











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La fragmentación se refiere a la pérdida de continuidad de la vegetación natural o ecosistemas, dejando remanentes de menor tamaño (parches o fragmentos) lo cual afecta tanto la estructura como la diversidad del paisaje (Lord y Norton, 1990; McGarigal y Marks, 1994; Gilbert-Norton et al., 2009). Los procesos más generales que propician la fragmentación de un paisaje son la actividad ganadera y la agrícola, la urbanización y la creación de infraestructura (Lord y Norton, 1990).
Con la finalidad de atenuar los efectos ecológicos negativos de la fragmentación es necesario mantener la conectividad del paisaje conservando las relaciones espaciales entre los elementos que lo conforman (Forman, 1995). La conectividad estructural se presenta cuando dos o más fragmentos se mantienen conectados por una franja de clase de hábitat similar y su ausencia incrementa la fragmentación del paisaje (Rabinowitz y Zeller, 2010). La conectividad funcional evalúa el grado de permeabilidad que presenta un paisaje para ser atravesado por organismos de una misma especie (Pither y Taylor, 1998), es decir, el grado en el cual el paisaje facilita o impide movimientos en ambientes con recursos diferentes a los fragmentos apropiados (Belisle, 2005).
En el sureste de México y Centroamérica la iniciativa del Corredor Biológico Mesoamericano (CBM) tiene como objetivo conservar la diversidad biológica a través de la conectividad de sus áreas naturales protegidas (ANP), además del fomento del desarrollo sostenible de las comunidades rurales (CONABIO, 2009). La Península de Yucatán forma parte de este proyecto y en su parte central se encuentran tres áreas naturales protegidas relevantes: la Reserva Biocultural del Puuc (RBP), el Área de Protección de Flora y Fauna Bala’an K’aax (APB), Quintana Roo y la Reserva de la Biosfera Calakmul (RBC), Campeche (Figura 1; Crooks, 2002; CONABIO, 2009). En esta región se ubica el paisaje Puuc-Chenes, el cual contiene importantes fragmentos de vegetación que en el pasado formaron un continuo entre las selvas caducifolias y subcaducifolias del norte de la Península de Yucatán con las selvas subperennifolias y perennifolias del sur de la Península y Centroamérica (Gobierno del estado de Campeche, 2007a; Bioasesores A. C., 2011).
Figura 1 Localización de la Región Puuc-Chenes y tres áreas naturales protegidas: la Reserva Biocultural del Puuc (RBP), Yucatán, el Área de Protección de Flora y Fauna de Bala’an K’aax (APB), Quintana Roo; la Reserva de la Biósfera de Calakmul (RBC), Campeche. Se muestran las clases de cobertura diferenciadas y fragmentos de selva prioritarios.
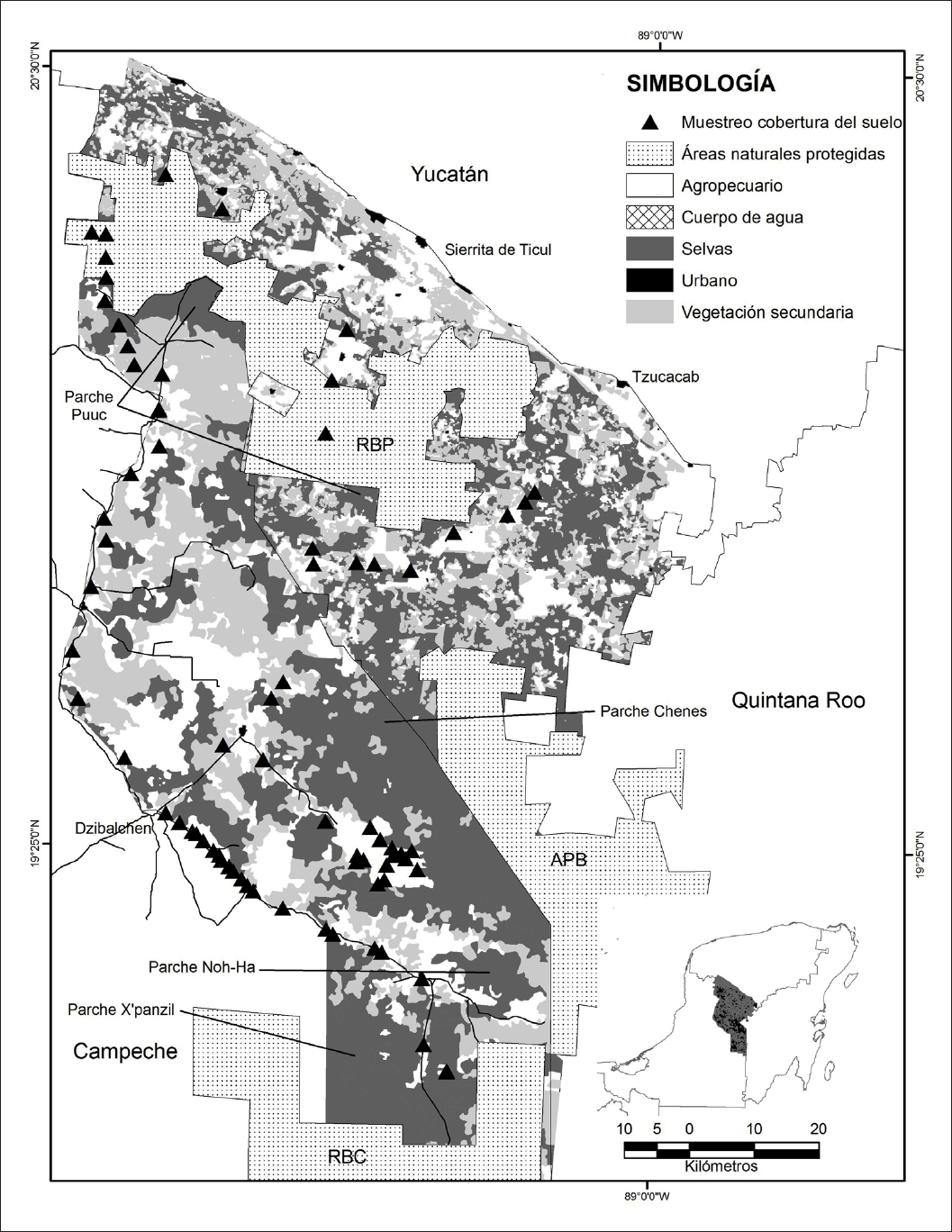
