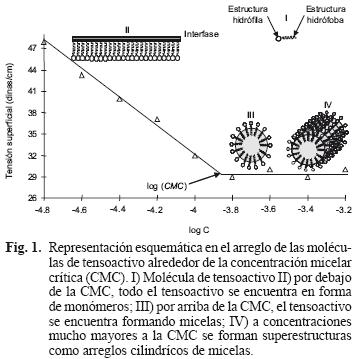

Además de la CMC, tres propiedades más caracterizan a los tensoactivos: el número de agregación (na), el balance hidrofílico–lipofílico (HLB) y el punto de nube (PN).
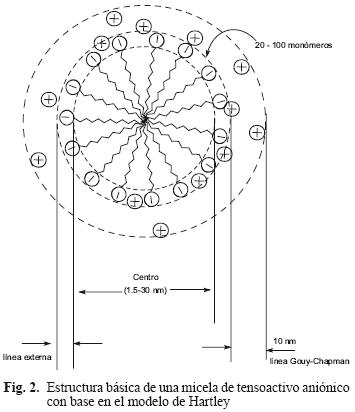
El na indica el promedio de moléculas de tenso–activo que se encuentran formando las micelas. Los agregados micelares están conformados generalmente de 20–100 monómeros, con un radio aproximado de 1.5–30 Å. El modelo físico más aceptado para la micela es el desarrollado por Hartley en 1936 y consiste en una configuración esférica con tres fases (Fig. 2). Adair et al. (1983), mediante mediciones de viscosidad a 20 °C, determinaron el número de agregación para diversos tensoactivos iónicos de cadena corta, obteniendo los siguientes valores de na: heptil sulfato de sodio (23), octal sulfato de sodio (29), dodecil sulfato de sodio (32), octanoato de sodio (16), cloruro de hexilamonio (20) y cloruro de octalamonio (25). En el área de biorremediación de suelos contaminados con hidrocarburos intemperizados de alto peso molecular, el modelo de Hartley y el na han sido utilizados para estimar el radio intermicelar de diversos tensoactivos, para así determinar la capacidad de éstos para solubilizar compuestos orgánicos hidrofóbicos (COH) de acuerdo con peso molecular (Chu 2003).
Por otra parte, el HBL es un valor que representa la capacidad de un tensoactivo para formar emulsiones estables en sistemas agua–aceite o aceite–agua, al compararse con valores estándar establecidos de HBL para los tensoactivos ácido oleico (HBL = 1) y oleato de sodio (HBL = 20). De esta manera, valores de HBL cercanos a 1 indican que el tensoactivo forma emulsiones estables en sistemas agua–aceite, mientras que valores de HBL cercanos a 20 señalan su capacidad para formar emulsiones estables en sistemas aceite–agua. En el cuadro III se da una clasificación general en la aplicación de los tensoactivos de acuerdo a los intervalos del HBL. Por ejemplo, el oleil–soforolípido, que posee un valor de HBL entre 7 y 8, ha mostrado ser un excelente humectante de piel (Brown 1991).

La temperatura también es un factor clave que incide directamente en la solubilidad de los tensoactivos en soluciones acuosas, y cuyo efecto ha sido observado principalmente en soluciones micelares de tensoactivos no iónicos (Timar–Balazsy e Eastop 1998). Este efecto se ha denominado punto de nube (PN) y se define como la temperatura a la cual el tensoactivo (generalmente no iónico) pierde su solubilidad en una solución acuosa, la cual a nivel macroscópico presenta una apariencia turbia, es decir, en forma de nube (Kim y Shah 2002). Al incrementarse la temperatura, la solubilidad de los tensoactivos decrece gradualmente y las moléculas no solubles rompen el equilibrio existente entre las del tensoactivo y las del agua, formando superagregados de tensoactivo, lo cual provoca un cambio de fase y genera la turbidez (Timar–Balazsy e Eastop 1998, Kim y Shah 2003, Kim et al. 2006). La determinación del PN se realiza utilizando una solución acuosa de tensoactivo a 1 % y es evaluada en un intervalo de 0 a 100 °C, midiendo la intensidad de luz transmitida a través de dicha solución. En el cuadro IV se muestran los PN de diferentes tipos de tensoactivos no iónicos (Van Os et al. 1993).
Dependiendo de la estructura química del anión o catión de la sal, éstas modifican el potencial químico de la solución, provocando cambios en el PN (Kissa 2001). Ejemplos de esto son el incremento en el PN de 38 a 62 °C al adicionarse NaCl a una solución de amitriptilina; y por el contrario, la disminución en el PN de 38 a 34 °C en la adición de urea a la misma solución de tensoactivo (Kim et al. 2006). Una aplicación práctica de los tensoactivos con relación a su PN ha sido para sistemas en los que es necesario una cierta generación de espuma. Así, el tensoactivo no iónico tritón C–32 con PN = 23 °C, es ideal para aplicaciones que involucren la formación de espuma a bajas temperaturas, mientras que a temperaturas mayores se recomienda el tritón–X100 cuyo PN = 66 °C (Kim et al. 2006).
Estos cuatro parámetros –CMC, na, HLB y PN– caracterizan a los tensoactivos que se utilizan en la industria y el ambiente por sus capacidades para solubilizar compuestos poco solubles, formar emulsiones entre fases inmiscibles, humectar superficies hidrófobas y determinar la temperatura a la cual se da un cambio en las propiedades del tensoactivo. De esta manera, las características y capacidades para cada grupo de tensoactivos están directamente relacionadas con su estructura química, la cual le provee ventajas en un nicho industrial o ecológico particular (Ron y Rosenberg 2001).
Los tensoactivos químicos, según su estructura hidrofílica, pueden subclasificarse en iónicos y no iónicos (Fig. 3). Los tensoactivos iónicos se caracterizan por presentar en su estructura hidrofílica grupos funcionales con cargas eléctricas negativas (aniónicos) o positivas (catiónicos), mientras que los no iónicos no presentan carga.
Dentro de los tensoactivos aniónicos, dos de los más importantes por su aplicación a nivel industrial son el lauril sulfato de sodio y el dodecil bencen sulfonato de sodio; entre los catiónicos, los más importantes son los derivados de las sales de alquil trimetil amonio. Los alquil fenol etoxilatos son los tensoactivos no iónicos más ampliamente usados, destacando las series del tritón y el tergitol (Fig. 4).
Además, a partir de la década de los 90 se han llevado a cabo investigaciones para la síntesis de una nueva clase de tensoactivos químicos denominados bitensoactivos, también llamados géminis; se caracterizan por poseer más de una parte hidrofóbica y más de un grupo hidrofílico, cualidad que al parecer les permite tener una mayor actividad superficial con respecto a los tensoactivos convencionales (Shukla y Tyagi 2006a, b).
En lo concerniente a los biotensoactivos, la parte hidrofílica consiste generalmente de alguna de las estructuras siguientes: aminoácidos, péptidos aniónicos o catiónicos y carbohidratos; la parte hidrofóbica regularmente está constituida de ácidos grasos saturados o insaturados. Los biotensoactivos más estudiados han sido los glicolípidos, lipopéptidos y lipoproteínas, fosfolípidos y ácidos grasos, tensoactivos poliméricos y tensoactivos particulados (Desai y Banat 1997). Los glicolípidos son los biotensoactivos más comunes; están formados por mono, di, tri y tetrasacáridos en combinación con una o más cadenas de ácidos alifáticos o ácidos hidroxialifáticos.
Los glicolípidos pueden subclasificarse en ramnolípidos, trehalolípidos y soforolípidos. Los ramnolípidos están constituidos por mono y disacáridos de ramnosa ligados por un enlace glicosídico a una molécula de β–hidroxidecanoil–β–hidroxidecanoato (Fig. 5a). Los trehalolípidos están constituidos por el disacárido trealosa, en el que a cada unidad de glucosa se encuentra ligado un ácido graso por medio de un enlace α–1–glicosídico (Fig. 5b). La estructura de los soforolípidos tiene como base al disacárido soforosa (β–D–Glc–(1,2)–D–Glc), el cual se encuentra ligado glicosídicamente al grupo 17–hidroxilo, ya sea a una molécula de ácido esteárico (ácido graso saturado de 18 carbonos) o a una molécula de ácido oleico (ácido insaturado cis–9–octadecaenoico). Dos características estructurales destacan en los soforo–lípidos: el grupo carboxilo está usualmente ligado a la segunda unidad de glucosa mediante el grupo 4'–hidroxilo para formar una lactona; por otra parte, los grupos 6–hidroxilo de cada unidad de glucosa pueden o no encontrarse acetilados (Fig. 5c).

Dentro de las estructuras de tipo lipopéptido, con significativa capacidad biotensoactiva, se tiene a los lipopéptidos cíclicos tales como la gramicidina S y las polimixinas (Fig. 6a y 6b). Estas dos clases de biotensoactivos poseen actividad antibacteriana debido a su capacidad de solubilizar enzimas de membrana. Otro biotensoactivo perteneciente a este grupo y reconocido como uno de los más poderosos es la surfactina (Fig. 6c), que en concentraciones menores del 0.005 % en peso reduce la tensión superficial del agua de 72 a 27.9 N/m (Arima et al. 1968).
