











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En la actualidad las emisiones de gases de efecto invernadero (GEI) siguen en aumento; el uso desmedido de combustible fósiles ha provocado que al año 2017 se estime un aumento de 1 ºC en la temperatura global (Infante, 2019). Otros registros similares de temperatura sobre la superficie terrestre y oceánica demuestran un calentamiento de 0.87 ºC (Allen et al., 2019). Se estima que las emisiones antrópicas de CO2 a la atmósfera fueron de 3.6 Gt de CO2 en el periodo de 2008 a 2018 (BP PLC, 2019), estas emisiones en su mayoría son almacenadas en plantas, suelo y océanos (IPCC, 2015); por lo anterior, el aumento de superficies boscosas, la conservación, buenas prácticas silvícolas y reforestación son importantes para el almacenamiento del carbono en la superficie terrestre (De Jong et al., 2004), de ahí la importancia de contar con ecuaciones alométricas para cuantificar el carbono almacenado en árboles individuales y por tipos de ecosistema (Graciano-Ávila et al., 2019; Montoya et al., 2018).
Para la estimación del carbono en la biomasa de árboles se han desarrollado métodos directos e indirectos, los primeros consisten en derribar los árboles, separar cada componente para obtener el peso total en verde y seco (Picard et al., 2012) y los segundos estiman la biomasa por medio de ecuaciones obtenidas con análisis de regresión sin derribar el árbol (Avendaño et al., 2009; Návar, 2009; Picard et al., 2012).
En algunas áreas es imposible derribar árboles para determinar de manera directa la biomasa; entonces, es conveniente utilizar el volumen del árbol, obtenido a través de métodos indirectos o por troceo simulado (Salas et al., 2005); posteriormente, el volumen se transforma en biomasa con el uso de la densidad básica correspondiente a la especie (Silva-Arredondo y Návar-Cháidez, 2010) para generar ecuaciones alométricas. En 1855 Maximilian Robert Pressler postuló un método, denominado como ecuación de Pressler, para obtener el volumen del fuste en el que sólo se necesita medir el diámetro normal (a 1.3 m) y la altura a la que se encuentra la mitad del diámetro normal; se aplica a cualquier forma geométrica que presente el fuste, siempre que sea no cilíndrico (Romahn, 1999). Los estudios de Curto et al. (2019), Lima et al. (2016) y Valencia-Manzo et al. (2017) constituyen ejemplo de estimaciones de volumen de fuste en especies arbóreas con la ecuación de Pressler.
Pseudotsuga menziesii (Mirb.) Franco es ampliamente distribuida en el oeste de EE.UU. y Canadá (Hermann y Lavender, 1999); en cambio, las poblaciones en México han sido deterioradas y se encuentran bajo protección especial en la Norma Oficial Mexicana NOM-059-SEMARNAT-2001 (SEMARNAT, 2002), y como consecuencia, su aprovechamiento no está permitido. Crece en forma natural en la porción norte de la Sierra Madre Occidental y en la parte norte de la Sierra Madre Oriental (Ventura et al., 2010), llega a alcanzar hasta 90 m de altura y 309 cm de diámetro (Van Pelt y Nadkarni, 2004), lo que supone que puede almacenar grandes cantidades de carbono en su biomasa, pero también, esto dificulta cuantificar su biomasa de forma directa, además de ser costoso.
La escasez de ecuaciones de biomasa para P. menziesii es evidente, salvo las investigaciones hechas por St. Clair (1993) y Feller (1992) en EE.UU. y otras por Návar (2009) en el noroeste de México, aunque existen otras temáticas referentes a la caracterización y conservación (Ventura et al., 2010), a su estructura silvícola (Guerra et al., 2012) y sobre aspectos ecológicos (Domínguez et al., 2004).
El presente estudio tuvo como objetivo generar una ecuación para estimar biomasa de fuste en Pseudotsuga menziesii (Mirb.) Franco en bosque sin manejo en la Sierra de Arteaga Coahuila, México, mediante el método de Pressler, que sirva de base para cuantificar los almacenes de carbono en este tipo de poblaciones.