Figure 1 Location of the study region Puuc-Chenes and three protected natural areas: the Biocultural Puuc Reserve (RBP), the Bala’an K’aax Flora and Fauna Protected Area (APB), Quintana Roo, and the Calakmul Biosphere Reserve (RBC), Campeche. The cover classes and priority forest fragments are shown.
Cada población de organismos responde de manera particular a la conectividad de un mismo paisaje, en virtud de que percibe el territorio a diferentes escalas y de acuerdo con requerimientos únicos de hábitat, alimentación y dispersión, entre otros (McGarigal y Marks, 1994). El jaguar (Panthera onca) es un depredador tope de la cadena alimenticia con amplia capacidad de desplazamiento a través de diversos ambientes naturales y modificados por el ser humano, por lo que son organismos clave para estudiar la conectividad (Lasch y Serrano, 2005; Chávez y Zarza, 2009; Colchero et al., 2011). Se ha reportado que los patrones de movilidad y ámbitos hogareños del jaguar son diferentes en función del sexo. Las hembras generalmente se comportan como especie de interior de bosque, mientras que los machos presentan mayor plasticidad que incluye el comportamiento de especie de borde y de interior (Conde et al., 2010); en consecuencia, se asume que la fragmentación del paisaje ejerce mayores restricciones en la conectividad del hábitat para hembras de P. onca y en su capacidad de dispersión en comparación con el P. onca macho. Estudios realizados en la Reserva de la Biosfera Calakmul (RBC) refieren que el ámbito hogareño mínimo para P. onca macho es de 60 a 65 km2, cuyo territorio se sobrepone al de dos hembras en promedio, con una movilidad anual de hasta1 000 km2, siendo la densidad estimada para P. onca de un individuo por cada 15 a 30 km2 (Ceballos et al., 2001; Zarza et al., 2007; Rodríguez-Soto et al., 2011). En el presente estudio se analiza la conectividad estructural y funcional de la región Puuc-Chenes con base en estos requerimientos de hábitat de P. onca en función de su sexo (Cuadro 1). Se considera paisaje a toda el área de estudio evaluada y conformada por diferentes tipos de cobertura natural y antrópica, aquí denominada como región Puuc-Chenes; una clase se refiere a las grandes categorías de cobertura que se diferenciaron en el paisaje (selva, vegetación secundaria, agropecuaria, urbana y cuerpos de agua) y un fragmento se refiere a una porción de alguna clase de cobertura que no mantiene conexión con otra porción del mismo tipo.
Cuadro 1 Requerimientos de hábitat para Panthera onca, con distinción del sexo. Estas variables fueron elegidas para modelar los mapas de hábitat de P. onca: Uso del suelo [selva, cuerpo de agua (Agua), vegetación secundaria (Veg Sec), agropecuario (Agro) y urbano (Urb)]; cercanía a poblados; cercanía a vías de comunicación; y tamaño de los fragmentos de selva. 1 = mínima preferencia; 5 = máxima preferencia (elaboración propia).

MATERIALES Y MÉTODOS
Área de estudio.