Los biotensoactivos poliméricos están constituidos por diversos grupos químicos. Un ejemplo es el emulsán (Fig. 7), en el cual los ácidos grasos se encuentran ligados a un esqueleto de heteropolisa–cáridos (Rosenberg et al. 1979). Otro biotensoactivo polimérico es el liposán, cuya estructura está constituida por 83 % de carbohidratos y 17 % de proteínas (Cirigliano y Carman 1984). Las vesículas y células microbianas con activad tensoactiva son denominadas biotensoactivos particulados. Ejemplos de microorganismos con una elevada hidrofobicidad superficial son las cianobacterias y algunos microorganismos patógenos del género Serratia.
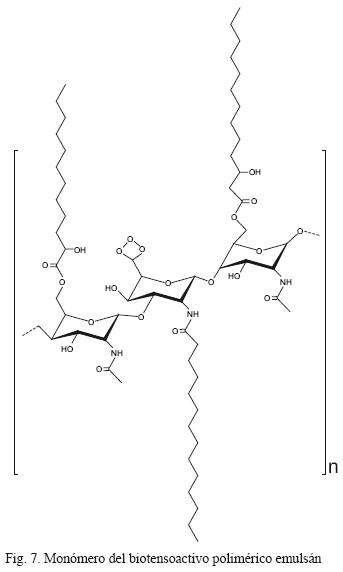
Las propiedades de na, HBL y PN no son propiedades reportadas para biotensoactivos. Para el caso de PN no se reportan debido a que la mayoría de los biotensoactivos son estables a temperaturas extremas.
Biotensoactivos: funciones microbianas y microorganismos productores
Varios tipos de microorganismos, incluyendo bacterias, levaduras y hongos, poseen un metabolismo capaz de producir biotensoactivos (Correa et al. 1999). Sin embargo, la mayoría de los biotensoactivos son producidos por bacterias (Cuadro V).

Los microorganismos utilizan una amplia variedad de compuestos orgánicos como fuentes de carbono y energía para su crecimiento. Cuando estos sustratos son inaccesibles por su baja solubilidad, competencia con otros microorganismos u otro tipo de factor ambiental, utilizan a los biotensoactivos para disponer de la fuente de carbono. Las principales funciones de los biotensoactivos a nivel fisiológico son:
• Capacidad para emulsificar y solubilizar compuestos insolubles en fases acuosas, como son los hidrocarburos, facilitando la disponibilidad de estos sustratos para el crecimiento y mantenimiento de los microorganismos (Francy et al. 1991).
• Adherencia y liberación de células en superficies. Ésta es una de las más importantes estrategias de supervivencia de los microorganismos debido a la presencia de biotensoactivos en la parte externa de la membrana o pared celular. Estos biotensoactivos son responsables de regular las propiedades de la superficie celular, promoviendo la adherencia o liberación de las células de sitios específicos de acuerdo a sus necesidades nutricionales o para evadir ambientes desfavorables (Rosenberg et al. 1989).
• Facilitar el transporte de hidrocarburos a nivel de membrana, función atribuida a los biotensoactivos del tipo glicolipídico ligados a membrana celular.
• Actividad antibiótica, la que ha sido demostrada por varios biotensoactivos, principalmente los lipopéptidos y glicopéptidos. Ejemplos de éstos son los ramnolípidos de Pseudomonas aeruginosa y la surfactina de Bacillus subtilis, los cuales presentan actividad como antibióticos, lo que les permite solubilizar los principales componentes de las membranas celulares microbianas y les brinda una mayor oportunidad de supervivencia en nichos con una alta competitividad por nutrientes (Lin 1996).
La principal función biológica de los biotenso–activos es su capacidad para solubilizar sustratos hidrofóbicos tales como hidrocarburos, lípidos, aceites, fármacos y otros (Morikawa et al. 1993, Zeng et al. 2005).
Ramnolípidos producidos por Pseudomonas spp.
Diversas especies de Pseudomonas destacan por su capacidad para producir ramnolípidos (Guerra–Santos et al. 1984, Cooper et al. 1989, Cameotra y Singh 1990, Deziel et al. 1996). Bergström et al. (1946) fueron los primeros en reportar la estructura de un glicolípido sintetizado por Pseudomonas pyocyanea, conformado por ramnosa y ácido β–hidroxidecanoico. Tres años después, Jarvis y Jonson (1949), empleando medios de cultivo con 3 % de glicerol, también aislaron ramnolípidos provenientes de Pseudomonas aeruginosa; determinaron que estos compuestos estaban formados por un glucósido de β–hidroxidecanoil–β–hidroxidecanoato y dos moléculas de ramnosa. Cabe mencionar que antes de elucidar con certeza la estructura de los ramnolípidos, en la década de 1950 ya eran usados en procesos de biorremediación de suelos contaminados con aceites y metales pesados (Hemminger 2005).
La primera estructura claramente elucidada de los ramnolípidos fue encontrada por Edwards y Hayashi (1965), quienes demostraron la unión glicosídica del disacárido de ramnosa al β–hidroxidecanoil–β–hidroxidecanoato y cuya estructura se ha denominado como R2 (Fig. 5a derecha). Esta misma estructura también fue observada por Hisatsuka et al. (1971), utilizando P. aeruginosa S7B1 cultivada en hexadecano y parafinas. Un año después, Itoh y Suzuki (1972) encontraron que P. aeruginosa KY 4025 cultivada con 10 % de n–alcanos produce el mono–ramnolípido correspondiente a L–α–ramnopiranosil–β–hidroxidecanoil–β–hidroxidecanoato, estructura a la cual se ha denominado R1 (Fig. 5a izquierda). Otros ramnolípidos similares a los mono– y disacáridos de ramnosa fueron caracterizados por Syldatk et al. (1985). Estos tipos de ramnolípidos se caracterizan por contener sólo una unidad de ácido β–hidroxidecanoico y se les ha denominado R3 y R4 respectivamente (Fig. 8).

En lo referente a la fuente de carbono, se ha reportado que la producción de ramnolípidos por el género Pseudomonas es estimulada por diferentes tipos de sustratos insolubles, los cuales van desde hidrocarburos alifáticos y aromáticos hasta aceites de origen vegetal (Rashedi et al. 2005). Dependiendo de la fuente de carbono utilizada, se ha visto que la CMC para ramnolípidos presenta valores diferentes debido a diferentes estructuras químicas dependiendo del ramnolípido formado. Nitschke et al. (2005b) han reportado valores de la CMC para ramnolípidos biosintetizados por Pseudomonas aeruginosa a partir del aceite de diferentes tipos de semillas: algodón (CMC = 86.79 mg/L), maíz (CMC = 43.21 mg/L), soya (CMC = 54.56 mg/L) y palma (CMC = 40.19 mg/L). Sin embargo, cabe destacar que no cambian significativamente los valores de la tensión superficial mínima alcanzada a estos valores de CMC para ramnolípidos biosintetizados de las diferentes fuentes de carbono, manteniéndose en promedio a 30.86 N/m.
La vía metabólica que interviene en la biosíntesis de los ramnolípidos está estrechamente ligada a la ruta de biosíntesis de ácidos grasos y también se relaciona con moléculas que contienen L–ramnosa o sus precursores, tales como el alginato (Maier y Soberón–Chávez 2000). En esta ruta se ha encontrado que la enzima ramnosiltransferasa 1 (RhlA) no sólo participa en la transferencia de una molécula de ramnosa de la dTDP–L–ramnosa a la parte lipídica de los ramnolípidos, sino que también es responsable de la síntesis del dímero de ácidos grasos que forma la molécula (Soberón–Chávez et al. 2005).
Otros biotensoactivos y sus microorganismos productores
La formación de trehalolípidos ha sido reportada en diferentes géneros de bacterias tales como Myco–bacterium, Nocardia, Corynebacterium y Brevibacteria (Asselineau y Asselineau 1978, Desai y Banat 1997). Sin embargo, los trehalolípidos provenientes de cada una de estas cepas diferen en tamaño y en la estructura de los ésteres de los ácidos micólicos que los integran debido a diferencias en el metabolismo de estos géneros. La formación de soforolípidos por levaduras de Torulopsis bombicola ATCC 22214 ha sido reportada por Cooper y Paddock (1984), quienes –al cultivar la levadura por 4 días en un medio con glucosa (4 %), extracto de levadura (1 %) y complementado con elementos traza– encontraron un decremento de la tensión superficial del medio de cultivo hasta de 32.7 N/m en promedio.
Microorganismos como Acinetobacter spp. y Thio–bacillus thiooxidans producen fosfolípidos. En el caso de Acinetobacter spp., cuando se cultivan con alcanos como sustratos, el nivel de fosfolípidos de membrana se incrementa (Makula et al. 1975). Por su parte, Thio–bacillus thiooxidans sintetiza y utiliza fosfolípidos para solubilizar sulfuro elemental, el cual es necesario para su crecimiento (Van Hamme et al. 2003).