Materiales y métodos
Área de estudio
El trabajo se realizó en un bosque de P. menziesii ubicado a 13 km al este del poblado San Antonio de las Alazanas en Arteaga, Coahuila, México, en las coordenadas 25º 13’ 59.25” N y 100º 26’ 55.97” O (Datum WGS84), con altitud de 2684 m; presenta clima de tipo Cw1 (templado subhúmedo) y Cb´(w1)x´ (templado semifrío subhúmedo), con temperatura media que oscila entre los 5 y 18 ºC y precipitación anual de 400 a 600 mm (García, 1998).
Muestreo de arbolado
Para la generación de la ecuación de biomasa de fuste (Bf) se usó el método indirecto; se aplicó un muestreo selectivo de árboles de P. menziesii, se consideraron todas las categorías diamétricas existentes, se seleccionaron árboles con fuste recto, sin daños, libres de plagas y enfermedades (Avendaño et al., 2009). Para medir el diámetro normal a 1.30 m (Dn), la altura del punto directriz (Hp), donde se ubica la mitad del Dn y la altura total (Ht) se utilizó un dendrómetro electrónico (Criterion RD1000®, Laser Technology, Inc, Centennial, CO, EUA), con estos datos se calculó el llamado volumen de Pressler (Vp) o volumen de fuste, mediante la ecuación de Pressler (Romahn, 1999) [Ec. 1]; posteriormente, el volumen se transformó a biomasa a través de la densidad de la madera de la especie, equivalente a 490 kg m-3 (Ordóñez et al., 2015).
Donde: Vp = volumen de Pressler (m3), g = área basal de la sección a 1.3 m de altura (m2), P = altura de Pressler = Hp + 1.95 (m), Hp = distancia entre la sección transversal a 1.3 m de altura y la sección con diámetro igual a la mitad del diámetro normal (m).
Ajuste del modelo
Con los datos de biomasa (variable dependiente) y el diámetro normal (variable independiente) se ajustó la ecuación alométrica potencial [Ec. 2] en su forma lineal [Ec. 3]. El modelo fue ajustado a través del método de mínimos cuadrados ordinarios (Montgomery et al., 2006) con el paquete stats del software R (R Core Team, 2019); también se obtuvo el factor de corrección (FC) por el sesgo obtenido al realizar la transformación logarítmica y regresarla a las unidades originales (Sprugel, 1983). La ecuación basada sólo en diámetro como variable independiente ha demostrado tener buenas predicciones de biomasa en varias especies forestales (Altanzagas et al., 2019; Avendaño et al., 2009; Feller, 1992; Flores-Nieves et al., 2011; Návar, 2009; Razo-Zárate et al., 2013); de acuerdo con la revisión de Picard et al. (2012), ésta es la más frecuentemente utilizada para predecir biomasa, puede interpretarse biológicamente y en su forma logarítmica es posible reducir la heterocedasticidad (Sprugel, 1983); además, por la naturaleza de los datos, describe perfectamente la relación de variables dasométricas con la biomasa (Picard et al., 2012).
Donde: Bf = biomasa de fuste (kg), Dn = diámetro normal (cm), ln = logaritmo natural, β0 y β1 = coeficientes de regresión.
Validación del modelo
El análisis de los supuestos del modelo de regresión lineal (normalidad, homogeneidad de varianza e independencia de los errores) es esencial para verificar si el modelo estimará correctamente la variable dependiente (Faraway, 2009; Fox, 2016; Montgomery et al., 2006). Para evaluar la normalidad se utilizó el paquete tseries de R (Trapletti y Hornik, 2019) a través de la prueba de Jarque-Bera, y el paquete lmtest (Zeileis y Hothorn, 2002) para la homogeneidad de varianza (Test de Breusch-Pagan) e independencia de los errores (Test de Durbin-Watson). El diagnóstico de observaciones atípicas e influyentes (distancia de Cook y Leverage) se evaluó mediante el paquete stats (Fox y Weisberg, 2019).