La región Puuc comprende parte del suroeste de Yucatán y el noreste de Campeche (Bioasesores A. C., 2011) y la región de Los Chenes, colinda en su extremo norte con el Puuc yucateco y abarca parte de los municipios de Hopelchén y Calakmul en Campeche (Porter-Bolland et al., 2008); en conjunto conforman la región Puuc-Chenes con una superficie de 972 578 ha. Al norte la limita la sierrita de Ticul en Yucatán y la carretera que corre paralela a la Sierrita, e incluye a la Reserva Biocultural del Puuc (RBP); al este está limitada por el borde norte y oeste del Área de Protección de Flora y Fauna Bala’an K’aax (APB), Quintana Roo, al sur por el borde norte de la Reserva de la Biósfera Calakmul (RBC), Campeche, y al oeste por la carretera que parte de Bolonchén de Rejón a Dzibalchén en el municipio de Hopelchén, Campeche (Figura 1).
En la región Puuc y las zonas norte-centro de Los Chenes la selva mediana subcaducifolia está inmersa en una matriz de vegetación secundaria, áreas de cultivo y potreros; en el sur de Los Chenes predomina la selva mediana subperennifolia (Gobierno del Estado de Campeche, 2007a).
Generación de mapas
Se elaboró un mapa de vegetación y uso de suelo de la zona de estudio con base en Imágenes Landsat 7 ETM+ con pixeles de 30 m, utilizando ArcMap (versión 9.3, ESRI Inc, New York Street, CA, USA). Para ello se contó con los siguientes mapas:
Región Puuc: mapa digital proveniente del Estudio Regional Forestal (Bioasesores A. C., 2011), que incluye los municipios de Akil, Dzan, Maní, Muna, Oxkutzcab, Sascalúm, Santa Elena, Tekax, Ticul y Tzucacab, ubicados en el sur y suroeste del estado de Yucatán. Este mapa fue elaborado con imágenes de satélite Landsat 7 ETM+ correspondientes a 2005 y 2008.
Los Chenes: mapa digital proveniente del Ordenamiento Ecológico y Territorial del municipio de Hopelchén, de 2007, elaborado con imágenes satelitales Landsat 7 ETM+ del 2003 (Gobierno del Estado de Campeche, 2007a).
Ambos mapas fueron actualizados a 2012, apoyados en recorridos de campo, lo que permitió corregir y recategorizar las etiquetas de los tipos de vegetación y uso del suelo. Se homogeneizaron los mapas ajustando la clasificación a las clases propuestas por INEGI serie IV (INEGI, 2012). La validación en campo tuvo lugar en los meses de estiaje (mayo-junio) dedicando 11 salidas a lo largo de los principales caminos y brechas de la región Puuc-Chenes.
En total se registraron 75 sitios (30 de vegetación secundaria, 30 de agropecuario, siete de selva mediana subcaducifolia, cuatro de selva mediana subperennifolia, tres de selva baja subcaducifolia y uno de selva baja subperennifolia). Del total de sitios muestreados, a 16 se les aplicaron cambios en la clasificación de su cobertura del suelo (21.33% del total) como sigue: 13 sitios referidos como selva se recategorizaron en la clase de agropecuario y tres sitios referidos en el mapa como selva mediana subcaducifolia correspondieron a selva baja subcaducifolia; estos cambios se integraron al mapa y se corrigieron las tablas de atributos. Posteriormente se llevó a cabo la rasterización de los mapas con un tamaño de celda de 50 x 50 m y se llevó a cabo su limpieza topológica en ambiente de ArcMap (versión 9.3). Finalmente, y con base en los requerimientos de hábitat de P. onca (Cuadro 1; Zarza et al., 2007, Chávez y Zarza, 2009; Conde et al., 2010), se llevó a cabo una reclasificación de las coberturas del suelo generando la categoría “selva” que incluyó a los cuatro tipos identificados en los mapas (selva mediana subperennifolia, selva mediana subcaducifolia, selva baja subperennifolia y selva baja subcaducifolia). Se consideró como selva a la vegetación con más de 30 años de edad, de acuerdo con la nomenclatura maya para las etapas sucesionales de la vegetación descrita para la Península de Yucatán por Flores y Ucán (1983) y Sánchez et al. (2007). La vegetación secundaria fue catalogada como una sola clase, sin considerar su edad sucesional; los pastizales y áreas agrícolas se agruparon en la tercera clase nominada agropecuaria; la clase urbano incluye poblados, caminos e infraestructura antrópica y, por último, se diferenció la clase cuerpos de agua (Figura 1). Cabe señalar que el uso de imágenes de satélite Landsat ETM+ para la clasificación de vegetación y uso de suelo es el más generalizado en estudios regionales (por ej., Vega-Guzmán et al., 2008; Redo, 2012; Zhao, 2013) y con base en estos mapas se han realizado estudios previos que analizan el funcionamiento del jaguar en el paisaje (Colchero et al., 2011; Rodríguez-Soto et al., 2011; Zeilhofer et al., 2014).