Los biotensoactivos lipopéptidos, como la gramicidina S y las polimixinas, son producidos por microorganismos del género Brevibacterium. En lo referente a la gramicidina S, es producida por Brevibacterium brevis, que se distingue por contener cantidades de este antibiótico unidas fuertemente en la superficie exterior de sus esporas, sin que se sepa en este momento cómo actúa preferentemente (Marahiel et al. 1977).
Diferentes especies del género Bacillus producen biotensoactivos del tipo lipopéptido. Las especies Bacillus subtilis y Bacillus licheniformis producen surfactina, que probablemente sirve para dispersar las esporas dentro de su ciclo biológico. Las cepas denominadas F2.2 de Bacillus licheniformis, aisladas de alimentos fermentados provenientes de Tailandia, producen un biotensoactivo similar a la surfactina, denominado plipastatina (Thaniyavarn et al. 2003). Para el caso de los favolípidos, biotensoactivos producidos por cepas de Flavobacterium sp., han mostrado la capacidad de un efectivo agente emulsificante en sistemas con hexadecano (Bodour et al. 2004). Morikawa et al. (1993), determinaron que la cepa MIS38 de Arthrobacter sp. produce un biotenso–activo de tipo lipopéptido denominado artrofactina, el cual reduce la tensión superficial del medio de cultivo por debajo del valor medio (24 N/m). Myxococcus xanthus produce un biotensoactivo lipopeptídico con actividad antibiótica denominado TA, que inhibe la síntesis de peptidoglucano al interferir en su polimerización (Zafriri et al. 1981). Ferraz et al. (2002), utilizando Serratia marcescens, produjeron biotensoactivos (serrawetina) con aceites vegetales como sustratos, encontrando un decremento en la tensión superficial de 64.54 a 29.57 N/m.
La gran variedad de microorganismos utilizados en la biosíntesis de biotensoactivos es parte de la búsqueda de biocatalizadores que disminuyan los costos asociados a su producción, permitiéndoles ser competitivos en un futuro con los tensoactivos sintéticos. Dicha competencia se dará al considerar que los biotensoactivos no son tóxicos y son biodegradables y que de acuerdo a las nuevas legislaciones se deberán hacer tratamientos con compuestos de esta naturaleza. Además, la optimización de las variables de proceso, la utilización de materiales baratos o de desecho y el uso de los microorganismos adecuados, permitirán que la producción de biotensoactivos sea un proceso económicamente rentable.
Aplicaciones de los biotensoactivos
La aplicación práctica de los biotensoactivos está limitada por sus costos de producción y propiedades como agentes de superficie en relación a la competencia con los tensoactivos sintetizados químicamente (Georgiou et al. 1992). Los biotensoactivos tienen una aplicación potencial tanto en sectores ambientales y de recuperación de crudo como en el área agrícola, farmacéutica, de alimentos y cosméticos.
Una aplicación directa de los biotensoactivos está basada en su actividad antimicrobiana, la cual es estimada de acuerdo a los valores de la menor concentración inhibitoria como agente antimicrobiano (MIC por sus siglas en inglés: Minor Inhibitory Concentration). Algunas MIC de los ramnolípidos de Pseudomonas aeruginosa AT10 contra algunos microorganismos son: Aspergillus niger, 16 μg/ mL; Candida albicans, 256 μg/mL; Streptoccocus faecalis, 64 μg/mL; Salmonella thyphimurium, 128 μg/mL y Staphyloccocus epidermi, 8 μg/mL (Abalos et al. 2001).
En la agricultura, los biotensoactivos son aplicados debido a su actividad insecticida y herbicida. Ramnolípidos producidos por Pseudomonas syringae son usados en el biocontrol de Canada thistle. Los ramnolípidos junto con el hongo Myrothecium verrucaria también han sido empleados de manera simultánea como herbicidas en el control de Pueraria lobata (Singh et al. 2006). Biotensoactivos producidos por bacterias del género Bacillus han sido ocupados para emulsificar y dispersar en fase acuosa pesticidas de solubilidades bajas como son los de tipo organoclorado (Patel y Gopinathan 1986).
A nivel industrial, los heteropolisacáridos de Macrocystis pirifera y Azotobacter vinelandii han sido utilizados como dispersantes en la producción de cerámica (Pellerin et al. 1991).
El espectro de aplicación de los biotensoactivos es muy amplio, e incluso han encontrado un uso en el campo de la biorremediación y mejoramiento en la extracción de crudo. Ejemplo de esta actividad son los productos utilizados por la empresa Rhamnolipods Co., que utiliza algunos productos para remediación como son Rhamnolipids Biorem 80, Esludger 60 y Biorem 70 ®.
Biorremediación
La biorremediación se define como el proceso por el cual los desechos orgánicos son biológicamente removidos o degradados bajo condiciones controladas a un estado inocuo, o a niveles de menor concentración de acuerdo a los límites establecidos por las autoridades reguladoras (Mueller et al. 1992, Vidali 2001). Cabe destacar que la biodegradación de un componente es comúnmente el resultado de la interacción de múltiples microorganismos.
La biorremediación es eficiente sólo cuando las condiciones del ambiente permiten el crecimiento y actividad enzimática de los microorganismos. Esta aplicación a menudo involucra la manipulación de parámetros para permitir el crecimiento microbiano y el proceso de degradación a una mayor velocidad, como son agitación, tamaño de inóculo, temperatura de crecimiento, relaciones C/N, etc. (Vidali 2001). El método empleado para llevar a cabo un proceso de biorremediación está determinado por el costo del tratamiento. En el cuadro VI son comparados algunos de estos procedimientos y su relación costo/ beneficio (Kosaric 2001).

Uno de los problemas ecológicos y sociales de mayor impacto, atendido por las tecnologías de bioremediación, ha sido el ocasionado por derrames de petróleo y sus derivados en medios acuosos, suelos y sedimentos. Esta problemática ha conducido a la remediación de estos sitios mediante la aplicación de biotensoactivos o las cepas de los microorganismos que los producen. En este punto, cabe mencionar que los tensoactivos químicos también han sido utilizados con el mismo propósito; sin embargo, al ser compuestos xenobióticos, presentan problemas importantes por su biodegradación y toxicidad. Un ejemplo claro son los nonilfenoletoxilatos como el tritón X–100, un tensoactivo químico no iónico cuyas cadenas etoxílicas han mostrado ser biodegradables por procesos aerobios y anaerobios. No obstante, la estructura nonilfenólica es altamente recalcitrante y tóxica, teniendo un efecto estrogénico en diversos organismos superiores (peces, reptiles, aves y humanos) y a diferentes niveles tróficos. Esto se debe a que interfiere con el sistema endocrino, pues su estructura es similar al β–estradiol, precursor de diferentes tipos de estrógenos (Gross et al. 2004). Este tipo de problemática hace de los biotensoactivos candidatos potenciales para reemplazar a los tensoactivos químicos en el futuro (Banat et al. 2000).
Cuando el petróleo es derramado en sistemas acuosos, la solubilización y volatilización son mecanismos abióticos que actúan inicialmente, removiendo a los hidrocarburos polares y de bajo peso molecular. Sin embargo, la mayor parte de los hidrocarburos presentes en el petróleo son compuestos que, comparados con el agua, poseen una menor densidad, mayor peso molecular y una solubilidad baja. Por tal razón, después de un derrame, la mayoría de los hidrocarburos permanece en la superficie acuosa. Diversos microorganismos productores de biotensoactivos y degradadores de hidrocarburos se encuentran presentes en aguas marinas, por lo que la biodegradación resulta uno de los métodos más eficientes para la remoción de estos contaminantes (Atlas 1993).
Se ha demostrado que en sistemas acuosos donde existen compuestos hidrofóbicos, la adición de biotensoactivos incrementa su solubilidad. Por ejemplo, la solubilidad del octadecano en agua es de 0.006 mg/L, pero la adición de ramnolípidos la incrementa hasta cerca de 250 mg/L, donde el aumento en la solubilidad está asociado al rápido decremento en la tensión superficial (Zhang y Miller 1992). Este comportamiento es corroborado por una extensa cantidad de trabajos. Wouther y Dick (2002) reportaron que la solubilidad de diferentes tipos de alcanos de cadena larga se incrementa al utilizar ramnolípidos como agentes tensoactivos, lo que a su vez condujo a un aumento en las tasas de biodegradación. Tang et al. (2007), durante un proceso de biodegradación de crudo, obtuvieron un decremento de tensión superficial de 50 a 35 N/m en 24 horas, alcanzando la máxima reducción a las 96 horas (32 N/m), con una biodegradación del compuesto de hasta 60 %. Chakrabarty (1985) fue de los primeros en reportar a una cepa (SB30) de Pseudomonas aeruginosa como microorganismo aislado de aguas marinas, capaz de producir un ramnolípido que dispersa rápidamente hidrocarburos en gotas muy pequeñas, lo que ha hecho de este microorganismo uno de los principales agentes bióticos para la remedición de costas contaminadas con hidrocarburos.