Resultados y discusión
Para el presente estudio se muestrearon 110 árboles, cuyos diámetros fluctuaron de 7.8 a 87.0 cm y las alturas de 4.7 a 36.0 m (Cuadro 1). Para este estudio se abarcaron categorías diamétricas desde 5 hasta 85 cm; el árbol más grande registró 3233.31 kg de biomasa (6.59 m3), mientras que el más pequeño sólo 6.79 kg, esta variable presentó un coeficiente de variación de más de 102.6 %; en altura, este valor ascendió sólo a 31.2 %. En esta misma especie, Guerra et al. (2012) reportaron diámetros desde 85 cm en Emiliano Zapata, Villarreal Terrenate, La Rosa Terrenate en Tlaxcala y Cuatexmola, Ixtacamaxtitlán, Puebla, hasta 110 cm en La Caldera, Ixtacamaxtitlán, Puebla. En su estudio, Návar (2009) reportó diámetros de 8.4 a 49.8 cm en el noreste México. Domínguez et al. (2004) registraron alturas hasta de 45 m en Pinal de Amoles, Querétaro, mientras que Van Pelt y Nadkarni (2004) midieron árboles con diámetros de 300 cm y 90 m de altura en una porción del sureste de las cascadas de Washington.
Cuadro 1 Estadísticos descriptivos de variables de Pseudotsuga menziesii (Mirb.) Franco en San Antonio de las Alazanas, Coahuila.
| Variable | Media | Mín | Máx | IC-M | Q10 | Mediana | Q90 | Rango | DE | CV (%) | RI |
| Dn (cm) | 40.80 | 7.80 | 87.00 | ±3.73 | 16.91 | 38.15 | 65.30 | 79.20 | 19.57 | 47.98 | 33.00 |
| Hp (m) | 9.80 | 3.70 | 16.50 | ±0.57 | 5.97 | 9.70 | 13.50 | 12.80 | 2.96 | 30.21 | 4.00 |
| Ht (m) | 19.24 | 4.70 | 36.00 | ±1.15 | 10.79 | 19.10 | 26.86 | 31.30 | 6.00 | 31.19 | 7.50 |
| Vol (m3) | 1.31 | 0.02 | 6.59 | ±0.26 | 0.11 | 0.78 | 2.84 | 6.58 | 1.34 | 102.64 | 1.69 |
| Bf (kg) | 640.32 | 6.79 | 3233 | ±125.36 | 54.88 | 383.67 | 1390.65 | 3226.52 | 657.20 | 102.64 | 830.53 |
Dn: diámetro normal, Hp: altura del punto directriz, Ht: altura total, Vol: volumen del fuste, Bf: biomasa de fuste, IC-M: intervalo de confianza de la media, Q10 y Q90: cuantiles de la variable, DE: desviación estándar, CV: coeficiente de variación, RI: rango intercuartílico.
Avendaño et al. (2009) encontraron que Abies religiosa (H.B.K.) Schl. et Cham. puede almacenar en el fuste hasta 4218.4 kg de biomasa (al diámetro máximo de esta especie, 87 cm), el cual es muy superior a lo que almacena P. menziesii; éstos y otros autores, como Flores-Nieves et al. (2011), indicaron que la proporción de la biomasa de fuste representa entre 80 y 97 % de la biomasa aérea total.
Cumplimiento de los supuestos del modelo
En un análisis preliminar de los residuos estudentizados se detectaron dos observaciones con valores superiores a 3 (Figura 1A), y de acuerdo con Montgomery et al. (2006), éstos son considerados como observaciones atípicas potenciales, mismas que fueron eliminadas; en este sentido, se reajustó nuevamente el modelo con 108 observaciones, de las cuales se obtuvieron resultados sin datos atípicos (Figura 1B).