Métricas para estudiar la fragmentación y conectividad estructural del paisaje
Se aplicó el programa FRAGSTATS (versión 4.0, McGarigal, 2015) para cuantificar la estructura del territorio usando las cinco clases de cobertura del suelo generadas: 1) selvas, 2) vegetación secundaria, 3) agropecuario, 4) urbano y 5) cuerpos de agua. Se consideró a las selvas y los cuerpos de agua como parte del paisaje natural del felino; la vegetación secundaria, al tratarse de un mosaico de coberturas en diferentes estadios de sucesión, fue considerada una cobertura de transición entre la cobertura natural; los espacios urbanizados y las áreas agropecuarias se consideraron como desfavorables para los felinos (Zarza et al., 2007, Chávez y Zarza, 2009; Conde et al., 2010). Con base en estos criterios se elaboró una matriz de contraste en la cual se asignaron valores en función del grado de contraste que percibe el felino al atravesar las diferentes clases de cobertura de suelo que conforman toda la estructura del paisaje. Para ello se partió de la noción que no todas las fronteras son iguales, por ello se asignó un valor (entre cero y uno) al que llamamos valor de contraste, que describió el grado de disimilitud entre pares de clases de usos de suelo a lo largo de su perímetro (Cuadro 2; Valls Dalmau, 2011).

Métricas del paisaje
De acuerdo con las recomendaciones encontradas en la literatura (McGarigal y Marks, 1994; Grainger et al., 2005; McGarigal, 2015), se eligieron medidas absolutas en el caso del paisaje, y medidas absolutas y relativas al comparar las clases en virtud de que tienen superficies y números de fragmentos diferentes. Con base en la significancia ecológica para evaluar la conectividad estructural desde la perspectiva de los requerimientos de hábitat de P. onca, se calcularon los índices de área y borde, de contraste, de aislamiento y de agregación. A nivel paisaje, las métricas de contraste se enfocan en los bordes como límites y potenciales impedimentos al movimiento de los organismos y hacen referencia a cómo los bordes influyen en la conectividad del paisaje; las métricas de aislamiento, describen el grado de separación espacial entre los fragmentos, haciendo referencia al grado de conectividad del paisaje así como su impacto en la dispersión de los organismos (McGarigal y Marks, 1994).
Descripción de los requerimientos de hábitat de Panthera onca en función de su sexo
La variable cobertura del suelo incluyó: selva, vegetación secundaria, área agropecuaria, urbana y cuerpos de agua. La literatura refiere que ambos —machos y hembras— prefieren cubierta de bosque denso (selvas), aunque los machos de P. onca transitan por vegetación secundaria e incluso de áreas agropecuarias con mayor frecuencia que las hembras, y ambos sexos evitan zonas urbanas (Colchero et al., 2006; Zarza et al., 2007; Conde et al., 2010). La segunda variable, cercanía a poblados, se separó en tres categorías: <2 km, 2 a 6.5 km y >6.5 km. La literatura refiere que P. onca evita los poblados hasta los 6 km de distancia; sin embargo, P. onca macho hace incursiones a los poblados con mayor frecuencia que la hembra (Colchero et al., 2006; Zarza et al., 2007; Conde et al., 2010). La tercera variable cercanía a las vías de comunicación (pavimentadas y no pavimentadas), en virtud de que se considera el efecto que ejercen estos caminos favoreciendo la cacería sobre P. onca (Hernández y Calmé, 2002), se diferenciaron las categorías: <1 km, 1 a 4 km y >4 km. Se reporta que P. onca evita las carreteras hasta los 4 km de distancia; sin embargo, el macho se acerca e incluso atraviesa las vías de comunicación (Colchero et al., 2006; Zarza et al., 2007; Conde et al., 2010) y la cuarta variable, tamaño de los fragmentos, se separó en dos categorías: <60 km2 y >60 km2; estas se determinaron de acuerdo al tamaño del ámbito hogareño de P. onca macho, el cual es de 60 km2, y que corresponde al territorio de dos hembras en promedio (Cuadro 1; Ceballos et al., 2001; Zarza et al., 2007; Rodríguez-Soto et al., 2011).
Análisis de la conectividad funcional para Panthera onca
De acuerdo con los requerimientos de P. onca (con análisis de cada sexo por separado), se elaboró un modelo de conectividad funcional identificando las rutas de menor costo que permitieran su movilidad en el paisaje con base en su resistencia al desplazamiento. Se utilizó el panel “Habitat Suitability” del módulo “Land Change Modeler” del programa IDRISI Taiga (Eastman, 2009). Para generar los mapas de hábitat para cada sexo se utilizó una resolución de pixeles de 50 × 50 m, ya que es el mínimo requerido por IDRISI para correr el módulo mencionado (Pascual-Hortal, 2006; Rabinowitz y Zeller, 2010; Rayfield et al., 2010). Las variables elegidas para modelar los mapas de hábitat se basaron en los requerimientos de hábitat que presenta cada sexo (Lasch y Serrano, 2005; Colchero et al., 2006; Zarza et al., 2007; Chávez y Zarza, 2009; Conde et al., 2010; Colchero et al., 2011; González, 2011). Las variables descriptivas del modelo fueron la cobertura del suelo, la cercanía a poblados, la cercanía a las vías de comunicación y el tamaño de los fragmentos; la cobertura de hidrología no se utilizó en virtud de que la zona carece de cuerpos de agua permanentes. Con base en estas variables se crearon categorías con sus respectivas zonas de influencia/amortiguación; a las variables cercanía a poblados y vías de comunicación se les asignó un valor numérico de acuerdo al sexo (Cuadro 1).