Otra aplicación importante de microorganismos productores de biotensoactivos en sistemas acuosos ha sido en la limpieza de tanques de almacenamiento de crudo (Banat et al. 1991). El biotensoactivo emulsán ha sido empleado en sistemas petróleo–agua para la limpieza de buques tanque, reducción en la viscosidad de petróleo pesado, mejoramiento en la recuperación de crudo y estabilización de emulsiones en combustibles (Banat et al. 1991, Banat 1995).
En los procesos de biorremediación de suelos contaminados con hidrocarburos, también ha sido extenso el uso de biotensoactivos (surfactina, ramnolípidos y otros) y los microorganismos que los producen (Bacillus subtillis, Pseudomonas aeruginosa y Pseudomonas putida, entre otros) (Atlas 1991, Dave et al. 1994, Ghosh et al. 1995). La degradación de los hidrocarburos contaminantes de suelos y sedimentos depende de diferentes factores, como la presencia de microorganismos degradadores, la composición de los hidrocarburos, la actividad del agua, el pH, la temperatura y la disponibilidad de nutrientes inorgánicos y oxígeno. Además de los anteriores, se debe considerar la dependencia de las interacciones fsicoquímicas que se establecen entre los hidrocarburos con la matriz porosa y la fracción de materia orgánica de suelos y sedimentos. Debido a la naturaleza de la matriz porosa del suelo y a su contenido de materia orgánica, los hidrocarburos de mayor peso molecular y menor solubilidad en agua tienden a intemperizarse en los microporos de las partículas del suelo, siendo inaccesibles como fuentes de carbono y energía para los microorganismos. Ante esta situación, los biotensoactivos actúan incrementando la biodisponibilidad de los contaminantes mediante la acción paralela de un fenómeno de desorción y solubilización, desplazando preferentemente el equilibrio de reparto de los hidrocarburos hacia la fase acuosa al interior de los microporos, lo que permite que se lleven a cabo los procesos de transferencia de masa y biodegradación (Fig. 9).
Diversos estudios han demostrado que el uso de biotensoactivos en suelos y sedimentos contaminados con hidrocarburos policíclicos aromáticos, conduce a un aumento en la biodisponibilidad y mineralización de este tipo de compuestos, tales como el naftaleno y fenantreno (Deziel et al. 1996). Oberbremer y Müller–Hurtig (1989), utilizando poblaciones mixtas provenientes de suelos contaminados y mezclas de hidrocarburos aromáticos, observaron un consumo inicial de naftaleno, seguido de una producción de biotensoactivos, reflejada por la disminución de la tensión superficial del medio junto con la degradación de los demás hidrocarburos componentes de la mezcla. Foght et al. (1989) reportan que el emulsán estimula la mineralización de compuestos aromáticos en el suelo por cultivos de bacterias puras, pero inhibe el proceso de degradación cuando se emplean cultivos mixtos. Por otra parte, la adición de soforolípidos incrementa tanto la magnitud en la degradación de hidrocarburos contaminantes del suelo, como el rendimiento final de biomasa (Oberbremer et al. 1990). Berg et al. (1990) utilizaron ramnolípidos provenientes de la cepa UG2 de Pseudomonas aeruginosa y encontraron un incremento en la solubilidad del hexaclorobifenilo presente en sedimentos, la cual condujo a una recuperación de 31 % del compuesto en fase acuosa. Esta recuperación fue tres veces mayor a la solubilización del mismo compuesto empleando el tensoactivo químico aniónico ligninsulfonato de sodio.
Los tensoactivos también se emplean en procesos de biodegradación de pesticidas y remoción de metales, sin embargo, este es un campo relativamente nuevo. En el caso de los pesticidas, los estudios se enfocan principalmente a los de tipo organoclorado, como son el DDT y los isómeros del hexa–cloro–ciclohexano (Karanth et al. 1999, Karanth et al. 2000).
Mejoramiento microbiano para la recuperación de petróleo (MEOR)
Dentro de la industria petrolera, una de las aplicaciones potenciales de los microorganimos productores de biotensoactivos es el mejoramiento en la recuperación de petróleo, la cual, por sus siglas en inglés, es denominada MEOR (Microbial Enhanced Oil Recovery). Este proceso es una tecnología insitu de recuperación terciaria que utiliza microorganismos productores de biotensoactivos con el objetivo de reducir las tensiones de las interfases presentes entre el petróleo, la superficie sólida del material rocoso y la fase acuosa al interior de los poros. Al disminuir estas fuerzas, se mejora la fluidez de los hidrocarburos a través de los poros de la roca, promoviendo así la recuperación de crudo residual.
Existen diversos procedimientos para el uso de microorganismos productores de biotensoactivos en los procesos de MEOR. El más empleado consiste en la inoculación de los microorganismos mediante inyección directa en los depósitos de petróleo, a los que se agregan nutrientes inorgánicos tales como fertilizantes junto con sustratos de bajo costo como las melazas. Este procedimiento se lleva a cabo con la finalidad de promover el crecimiento microbiano y la producción de los biotensoactivos (Banat 1995).
Para que un proceso MEOR sea factible, es necesario que los microorganismos utilizados sean extremóflos, es decir, que sean capaces de crecer y producir biotensoactivos bajo condiciones ambientales severas, tales como tensión de oxígeno baja, temperatura, presión y alta concentración salina (Karanth et al. 1999). Diversos microorganismos termóflos, aerobios y anaerobios, tolerantes a salinidad y presión moderadas, con capacidad de fuidificar petróleo a nivel laboratorio, han sido aislados y estudiados (Janneman et al. 1983). Debido a los altos incrementos del crudo, estos estudios, junto con los reportes hechos por diversas compañías petroleras, hacen del proceso MEOR una herramienta fundamental para la explotación más eficiente del petróleo en el futuro (Banat 1995).
Otros nichos potenciales de la aplicación de biotensoactivos
Debido a su poder para formar emulsiones y a su baja o nula toxicidad, los biotensoactivos también han encontrado un nicho potencial en el área de alimentos y cosméticos (Banat et al. 2000).
La aplicación de los tensoactivos en la industria de cosméticos y cuidado de la salud es diversa, por ejemplo: repelentes, insecticidas, antiácidos, toallas antiacné, champú anticaspa, soluciones de lentes de contacto, desodorantes, colorantes, mascarillas, limpiadores dentales, antisépticos, entre otros. A pesar de que el uso de biotensoactivos en la industria alimenticia y de cosméticos posee mayores ventajas que los tensoactivos químicos, estos últimos siguen predominando en estas áreas. Una de las principales ventajas de los biotensoactivos es que son compatibles con la piel, ya que no son irritantes (Kleckner y Kosaric 1993).
En el sector alimenticio han sido utilizados en la estabilización de materias primas, la solubilización de aromas (Banat 2000) y como modificadores de las características reológicas de productos derivados de la carne (Vater 1986). En el sector de los cosméticos, los soforolípidos son los biotensoactivos más utilizados (Yamane 1987). En combinación con aditivos como el propilenglicol y las lipasas, están siendo empleados como hidratantes en productos para piel como cremas y maquillajes, y para productos del cabello (Desai y Banat 1997).
Otra aplicación de los biotensoactivos es en el área farmacéutica. En este sector destacan los lipopéptidos surfactina e iturina producidos por Bacillus subtilis, los cuales además de poseer actividad antibacteriana y antifúngica, también han mostrado actividad antitumoral y antivirial, así como efectividad en inhibir la formación de coágulos (Arima et al. 1968, Thimon et al. 1995, Peypoux et al. 1999). Otro ejemplo en esta área es el biotensoactivo producido por Rhodo–coccus erythropolis (lípidos de succinoltrealosa), el cual inhibe tanto al herpes simple como a una cepa del virus de la influenza (Uchida et al. 1989). Ejemplo claro de esto es la utilización de los ramnolípidos en cosméticos en Japón (Maier y Soberón–Chávez 2000).
Además de las aplicaciones anteriores, los biotensoactivos también han encontrado nichos en industrias tales como la metalúrgica (espumantes, lubricantes y anticorrosivos), de productos de limpieza (agentes espumantes y limpiadores), papelera (dispersante y lavado), pinturas (emulsiones, dispersión de pigmentos, estabilizador de caucho y retardador de sedimentación), textil (detergente, emulsificante y dispersante) y de plásticos (solubilizador y emulsificador de monómeros) (Kosaric 2001).