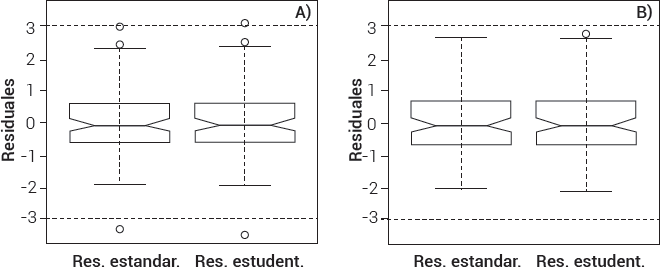
Figura 1 A) datos atípicos detectados a través del análisis preliminar de residuales estandarizados y estudentizados y B) ausentes en el modelo final de Pseudotsuga menziesii (Mirb.) Franco.
La prueba de Jarque-Bera (X2 = 1.724, gl = 2, P = 0.422) y la de Breusch-Pagan (BP = 2.839, gl = 1, P = 0.092) fueron significativas; es decir, los residuos tienen distribución normal (Figura 2A) y son homocedásticos (Figura 2B). Con fines de comprobación estadística de los residuales, la prueba de Durbin-Watson (DW = 1.818, P = 0.147) demostró independencia de los errores; hasta 20 retardos (lags), la autocorrelación de los residuales es < 0.2 (α = 0.5 %) (Figura 2C), con lo que se cumplen los supuestos del modelo lineal, lo cual permite que las estimaciones sean confiables. Algunos autores sólo muestran parámetros de la regresión, R2 aj, error estándar y el CV (Altanzagas et al., 2019; Avendaño et al., 2009; Feller, 1992; Montoya et al., 2018); sin embargo, es recomendable que los modelos cumplan con los supuestos establecidos (Fox, 2016; Montgomery et al., 2006).
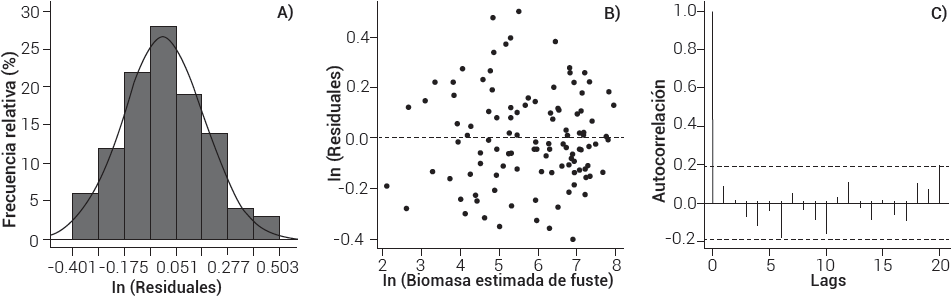
Figura 2 A) histograma de residuales, B) valores estimados contra residuales, C) correlograma de residuales del modelo de Pseudotsuga menziesii (Mirb.) Franco en San Antonio de las Alazanas, Coahuila.
El diagnóstico de observaciones atípicas e influyentes (distancia de Cook y Leverage) indicó que algunas observaciones presentaron influencia potencial (Figura 3), pero sin poner en riesgo las estimaciones. El estadístico de distancia de Cook demostró que tampoco existe influencia real en ninguna observación (Cuadro 2). No se presentó ninguna influencia significativa (P ≤ 0.05) en los estimadores (dfbetas), ni sobre el ajuste del modelo (dffit).
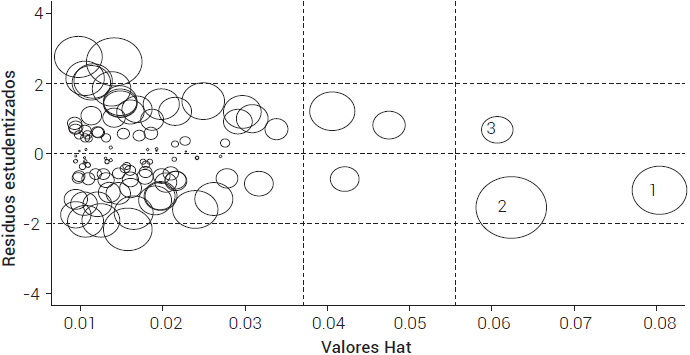
Figura 3 Observaciones influyentes potenciales del modelo de biomasa de fuste de Pseudotsuga menziesii (Mirb.) Franco en San Antonio de las Alazanas, Coahuila.