Modelación del hábitat
Se utilizó el módulo “Land Change Modeler” para modelar el hábitat de P. onca, a través de la creación de un modelo empírico, ya que en el presente estudio se contó con 19 registros de presencia de P. onca en la región, los cuales se obtuvieron de observaciones personales, mediante reportes técnicos e imágenes captadas con cámara trampa (Observaciones del Fondo de aseguramiento de la Confederación Nacional de Organizaciones Ganaderas, 2011 y de Pronatura Península de Yucatán A.C., 2012). Se aplicó el modelo estadístico “Mahalanobis typicality” (Rodríguez-Soto et al., 2011) del programa IDRISI Taiga (Eastman, 2009). Este algoritmo marcó una celda con valor de 1.0 cuando determinó que, para ese punto, las variables analizadas en la construcción del modelo (medioambientales y antrópicas) eran idénticas a la media de las variables encontradas en los puntos de entrenamiento del paisaje (datos de presencia de P. onca); los valores se aproximaron a 0 cuando para determinada celda las variables analizadas fueron diferentes a las encontradas en los puntos de entrenamiento. Cualquier celda con valor superior a cero presentó alguna similitud con los puntos tomados como datos de entrenamiento.
Definición de corredores
No hay evidencia en la literatura que describa el ancho de los corredores para P. onca; sin embargo, hay reportes para puma (Puma concolor) los cuales oscilan entre 90 y 500 m (Beier, 1995; Maehr et al., 2002; González, 2011). Con esta base se optó por asignar un ancho promedio de 200 m para el ancho del corredor de P. onca.
Posteriormente, con base en los mapas de hábitat, se identificaron con el panel “Corridor Planning” del módulo “Land Change Modeler” (IDRISI Taiga, Eastman, 2009), los corredores biológicos —con un ancho de 200 m— entre los tres fragmentos más importantes del paisaje (Puuc, Chenes y X´Panzil). La validación en campo del estado actual de los corredores se llevó a cabo en puntos cercanos a las vías de comunicación.
Identificación de áreas prioritarias para su conservación en el paisaje Puuc-Chenes
Se consideraron como áreas estratégicas para la conservación los fragmentos que cumplieron con las siguientes características: a) fragmentos de selva con áreas mayores a los 900 km2, en virtud de que esta es la extensión mínima necesaria para conservar una población viable de P. onca (Ceballos et al., 2001; Rodríguez-Soto, 2011) y b) fragmentos de selva prioritarios para mantener la conectividad para P. onca a lo largo de los corredores biológicos identificados. Se utilizó el programa ArcMap (versión 9.3. ESRI Inc, New York Street, CA, USA) para seleccionar las áreas de importancia para la conservación.
RESULTADOS
Conectividad estructural del paisaje Puuc-Chenes con base en los requerimientos de hábitat de Panthera onca
Índices de área y número de fragmentos. La extensión total del paisaje Puuc-Chenes es de 972 578 ha. La clase de paisaje con mayor superficie fue la selva, que ocupa 49.8% del área. El fragmento más grande del Paisaje Puuc-Chenes también es de selva, que abarca 19.7% del paisaje y cuya extensión es de 191 600 ha (LPI). El paisaje total presentó 2 509 fragmentos, de los cuales 935 son de origen antrópicos (agropecuario con 904 y urbano con 31), 1 254 corresponden a vegetación secundaria, 313 a selvas y siete a cuerpos de agua (Cuadro 3).
Cuadro 3 Resultados de las métricas aplicadas a nivel paisaje y de clases desde la perspectiva de Panthera onca. Clases de uso del suelo: selva, vegetación secundaria, agropecuario, urbano y cuerpos de agua.

Índices de borde y de contraste. El índice de contraste total de los bordes (TECI) para el paisaje es de 43.7%. El valor más alto del contraste ponderado de la densidad del borde (CWED) se encuentra en la clase de selva (2.5 m ha−1) y el valor más bajo de este índice lo presentan los cuerpos de agua (0.0). La clase con el mayor índice de densidad de borde (ED) es la vegetación secundaria con 5.6 m ha−1, seguido por la selvas con 4.8 m ha−1.
Índices de aislamiento. Los fragmentos de selva presentan el índice de proximidad ponderado por área (PROX_AM) más alto (8,701) y el valor del índice más bajo se encontró en los cuerpos de agua (0). La distancia euclidiana más cercana (ENN_AM) corresponde a la selva (221 m) lo cual nos indica que los fragmentos de selva presenta la menor distancia entre ellos. Los cuerpos de agua presentan la distancia más lejana (11 049 m) entre ellos. En cuanto al índice de contagio (CONTAG) para el total del paisaje es de 62.5%.