Producción de biotensoactivos
La viabilidad económica en la producción a gran escala de diversos metabolitos mediante procesos biotecnológicos suele tener como cuello de botella las etapas de biosíntesis o purificación. En el caso particular de los biotensoactivos, se ha determinado que la biosíntesis es el paso limitante para su producción a gran escala. Para tener un proceso de producción de biotensoactivos económicamente competitivo, es recomendable que se den los siguiente factores (Mukherjee et al. 2006): i) el uso de sustratos baratos o de desecho que disminuyan los costos del proceso, ii) desarrollo de bioprocesos eficientes, incluida la optimización de las condiciones de cultivo y procesos de separación rentables para la máxima producción y recuperación de biotensoactivos y iii) desarrollo y uso de cepas sobreproductoras, mutantes o recombinantes, para aumentar los rendimientos. Para cualquiera de estos tres factores es necesario un entendimiento de los mecanismos de regulación y de las cinéticas de producción con base en el microorganismo y sustrato elegidos (Nitschke et al. 2005a).
Los biotensoactivos pueden sintetizarse a partir de diversos sustratos, sin embargo, se está dando una gran importancia a los provenientes de recursos renovables como son aceites vegetales, desechos de destilerías y lecherías, que son baratos, pero no han sido reportados a detalle (Makkar y Cameotra 2002).
Varios elementos y precursores presentes en los medios de cultivo para la síntesis de biotensoactivos han reportado un efecto sobre sus propiedades y estructura, así como en la eficiencia de los procesos para su producción. Elementos traza como hierro y manganeso modifican el rendimiento del biotensoactivo (Mukherjee et al. 2006). Otro ejemplo es la limitación de nitrógeno, la cual conduce a un incremento en la producción del biotensoactivo, siendo el caso de los ramnolípidos provenientes de Pseudomonas aeruginosa (Guerra–Santos et al. 1984). Otros estudios se han enfocado en la optimización de medios al probar diferentes fuentes de carbono y nitrógeno, así como niveles de concentración, con el propósito de mejorar la producción de biotensoactivos (Santa Anna et al. 2002, Rashedi et al. 2005).
Efecto de la fuente de carbono
La fuente de carbono influencia las rutas de síntesis de biotensoactivos por inducción o represión. En la mayoría de los casos, la adición de sustratos inmiscibles en agua, como alcanos y otros tipos de hidrocarburos, promueve la inducción en la producción del biotensoactivo. La represión catabólica en la síntesis de biotensoactivos por glucosa, acetato y ácidos tricarboxílicos también ha sido observada (Nitschke et al. 2005a).
Un efecto de la fuente de carbono en el biotensoactivo producido fue observado por Robert et al. (1989), que utilizando como fuentes de carbono glicerol, manitol, aceite de oliva, fructosa, succinato y piruvato con P. aeruginosa 44T1, produjeron ramnolípidos, con reducciones de la tensión superficial del medio de 72 N/m hasta 30, 28.5, 28.4, 31.7, 29.6 y 32.9 N/m, respectivamente. Otras fuentes de carbono utilizadas para la producción de biotensoactivos por Pseudomonas aeruginosa han sido aceite de soya, n–hexadecano, aceite de maíz, efluentes de la producción de aceite de girasol y etanol entre otras (Haba et al. 2000, Nitschke et al. 2005b). De las fuentes de desecho que se han utilizado para la producción de biotensoactivos destaca el suero de la industria del queso, que se ha empleado para el crecimiento de P. aeruginosa en la producción de ramnolípidos en estado cristalino (Dubey y Juwarkar 2001).
Bodour et al. (2004) obtuvieron una nueva clase de biotensoactivo: los favolípidos. Cultivaron cepas de Flavobacterium sp. en un medio mineral con 2 % de glucosa como fuente de carbono y energía, logrando reducir la tensión superficial del medio hasta 26 N/m, siendo la inicial la del agua.
Javaheri et al. (1985) produjeron biotensoactivos a partir de cepas de Bacillus licheniformis JF–2, creciendo este microorganismo en un medio mineral complementado con glucosa; observaron una reducción de la tensión superficial de 70 a 28 N/m. Cultivada con glucosa, B. licheniformis BAS–50 produce liquenisina A, una mezcla de lipopéptidos que destaca por su alta actividad antibacterial y es capaz de disminuir la tensión superficial hasta valores cercanos a 28 N/m (Yakimov et al. 1995). Bacillus subtilis produce surfactina con sacarosa como fuente de carbono (Makkar y Cameotra 1998). Torulopsis bombicola produce soforolípidos con altos rendimientos al usar azúcares y aceites vegetales como sustratos (Kosaric 2001). Candida sp. SY16 produce glicolípidos que reducen la tensión superficial hasta 30 N/m al cultivarse con aceite de soya (Kim et al. 2006).
Fuentes de nitrógeno y fósforo
Se ha mostrado que el nitrato de sodio es más eficiente que el sulfato de amonio y urea en la producción de ramnolípidos por Pseudomonas aeruginosa. Tanto Guerra Santos et al. (1984) como Rashedi et al. (2005), evaluando la fuente de nitrógeno en la producción de ramnolípidos con P. aeruginosa y glucosa como fuente de carbono, determinaron que los mayores rendimientos se alcanzan al emplear nitrato de sodio respecto al sulfato de amonio a una relación carbono/nitrógeno (C/N, mol) de 18. Otros investigadores han reportado valores similares en los que la relación C/N con nitrato de sodio debe encontrarse en un intervalo óptimo de 15 a 23 con glucosa y aceites vegetales como fuentes de carbono (Ochsner et al. 1996, Santa Anna et al. 2002). También se ha reportado que el nitrato de sodio y potasio son consumidos preferentemente por Bacillus subtilis al cultivarse con sacarosa y almidón, alcanzando una mayor reducción en la tensión superficial con respecto al uso de sulfato de amonio, urea y nitrato de amonio (Makkar y Cameotra 1998). El incremento en la producción de ramnolípidos por especies de Pseudomonas muy probablemente se debe a una inhibición de la glutamato sintetasa por NH4+, glutamina, asparagina y metionina como fuentes de nitrógeno. Se ha reportado que esta enzima es responsable de la disminución en la producción de ramnolípidos cuando se utiliza amonio y compuestos afines como fuente de nitrógeno, mientras que su actividad aumenta con la presencia de nitrato en el medio (Venkata y Karanth 1989, Köhler et al. 2000).
Otro elemento importante para la biosíntesis de tensoactivos es el fósforo. Guerra Santos et al. (1984) evaluaron la fuente de fósforo en un intervalo carbono/fósforo (C/P, mol) de 10 a 32, con glucosa como fuente de carbono. La máxima producción del biotensoactivo fue obtenida con una relación C/P = 16. Rashedi et al. (2006) evaluaron la relación C/P, alcanzando la máxima producción de ramnolípidos con una relación C/P = 20, utilizando Pseudomonas aeruginosa MM1011 como cepa productora.
Iones y elementos traza
Las limitaciones en la concentración de iones multivalentes de Mg+2, Ca+2, K+, Na+ y otros elementos traza, conducen a mejores rendimientos en la producción de ramnolípidos por Pseudomonas aeruginosa DSM 2659 (Nitschke et al. 2005a). Concentraciones altas de iones en la fase acuosa inhiben la producción de biotensoactivos al afectar sus propiedades de interfase; por ejemplo, altas concentraciones de cloruro de sodio inactivan a los glicolípidos de Torulopsis apicola (Georgiou et al. 1992). Sin embargo, también se ha reportado la producción de biotensoactivos como la liquenisina A, a partir de Bacillus licheniformis BAS–50, en medios con concentraciones mayores a 13 % de cloruro de sodio (Yakimov et al. 1995).
El hierro también es un elemento traza que incide en la síntesis de los biotensoactivos. Se ha reportado un incremento en la producción de biotensoactivos por Pseudomonas putida con una relación carbono/ hierro (C/Fe mol:mol) igual a 26,000. Sin embargo, la reducción de la tensión superficial en el medio no ha mostrado cambios significativos con respecto a otras relaciones C/Fe (Amézcua et al. 2004). Rashedi et al. (2006) evaluaron la relación C/Fe, encontrando que era necesario limitar la concentración de hierro a 27.5 μg de FeSO4 7H2O por gramo de glucosa en el medio. Bacillus subtilis también mejora significativamente la producción de biotensoactivos al remover la espuma formada en el cultivo y adicionar sales de hierro y manganeso al medio (Cooper et al. 1981).
Efecto de las condiciones de cultivo
Las condiciones de cultivo tales como temperatura, pH, agitación y disponibilidad de oxígeno, también han mostrado un efecto sobre la producción del biotensoactivo. Robert et al. (1989) encontraron que la mejor temperatura para la formación de biotensoactivo es de 37 °C para Pseudomonas aeruginosa 44T. Bacillus subtilis crece y produce surfactina en diferentes fuentes de carbono y de nitrógeno, bajo condiciones termofílicas y mesofílicas a 45 y 30 °C respectivamente. En estos experimentos, después de 96 h de crecimiento a 45 °C y con almidón como fuente de carbono, la tensión superficial disminuyó de 68 a 32 N/m. Bajo las mismas condiciones, una mayor reducción de tensión superficial fue alcanzada en condiciones mesofílicas (Makkar y Cameotra 1998).
Yateem et al. (2002) reportan que a 35 °C dos especies de Pseudomonas alcanzan su máximo de crecimiento y producción de biotensoactivo a valores de pH neutros, y que su capacidad decrece conforme el medio se va haciendo alcalino. Pruthi y Cameotra (2003), utilizando Pseudomonas putida, alcanzaron la máxima concentración de biomasa y producción de biotensoactivo a pH entre 6.4 y 7.2 y temperatura entre 30 y 40 °C. Señalan que el rendimiento más alto se presenta en la etapa de estado estacionario, sugiriendo que el biotensoactivo es un metabolito secundario. Tarek et al. (2006) optimizaron la producción de biotensoactivo a partir de Nocardia amarae con aceite de oliva como fuente de carbono; reportan que los cultivos agitados tienen un mayor rendimiento que los cultivos estáticos, alcanzando el mayor rendimiento a 150 rpm. Chayabutra et al. (2001) determinaron que la producción de ramnolípidos a partir de Pseudomonas aeruginosa es mayor bajo condiciones aerobias que desnitrificantes, pero que la biosíntesis mediante desnitrificación podría ser una ruta factible debido a la menor generación de espuma en el cultivo. La producción de biotensoactivos con extracto de levadura en el medio de cultivo, también se ha visto empobrecida debido a que conduce a un metabolismo anabólico con alta producción de biomasa, disminuyendo con esto los rendimientos en la producción de ramnolípidos en especies de Pseudomonas (Guerra Santos et al. 1984, Rashedi et al. 2006).
CONCLUSIONES
Los tensoactivos químicos son compuestos anfifílicos cuyo potencial y caracterización en su capacidad de emulsión y como agentes de superficie, están determinados de manera individual por cuatro parámetros importantes (CMC, na, HLB y PN). Sin embargo, los biotensoactivos muestran un potencial mayor sobre los tensoactivos químicos para aplicaciones a nivel industrial y ambiental; el primero de ellos es su inocuidad con el ambiente asociada a una toxicidad prácticamente nula y a una fácil biotransformación. El segundo es que –a diferencia de los tensoactivos químicos, sintetizados a partir de fuentes no renovables como el petróleo– los biotensoactivos pueden biosintetizarse a partir de desechos y fuentes renovables como son aceites y grasas, ya sean de origen vegetal o animal. El tercero es la termoestabilidad que le permite mantener su capacidad tensoactiva. No obstante, la factibilidad en el uso práctico de los biotensoactivos está fuertemente limitada por sus altos costos de producción y purificación. En particular, la producción de ramnolípidos a partir de Pseudomonas aeruginosa y otras especies del mismo género está significativamente influenciada por el tipo y concentración de nutrientes como las fuentes de carbono, nitrógeno, fósforo y la presencia de ciertos elementos traza, así como de las condiciones de cultivo tales como pH, temperatura, agitación y tensión de oxígeno. Por lo anterior, la investigación a nivel básico de cómo inciden estos factores en la producción de biotensoactivos, junto con estudios de optimización y mejoramiento de las cepas de microorganismos productores, permitirán el desarrollo tecnológico para la producción económica de los biotensoactivos.
AGRADECIMIENTOS
A la beca PromeP de la Secretaría de Educación Pública (SEP).
REFERENCIAS
Abalos A., Pinazo A., Infante M.R., Casals M.E., García F. y Manresa A. (2001). Physicochemical and antimicrobial properties of new rhamnolipids produced by Pseudomonas aeruginosa AT10 from soybean oil refinery wastes. Langmuir 17, 1367–1371. [ Links ]
Adair D.A.W., Hicks J.R., Jobe D.J. y Reinsborough V.C. (1983). Micellar aggregation numbers of short–chained ionic surfactants through viscosity measurements. Aust. J. Chem. 36, 1021–1025. [ Links ]
Amézcua V.C., Ferrera C.R., Esparza G.F., Ríos L.E. y Rodríguez V.R. (2004). Effect of combined nutrients on biosurfactant produced by Pseudomonas putida. J. Environ. Sci. Health. 39, 2983–2991. [ Links ]
Arima K., Kakinuma A. y Tamura G. (1968). Surfactin, a crystalline peptidelipid surfactant produced by Bacillus subtilis: isolation, characterization and its inhibition of fbrin clot formation. Biochem. Biophys. Res. Commun. 31, 488–494. [ Links ]
Asselineau C. y Asselineau J. (1978). Trehalose–containing glycolipids. Prog. Chem. Fats Other Lipids. 16, 59–99. [ Links ]
Atlas R.M. (1991). Microbial hydrocarbon degradation–bioremediation of oil spills. J. Chem. Technol. Biotechnol. 52, 149–156. [ Links ]
Atlas R.M. (1993). Bacteria and bioremediation of marine oil–spills. Oceanus 36, 71–81. [ Links ]
Banat I.M., Samarah N., Murad M., Horne R. y Benerjee S. (1991). Biosurfactant production and use in oil tank clean–up. World J. Microb. Biot. 7, 80–88. [ Links ]
Banat I.M. (1995). Biosurfactants production and possible uses in microbial enhanced oil recovery and oil pollution remediation: a review. Bioresource Technol. 51, 1–12. [ Links ]
Banat I.M. (2000). Biosurfactants, more in demand than ever. Biofutur. 198, 44–47. [ Links ]
Banat I.M., Makkar R.S. y Cameotra S.S. (2000). Potential commercial applications of microbial surfactants. Appl. Environ. Microb. 53, 495–508. [ Links ]
Berg G., Seech A.G., Lee H., Trevors J.T. (1990). Identification and characterization of a soil bacterium with extracellular emulsifying activity. J. Environ. Sci. Health 25, 753–764. [ Links ]
Bergström S., Theorell H. y Davide H. (1946). On a metabolic product of Ps. pyocyanea, pyolipic acid, active against Myobact. tuberculosis. Ark. Kem. Mineral Geol. 23A, 1–12. [ Links ]
Bodour A. A., Guerrero–Barajas C., Jiorle B.V., Paull A.K., Somogyi A., Trinh L.N., Bates R.B. y Maier R.M. (2004). Structure and characterization of favolipids, a novel class of biosurfactants produced by Flavobacterium sp. Strain MTN11. Appl. Environ. Microb. 70, 114–120. [ Links ]
Brown M.J. (1991). Biosurfactants for cosmetic applications. Int. J. Cosmetic Sci. 13, 61–64. [ Links ]
Cameotra S.S. y Bollag J.M. (2003). Biosurfactantenhanced bioremediation of polycyclic aromatic hydrocarbons. Crit. Rev. Env. Sci. Tec. 33, 111–126. [ Links ]
Cameotra S.S. y Singh H.D. (1990). Purification and characterization of alkane solubilizing factor produced by Pseudomonas PG–1. J . Ferment. Bioeng. 69, 341–344. [ Links ]
Chakrabarty A.M. (1985). Genetically–manipulated microorganisms and their products in the oil service industries. Trends Biotechnol. 3, 32–39. [ Links ]
Chayabutra C., Wu J. y Lu–Kwang J. (2001). Rhamnolipid production by Pseudomonas aeruginosa under denitrification: effects of limiting nutrients and carbon substrates. Biotechnol. Bioeng. 72, 25–33. [ Links ]
Christofi N. y Ivshina I.B. (2002). Microbial surfactants and their use in field studies of soil remediation. J. Appl. Microbiol. 93, 915–929. [ Links ]
Chu W. (2003). Remediation of contaminated soil by surfactantaided soil washing. Practice Periodical of Hazardous, Toxic, and Radioactive Waste Management 7, 19–24. [ Links ]
Cirigliano M. C. y Carman G. M. (1984). Isolation of a bioemulsifier from Candida lipolytica. Appl. Environ. Microb. 48, 747–750. [ Links ]
Costa S.G., Nitschke M., Haddad R., Eberlin M.N. y Contiero J. (2006). Production of Pseudomonas aeruginosa LBI rhamnolipids following growth on Brazilian native oils. Process Biochem. 41, 483–488. [ Links ]
Cooper D.G. y Paddock D.A. (1984). Production of a biosurfactant from Torulopsis bombicola. Appl. Environ. Microb. 47, 173–176. [ Links ]
Cooper D.G., Liss S.N., Longay R. y Zajic J.E. (1989). Surface activities of Mycobacterium and Pseudomonas. J. Ferment. Technol. 59, 97–101. [ Links ]