Identificación de áreas prioritarias para su conservación en el paisaje Puuc-Chenes
El paisaje total de la región Puuc-Chenes ocupa 9 725.8 km2. En este se identificaron dos fragmentos de selva en el paisaje Puuc-Chenes con área superior a los 900 km2 para ser considerada como mínima necesaria para conservar una población viable de P. onca (Ceballos et al., 2001; Rodríguez-Soto, 2011): el fragmento del Puuc con 1 916 km2 y el fragmento de Los Chenes con 1 380 km2. Además, se registraron dos fragmentos de selva con área menor a los 900 km2 pero prioritarios para mantener la conectividad de P. onca a lo largo de los corredores biológicos: el fragmento X´Panzil con 679 km2 y el fragmento Noh-Ha con 88.5 km2, lo que da un total del paisaje identificado para la conservación de jaguar de 4 063 km2 (Figura 2).
Figura 2 Acercamiento a la región Puuc-Chenes donde se localizan los cuatro corredores identificados para mantener la conectividad de P. onca entre los cuatro fragmentos de selva prioritarios (Puuc, Chenes, Noh-Ha y X´Panzil) y las tres Áreas Naturales Protegidas del paisaje Puuc-Chenes: la Reserva Biocultural del Puuc (RBP), Yucatán, el Área de protección de flora y fauna de Bala’an K’aax (APB), Quintana Roo; la Reserva de la Biósfera de Calakmul (RBC), Campeche.

Figure 2 Outlining the biological corridors for the P. onca functional connectivity in the Puuc-Chenes region, linking the four priority forest fragments (Puuc, Chenes, Noh-Ha and X´Panzil) and the three natural protected areas of the Puuc-Chenes landscape: The Biocultural Puuc Reserve (RBP), the Bala’an K’aax flora and fauna protected area (APB), Quintana Roo, and the Calakmul Biosphere Reserve (RBC), Campeche.
Conectividad funcional del paisaje Puuc-Chenes con base en los requerimientos de hábitat de Panthera onca
En el paisaje Puuc-Chenes se ubicaron cuatro corredores biológicos para P. onca de al menos 200 m de ancho; de estos, los dos primeros, corredor 1 y 2, enlazan los fragmentos Puuc y Chenes (corredores Puuc-Chenes) y los corredores 3 y 4 enlazan los fragmentos Chenes y X´Panzil (corredores X´Panzil-Chenes; Figura 2). El corredor 1 con conectividad para ambos sexos se ubicó entre los poblados de Poltok, Tekax y San Sebastián, Tzucacab y Nacaveba en Yucatán.; este corredor no presentan carreteras y solo es atravesado por terracerías; siendo sus principales coberturas de suelo la vegetación secundaria y la agropecuaria. El corredor 2, con conectividad para P. onca macho, se ubicó entre los poblados de San Felipe Segundo y Tres Hermanitas, ambos en municipio de Tekax, en Yucatán; está compuesto por un mosaico de selvas, vegetación secundaria y cultivos y es atravesado por la carretera pavimentada dos carriles que une a los poblados de Hunto-Chac y Becanchén. Los corredores 3 y 4 (corredores X´Panzil-Chenes) se caracterizan por conectar fragmentos de selva en buen estado de conservación y con pocos poblados; los dos corredores presentaron conectividad funcional para ambos sexos. La ruta del corredor 3 se conecta a través del fragmento Noh-Ha, se ubica entre los poblados de Chan Chen y Chun’Ek, Hopelchén y solo lo atraviesa una terracería. El corredor 4 es atravesado por la carretera estatal que conecta los poblados de Chunchintok y Ukum a nivel de la siguiente coordenada: 19º 17’ 57.52” N y 89º 28’ 20.95” W, Hopelchén.