Cooper D.G., Zajic J.E. y Denis C. (1981). Surface active properties of a biosurfactant from Corynebacterium lepus. J. Am. Oil Chem. Soc. 58, 77–80. [ Links ]
Correa B.F., Colombo F.L. y Ayub M. A. Z. (1999). Production of biosurfactants by hydrocarbon degrading Rhodococcus ruber and Rhodococcus erythropolis. R. Microbiol. 30, 231–236. [ Links ]
Dave H., Ramakrishna C., Bhatt B.D. y Desai J.D. (1994). Biodegradation of slop oil from a petrochemical industry and bioremediation of slop oil contaminated soil. World J. Microb. Biot. 10, 653–656. [ Links ]
Desai J.D. y Banat I.M. (1997). Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 61, 47–64. [ Links ]
Deziel E., Paquette G., Villemur R., Lepine F. y Bisaillon J. (1996). Biosurfactant production by a soil Pseudomonas strain growing on polycyclic aromatic hydrocarbons. Appl. Environ. Microb. 62, 1908–1912. [ Links ]
Dubey K. y Juwarkar A. (2001). Distillery and curd whey waste as viable alternative sources for biosurfactant production. World. J. Microb. Biot. 17, 61–69. [ Links ]
Edwards J.R. y Hayashi J.A. (1965). Structure of a rhamnolipid from Pseudomonas aeruginosa. Arch. Biochem. Biophys. 111, 415–421. [ Links ]
Ferraz C., De Araújo A.A. y Pastore G.M. (2002). The influence of vegetable oils on biosurfactant production by Serratia mercescens. Appl. Biochem. Biotech. 100, 841–848. [ Links ]
Foght J.M., Gutnick D.L. y Westlake D.W.S. (1989). Effect of emulsán on biodegradation of crude oil by pure and mixed bacterial cultures. Appl. Environ. Microb. 55, 36–42. [ Links ]
Francy D.S., Thomas J.M., Raymond R.L. y Ward C.H. (1991). Emulsification of hydrocarbons by subsurface bacteria. J. Ind. Microbiol. 8, 237–245. [ Links ]
Gautam K.K. y Tyagi V.K. (2006). Microbial surfactants: a review. J. Oleo Sci. 55, 155–166. [ Links ]
Georgiou G., Lin S.C. y Sharma M.M. (1992). Surfaceactive compounds from microorganisms. Biotechnol. 10, 60–65. [ Links ]
Ghosh M.M., Yeom I.T., Shi Z., Cox C.D. y Robinson K.G. (1995). Surfactant–enhanced bioremediation of PAH– and PCB–contaminated soils. Microbial Processes for Bioremediation. (R.E. Hinchee, C.M. Vogel y F.J. Brockman, Ed.). Third International Symposium on In Situ and On–Site Bioreclamation, San Diego, abril 24–27, 1995. Batelle Press, Columbus, Ohio. pp 15–23. [ Links ]
Gross B., Montgomery–Brown J., Naumann A. y Reinhard M. (2004). Occurrence and fate of pharmaceuticals and alkylphenol ethoxylate metabolites in an effluent–dominated river and wetland. Environ. Toxicol. Chem. 23, 2074–2083. [ Links ]
Guerra–Santos L., Käppeli O. y Fiechter A. (1984). Pseudomonas aeruginosa biosurfactant production in continuous culture with glucose as carbon source. Appl. Environ. Microb. 48, 301–305. [ Links ]
Haba E., Espuny M.J. Busquets M. y Manresa A. (2000). Screening and production of rhamnolipids by Pseudomonas aeruginosa 47T2 NCIB40044 from waste frying oils. J. Appl. Microbiol. 88, 379–387. [ Links ]
Hartley G.S. (1936). Aqueous solution of the paraffinchains salts: a study in micelle formation. Hermann, París, 69 pp. [ Links ]
Hemminger P. (2005). Bioremediation of contaminated soils. Biocycle 46, 35–38. [ Links ]
Hisatsuka K., Nakahara T., Sano N. y Yamada K. (1971). Formation of rhamnolipid by Pseudomonas aeruginosa and its function in hydrocarbon fermentation. Agr. Biol. Chem. Tokyo 35, 686–692. [ Links ]
Hung H.C. y Shreve G.S. (2001). Effect of the hydrocarbon phase on interfacial and thermodynamic properties of two anionic glycolipid biosurfactants in hydrocarbon/water systems. J. Phys. Chem. 105, 12596–12600. [ Links ]
Itoh S. y Suzuki T. (1972). Effect of rhamnolipids on growth of Pseudomonas aeruginosa mutant deficient in n–parafifin–utilizing ability. Agric. Biol. Chem. Tokyo 36, 2233–2235. [ Links ]
Janneman G.E., McInerney M.J., Knapp R.M., Clark J.B., Feero J.M., Revus D.E. y Menzie D.E. (1983). A halotolerant biosurfactant producing Bacillus species potentially useful for enhanced oil recovery. Dev. Ind. Microbiol. 24, 485–492. [ Links ]
Jarvis F.G. y Johnson M.J. (1949). A glyco–lipide produced by Pseudomonas aeruginosa. J. Am. Chem. Soc. 71, 4124–4126 [ Links ]
Javaheri D., Jenneman G.E., McInerney M.J. y Knapp R.M. (1985). Anaerobic production of a biosurfactant by Bacillus licheniformis JF–2. Appl. Environ. Microb. 50, 698–700. [ Links ]
Karanth N.G.K., Deo P.G. y Veenanadig N.K. (1999). Microbial production of biosurfactants and their importance. Current Science 77, 116–126. [ Links ]
Karanth N.G.K., Gowthaman M.K. y Veenanadig N.K. (2000). Scale up studies for the production of biosurfactant in packed column bioreactor. Bioproc. Biosyst. Eng. 22, 95–99. [ Links ]
Kim E.J. y Shah D.O. (2002). Cloud point phenomenon in amphiphilic drug solutions. Langmuir 18, 10105–10108. [ Links ]
Kim E.J. y Shah D.O. (2003). A cloud point study on the micellar growth of an amphiphilic drug in the presence of alcohol and ionic surfactant. J. Phys. Chem. B. 107, 8689–8693. [ Links ]
Kim H. S., Jeon J.W., Kim B.H., Ahn C.H., Oh C.H. y Yoon B.D. (2006). Extracellular production of a glycolipid biosurfactant, mannosylerythritol lipid, by Candida sp. SY16 using fed–batch fermentation. Appl. Microbiol. Biot. 70, 391–396. [ Links ]
Kissa E. (2001). Fluorinated surfactants and repellents. 2ªed. CRC Press, Boca Ratón, Florida, 640 pp. [ Links ]
Klekner V. y Kosaric N. (1993). Biosurfactants for cosmetics. Biosurfactants: production, properties, applications (V. Klekner, Ed.). Marcel Dekker, Nueva York. pp. 329–372. [ Links ]
Köhler T., Curty L.C., Barja F., Van Delsen C. y Pechère J.C. (2000). Swarming of Pseudomonas aeruginosa is dependent on cell–to–cell signaling and requires fagella and pili. J. Bacteriol. 182, 5990–5996. [ Links ]
Kosaric N. (2001). Biosurfactants and their application for soil bioremediation. Food Technol. Biotechnol. 39, 295–304. [ Links ]
Lin S.C. (1996). Biosurfactants: recent advances. J. Chem. Technol. Biotechnol. 66, 109–120. [ Links ]
Liu Z., Laha S. y Luthy R.G. (1991). Surfactant solubilization of polycyclic aromatic hydrocarbon compounds in soilwater suspensions. Water Sci. Technol. 23, 475–485. [ Links ]
Maier R.M. y Soberón–Chávez G. (2000). Pseudomonas aeruginosa rhamnolipids: biosynthesis and potential applications. Appl. Microbiol. Biot. 54, 625–633. [ Links ]
Makkar R.S. y Cameotra S.S. (1998). Production of biosurfactant at mesophilic and thermophilic conditions by a strain of Bacillus subtilis. J. Ind. Microbiol. Biotechnol. 20, 48–52. [ Links ]
Makkar R.S. y Cameotra S.S. (2002). An update on the use of unconventional substrates for biosurfactant production and their new applications. Appl. Microbiol. Biot. 58, 428–434. [ Links ]
Makula R.A., Lookwood P.J. y Finnerty W.R. (1975). Comparative analysis of the lipids of Acinetobacter species grown on hexadecane. J. Bacteriol. 121, 250–258. [ Links ]
Marahiel M., Denders W., Krause M. y Kleinkauf H. (1977). Biological role of gramicidin S in spore functions. Studies on gramicidin–S–negative mutants of Bacillus brevis ATCC 9999. Eur. J. Biochem. 99, 49–52. [ Links ]
Morikawa M., Daido H., Takao T., Murata S., Shimonishi Y. y Imanaka T. (1993). A new lipopeptide biosurfactant produced by Arthrobacter sp. strain MIS38. J. Bacteriol. 175, 6459–6466. [ Links ]
Mueller J.G., Resnick S.M., Shelton M.E. y Pritchard P.H. (1992). Effect of inoculation on the biodegradation of weathered Prudhoe Bay crude oil. J. Ind. Microb. 10, 95–102. [ Links ]