DISCUSIÓN
La fragmentación y su impacto en Panthera onca
En el paisaje Puuc-Chenes la distribución espacial de sus elementos no es homogénea. Las clases antrópicas (agropecuario y urbano) se ubican cercanas a las principales vías de comunicación (Maxcanú-Tzucacab y Hunto Chac-Sudzal Chico en Yucatán, y Bolonchén de Rejón-Dzibalchén en Campeche) y los fragmentos de selva de mayor extensión y conectividad se localizan de norte a sur a lo largo de la parte oriental del paisaje. Nuestros resultados muestran que el área de estudio aún presenta alto valor para la conservación del hábitat para P. onca, ya que 49.8% son selvas y 27% son de vegetación secundaria, de tal manera que 76.8% de la cobertura total del suelo en el área de estudio corresponde a clases de vegetación utilizadas comúnmente como hábitat por P. onca (Zarza et al., 2007; Chávez y Zarza, 2009; Rabinowitz y Zeller, 2010; Colchero et al., 2011). Los cuatro fragmentos de selva identificados representan en conjunto 41.8% del paisaje Puuc-Chenes. Sin embargo, el fragmento del Puuc que es el de mayor extensión presenta un elevado grado de perforación, principalmente en su área central y sur; es decir, no conforma una masa continua de selva ya que tiene fragmentos de otro tipo de cobertura inmersos en el mismo, lo que afecta la conectividad al interior del fragmento y su calidad como hábitat. Se ha señalado que la extensión mínima de hábitat necesaria para conservar una población viable de P. onca es de 900 km2 (Schaller y Crawshaw, 1980; Ceballos et al., 2001; Scognamillo, 2001; Maffei, 2004; Zarza et al., 2007; Conde et al., 2010; Rodríguez-Soto et al., 2011 ); en el presente estudio encontramos que la extensión de hábitat para P. onca en el paisaje Puuc-Chenes, aunada al área correspondiente a las ANP adyacentes (Reserva de la Biosfera Calakmul, Campeche, el Área de Protección de Flora y Fauna Bala’an K’aax, Quintana Roo y la Reserva Biocultural del Puuc, Yucatán), presenta un total de 15 943 km2, lo cual asegura la persistencia de P. onca en la región. Sin embargo, esta zona presenta fuertes presiones antrópicas resultantes de la expansión de grupos étnicos y de nuevas colonias de menonitas asentadas en el municipio de Hopelchén. Este tipo de productores se dedican a lo agrícola-ganadero y utilizan tecnología mecanizada para trabajar las tierras —las cuales compran a los ejidos y pequeños propietarios de la región— y producen carbón como un producto del desmonte que realizan para aprovechar sus tierras (Gobierno del estado de Campeche, 2007b). En los últimos años las poblaciones aledañas menonitas están cambiando rápidamente la estructura y composición del paisaje, siendo un factor preponderante en el aumento de las tasas de deforestación. Esta presión está dada tanto por el crecimiento demográfico como por el uso extensivo del suelo, con grandes desmontes, cuyo método, además de ser mecanizado, se lleva a cabo utilizando maquinaria pesada, aplicando fertilizantes y pesticidas, los cuales alteran la recuperación del suelo y, por ende, la de la selva (Porter-Bolland et al., 2008), cambio que está conduciendo a una cascada de efectos negativos para el hábitat y las presas del jaguar.
Conectividad estructural del paisaje Puuc-Chenes para Panthera onca
Diversos estudios destacan la relevancia del índice de contraste total de los bordes (TECI). Esta es una métrica relacionada con el grado de conectividad del paisaje ya que resalta la magnitud de contraste entre los fragmentos de las diferentes clases con la finalidad de determinar el potencial de desplazamiento de los organismos a través del paisaje (Mc-Garigal y Marks, 1994; Botequilha y Ahern, 2002). En el presente trabajo, el TECI presentó un valor de 43.7%, el cual indica un contraste medio entre los fragmentos (McGarigal, 2015). Es interesante resaltar que al utilizar una métrica ponderada se amortigua la influencia del tamaño de las clases, ya que este índice estandariza la longitud de los bordes por unidad de área y facilita la comparación entre clases de diferente tamaño (McGarigal y Marks, 1994). Sin embargo, los resultados reportados por el contraste ponderado de la densidad de borde (CWED) para la vegetación secundaria (matriz del paisaje) y la selva deben ser considerados con cautela al tratarse de las clases dominantes del paisaje y, en consecuencia, presentar una mayor probabilidad de que sus bordes entren en contacto con las demás clases estudiadas y con el borde del paisaje.
Con respecto al análisis del aislamiento, la selva presentó el índice de proximidad ponderado por área más alto (PROX_AM de 8 701), lo cual confirma que la selva presenta fragmentos más grandes y más contiguos (menor aislamiento), en comparación con las otras clases del paisaje y, en consecuencia, presentan la mejor conectividad estructural del paisaje en virtud de que favorece la capacidad de dispersión de P. onca. Por su parte, Bender et al. (2003) señala la relevancia de evaluar el aislamiento entre los fragmentos, ya que con esto se evalúa la dispersión de los organismos y apunta que la métrica más recomendada es el índice de distribución de proximidad.
La métrica de agregación evaluada aquí con el índice de contagio (CONTAG 62.5%) indica que existe un paisaje con fragmentos más grandes y más contiguos (McGarigal y Marks, 1994; Badii y Landeros, 2007). Grainger et al. (2005) encontraron que un índice de contagio elevado está relacionado positivamente con un incremento de la conectividad y, por ende, en una mayor área de distribución de elefantes en África.