Mukherjee S., Das P. y Sen R. (2006). Towards commercial production of microbial surfactants. Trends Biotechnol. 24, 509–515 [ Links ]
Nitschke M., Costa S.G.V. y Conteiro J. (2005a). Rhamnolipid surfactants: an update on the general aspects of these remarkable biomolecules. Biotechnol. Prog. 21, 1593–1600. [ Links ]
Nitschke M., Costa S.G.V., Haddad R., Gonçalves L.A.G., Eberlin M.N. y Conteiro J. (2005b). Oil wastes as unconventional substrates for rhamnolipid biosur–factant production by Pseudomonas aeruginosa LBI. Biotechnol. Prog. 21, 1562–1566. [ Links ]
Oberbremer A. y Müller–Hurtig R. (1989). Aerobic stepwise hydrocarbon degradation and formation of biosurfactants by an original soil population in a stirred reactor. Appl. Microbiol. Biot. 31, 582–586. [ Links ]
Oberbremer A., Müller–Hurtig R. y Wagner F. (1990). Effect of the addition of microbial surfactants on hydrocarbon degradation in a soil population in a stirred reactor. Appl. Microbiol. Biot. 32, 485–489. [ Links ]
Ochsner U.A., Hembach T. y Fiecher A. (1996). Production of rhamnolipid biosurfactants. Adv. Biochem. Eng. Biotechnol. 53, 89–118. [ Links ]
Patel M.N. y Gopinathan K.P. (1986). Lysozyme–sensitive bioemulsifier for immiscible organophosphorus pesticides. J. Appl. Microbiol. 52, 1224–1226. [ Links ]
Pellerin N.B., Staley J.T., Ren T., Graff G.L., Treadwell D.R. y Aksay I.A. (1991). Acidic biopolymers as dispersants for ceramic processing. Mater. Res. Soc. Sym. Proc. 218, 123–128. [ Links ]
Perfumo A., Banat I.M., Canganella F. y Marchant R. (2006). Rhamnolipid production by a novel thermophilic hydrocarbon–degrading Pseudomonas aeruginosa AP02–1. Appl. Microbiol. Biot. 72, 132–138. [ Links ]
Peypoux F., Bonmatin J.M. y Wallach J. (1999). Recent trends in the biochemistry of surfactin. Appl. Microbiol. Biot. 51, 553–563. [ Links ]
Pruthi V. y Cameotra S.S. (2003). Effect of nutrients on optimal production of biosurfactants by Pseudomonas putida—A gujarat oil field isolate. J. Surfactants Deterg. 6, 65–68. [ Links ]
Rashedi H., Mazaheri A.M., Bonakdarpour B. y Jamshidi E. (2005). Environmental importance of rhamnolipid production from molasses as a carbon source. Int. J. Environ. Sci. Tech. 2, 59–62. [ Links ]
Rashedi H., Jamshidi E., Mazaheri A. M. y Bonakdarpour B. (2006). Biosurfactant production with glucose as a carbon source. Chem. Biochem. Eng. Q. 20, 99–106. [ Links ]
Robert M., Mercadé M.E., Bosch M.P., Parra J.L., Espuny M.J., Manresa M.A. y Guinea J. (1989). Effect of the carbon source on biosurfactant production by Pseudomonas aeruginosa 44T1. Biotechnol. Lett. 11, 871–874. [ Links ]
Ron E.Z. y Rosenberg E. (2001). Minireview: natural roles of biosurfactants. Environ. Microbiol. 3, 229–236. [ Links ]
Rosenberg E., Zuckerberg A., Rubinovitz C. y Gutnick D. L. (1979). Emulsifer of Arthrobacter RAG–1: isolation and emulsifying properties. Appl. Environ. Microb. 37, 402–408. [ Links ]
Rosenberg E., Schwartz Z., Tenenbaum A., Rubinovitz C., Legmann R. y Ron E. Z. (1989). A microbial polymer that changes the surface properties of limestone: Effect of biodispersan in grinding limestone and making paper. J. Disper. Sci. Technol. 10, 241/250. [ Links ]
Santa–Anna L.M., Sebastian G.V., Menezes E.P., Alves T.L.M., Pereira Jr.N. y Freire D.M.G. (2002). Production of biosurfactants from Pseudomonas aeruginosa PA1 isolated in oil environments. Braz. J. Chem. Eng. 19, 159–166. [ Links ]
Shukla D. y Tyagi V.K. (2006a). Anionic gemini surfactants: a distinct class of surfactants. J. Oleo Sci. 55, 215–226. [ Links ]
Shukla D. y Tyagi V.K. (2006b). Cationic gemini surfactants: a review. J. Oleo Sci. 55, 381–390. [ Links ]
Singh A., Van Hamme J.D. y Ward O.P. (2006). Surfactants in microbiology and biotechnology: Part 2. Application aspects. Biotechnol. Adv. 25, 99/121. [ Links ]
Soberón–Chávez G., Aguirre–Ramírez M. y Sánchez R. (2005). The Pseudomonas aeruginosa RhIA enzyme is involved in rhamnolipid and polyhydroxyalkanoate production. J. Ind. Microbiol. Biot. 32, 675–677. [ Links ]
Stoyanov S.D., Rehage H. y Paunov V.N. (2003). Novel surface tension isotherm for surfactants based on local density functional theory. Phys. Rev. Lett. 91, 086102.1–086102.4 [ Links ]
Syldatk C., Lang S., Matulovic U. y Wagner F. (1985). Production of four interfacial active rhamnolipids from n–alkenes or glycerol by resting cell of Pseudomonas species DSM 2874. Z. Naturforsch. C. 40, 61–67. [ Links ]
Tang X., Zhu Y. y Men Q. (2007). Enhanced crude oil biodegradability of Pseudomonas aeruginosa ZJU after preservation in crude oil–containing medium. World J. Microb. Biot. 23, 7–14. [ Links ]
Tarek A.A.M., Gaber M.A. y Shereen M.S.A. (2006). Optimization of cultural conditions for biosurfactant production from Nocardia amarae. J. Appl. Sci. Res. 2, 844–850. [ Links ]
Thaniyavarn J., Roongsawang N., Kameyama T., Haruki M., Imanaka T., Morikawa M. y Kanaya S. (2003). Production and characterization of biosurfactants from Bacillus licheniformis F2.2. Biosci. Biotech. Biochem. 67, 1239–1244. [ Links ]
Thimon L., Peypoux F., Wallach J. y Michel M.G. (1995). Effect of the lipopeptide antibiotic, iturin A, on morphology and membrane ultrastructure of yeast cells. FEMS Microbiol. Letts. 128, 101–106. [ Links ]
Timar–Balazsy A. e Eastop D. (1998). Chemical principles of textile conservation. Butterworth–Heinemann, London, 480 p. [ Links ]
Uchida Y., Misava S., Nakahara T. y Tabuchi, T. (1989). Factors affecting the production of succinoltrehalose lipids by Rhodococcus erythropolis SD–74 grown on n–alkanes. Agric. Biol. Chem. 53, 765–769. [ Links ]
Van Hamme J.D., Singh A. y Ward O.P. (2003). Recent advances in petroleum microbiology. Microbiol. Mol. Biol. R. 67, 503–549. [ Links ]
Van Os N.M., Haak J.R. y Rupert L.A.M. (1993). Physicochemical properties of selected anionic, cationic and nonionic surfactants. Elsevier, Amsterdam, 608 p. [ Links ]
Vater J. (1986). Lipopeptides, an attractive class of microbial surfactants. Prog. Colloid Polym. Sci. 72, 12–18. [ Links ]
Venkata R.K. y Karanth N.G. (1989). Factors affecting biosurfactant production using Pseudomonas aeruginosa CFTR–6 under submerged conditions. J. Chem. Technol. Biot. 45, 249–257. [ Links ]
Vidali M. (2001). Bioremediation: an overview. Pure Appl. Chem. 73, 1163–1172. [ Links ]
Wouther H. N. y Dick B. J. (2002). Rhamolipids stimulates uptake of hydrophobic compounds by Pseudomonas aeruginosa. Appl. Environ. Microb. 68, 4502–4508. [ Links ]
Yakimov M.M., Timmis K.N., Wray V. y Fredrickson H.L. (1995). Characterization of a new lipopeptide surfactant produced by thermotolerant and halotolerant subsurface Bacillus licheniformis BAS50. Appl. Environ. Microb. 61, 1706–1713. [ Links ]
Yamane T. (1987). Enzyme technology for the lipids industry: an engineering overview. J. Am. Oil Chem. Soc. 64, 1657–1662. [ Links ]
Yateem A., Balba M.T., Al–Shayji Y. y Al–Awadhi. (2002) Isolation and characterization of biosurfactant–producing bacteria from oil–contaminated soil. Soil Sediment. Contam. 11, 41–55. [ Links ]
Zafriri D., Rosenberg E. y Mirelman D. (1981). Mode of action of Myxococcus xanthus antibiotic TA. Antimicrob. Agents Chemother. 19, 349–351. [ Links ]
Zeng G., Zhong H., Huang G. y Fu H. (2005). Physicochemical and microbiological effects of biosurfactant on the remediation of HOC–contaminated soil. Prog. Nat. Sci. 15, 577–585. [ Links ]
Zhang Y. y Miller R.M. (1992). Enhanced octadecane dispersion and biodegradation by Pseudomonas rhamnolipid surfactant (biosurfactant). Appl. Environ. Microb. 58, 3276–3282. [ Links ]