Con respecto a la cantidad de fragmentos, los máximos valores los obtuvieron la vegetación secundaria y las áreas agropecuarias (1 254 y 904, respectivamente), ocupando en conjunto casi el 50% del paisaje, lo cual es un indicador de fragmentación. Este resultado era de esperarse, ya que se trata de la matriz del paisaje y representa las clases más fragmentadas. Por el contrario, la selva ocupa 49.8% del paisaje con tan solo 313 fragmentos. Sin embargo, para P. onca esta matriz no resulta tan contrastante (Zarza et al., 2007) dado que la vegetación secundaria ocupa 27% del paisaje,. El paisaje Puuc-Chenes presenta una conectividad estructural superior a la media para P. onca (en virtud de que los valores referidos por el índice de contagio y el índice de contraste total de los bordes, son superiores al valor medio de estos índices) y presenta una importante cantidad de hábitat para el mantenimiento de una población viable, situación que favorece la presencia y dispersión de P. onca en el paisaje.
Conectividad funcional del paisaje Puuc-Chenes para Panthera onca
Los grandes carnívoros son organismos que tienen bajas densidades poblacionales, ya que requieren de amplios territorios (De Angelo et al., 2011), situación que los hace especialmente susceptibles a las modificaciones de la cobertura del suelo. Crooks (2002) documentó los efectos negativos (baja persistencia de la especie a largo plazo como consecuencia de su baja densidad, endogamia y la necesidad de amplios territorios para cazar sus presas) que la fragmentación y la pérdida del hábitat ejercen sobre los carnívoros de talla grande, y se reporta que los corredores favorecen el intercambio genético a través de la dispersión de los organismo entre los fragmentos (Rabinowitz y Zeller, 2010). Gilbert-Norton et al. (2009) comprobó que los corredores incrementan en casi un 50% el movimiento de los individuos entre los fragmentos en comparación con aquellos fragmentos sin conexión, información que resalta la relevancia de conservar los corredores identificados para P. onca entre los principales fragmentos del paisaje Puuc-Chenes.
Gavashelishvili y Lukarevskiy (2008), así como Rabinowitz y Zeller (2010), determinaron corredores de menor costo para el leopardo (Panthera pardus) en Asia y para P. onca a lo largo de su área de distribución y González (2011) estableció corredores para el puma (Puma concolor); sin embargo, en estos no se hizo diferencia entre corredores para cada sexo a pesar de que la literatura sugiere su relevancia. Como ya se mencionó, P. onca macho y hembra presentan diferentes requerimientos en el uso del hábitat (Conde et al., 2010).
Es relevante señalar que en el presente estudio se extrapolaron valores promedios de pumas (Puma concolor) de Canadá y Estados Unidos de Norteamérica para determinar el ancho de los corredores para P. onca, de tal manera que se obtuvo una aproximación a los datos biológicos de P. onca.
En esta investigación encontramos que, de los cuatro corredores resultantes, tres fueron viables para machos y hembras (corredores 1, 3, y 4) y solo uno adecuado para el macho de P. onca (corredor 2). No se esperaba que el corredor 1 presentara conectividad para hembras de P. onca, debido a estas tratan de evitar las áreas agropecuarias, prefiriendo las selvas (Conde et al., 2010); sin embargo, también refiere la literatura que hembras de P. onca muestran preferencia por los paisajes sin carreteras y sin poblaciones cercanas (Zarza et al., 2007; Conde et al., 2010), características que presentan el corredor 1. El corredor 2 solo establece conectividad exclusiva para machos de P. onca, que aunque presenta un mejor estado de conservación de la selva remanente, es atravesado por una carretera estatal de dos carriles, las cuales tienden a ser evitadas por las hembras de P. onca (Lasch y Serrano, 2005; Chávez y Zarza, 2009). El corredor 3 conecta áreas mejor conservadas del paisaje Puuc-Chenes, ya que además por el este se continúan con vegetación de selva en Quintana Roo que es parte de la APB. El corredor 4, a pesar de ser atravesado por una carretera federal de cuatro carriles, no presenta poblaciones cercanas y aún conserva su cobertura de selva natural, lo cual hace a ambos corredores accesibles para hembras de P. onca; sin embargo, estos corredores se ven amenazados por el avance de la frontera agrícola.
Al ser P. onca un organismo críptico (Isasi-Catalá y Barreto, 2008) se sugiere realizar más investigaciones con la finalidad de validar los corredores identificados en el presente estudio utilizando métodos de campo que permitan determinar la presencia/ausencia de P. onca en los mismos, como son es el uso de cámaras trampa, registro de huellas y telemetría de los organismos tal como sugieren otros trabajos que estudiaron poblaciones de tigres, linces y jaguares (Kramer-Schadt et al., 2004; Carroll y Miquelle, 2006; Linkie et al., 2006; Michalski et al., 2006). También es importante elaborar mapas de calidad de hábitat con variables no consideradas en el presente estudio tales como: densidad de presas, condiciones ecológicas de los fragmentos y escorrentías del paisaje, entre ellas la presencia de fuentes de aprovisionamiento de agua de menor magnitud (aguadas, sartenejas y rejolladas; Contreras, 2011; Rodríguez-Soto, 2011).