Cuadro 2 Detección de observaciones influyentes reales y potenciales del modelo para Pseudotsuga menziesii (Mirb.) Franco en San Antonio de las Alazanas, Coahuila.
| Observación | Influencia potencial de observaciones | |||||
| dfb.l | dfb.l(Dn) | dffit | cov.r | cook, d | Hat | |
| 1 | -0.30 | 0.29 | -0.31 | 1.09* | 0.05 | 0.08* |
| 2 | -0.38 | 0.36 | -0.39 | 1.04 | 0.08 | 0.06* |
| 3 | 0.16 | -0.16 | 0.17 | 1.08* | 0.01 | 0.06* |
*: P ≤ 0.05.
Modelos de predicción de biomasa de fuste en Pseudotsuga menziesii
El modelo alométrico para P. menziesii en su forma lineal presentó resultados satisfactorios, el diámetro normal explicó el 98.0 % de la variabilidad de la biomasa de fuste, mientras que en su forma original explicó 96.0 % (Cuadro 3) con 108 árboles y diámetros de 7.8 a 87 cm. En esta misma especie, Espinosa y Perry (1987) mostraron que el diámetro normal explicó en 89 % la biomasa de fuste en árboles jóvenes (22 años), en tanto que el modelo generado por Gower et al. (1987) explicó el 98 %, similar a lo obtenido aquí con el modelo lineal. Návar (2009) y St. Clair (1993) reportaron una R2 de 0.92 en biomasa de fuste, con muestras de 21 y 249 árboles, respectivamente, pero en diámetros de 9 a 26 cm; en cambio, Feller (1992), en Coastal British Columbia, Canadá, obtuvo una R2 de 0.97 y 0.96.0 con el modelo lineal para sitios de buena (n = 46) y mala calidad (n = 42), respectivamente, en diámetros de 5 a 64 cm de P. menziesii. Las estimaciones en todos los casos son aceptables, pero existen diferencias de ajuste entre modelos, lo cual pudiera deberse al tamaño de muestra, al intervalo considerado en las variables dependientes e independientes y a las variaciones de crecimiento de la especie en cada lugar.
Cuadro 3 Parámetros y estadísticos del modelo de biomasa de fuste de Pseudotsuga menziesii (Mirb.) Franco en San Antonio de las Alazanas, Coahuila.
| Ecuación | R2aj | Sxy | CV % | Valor P | FC |
| Bf = 0.0565 × Dn2.4237 | 0.96 | 209.38 | 32.70 | 2.20 × 10-16 | |
| Ln(Bf) = -2.8732(± 0.238) + 2.4237(± 0.066) × ln(Dn) | 0.98 | 0.19 | 3.26 | 2.20 × 10-16 | 1.018 |
Bf: biomasa de fuste (kg), Dn: diámetro normal (cm), R2 aj: coeficiente de determinación ajustado, Sxy: error estándar (kg), CV: coeficiente de variación, ln: logaritmo natural, ±: intervalo de confianza de 95 % del coeficiente de regresión, FC: factor de corrección.
El factor de corrección estimado en este estudio para P. menziesii fue de 1.018 (1.8 %); éste debe calcularse para corregir el sesgo debido a la trasformación logarítmica de las variables, y es multiplicativo a la biomasa estimada (Sprugel, 1983); en los estudios revisados para esta especie se omiten estos valores, o no son calculados, pero en Picea obovata (Altanzagas et al., 2019) y en Pinus halepensis (Montoya et al., 2018) este valor fue de hasta 1.03. Con esta ecuación alométrica es posible conocer el volumen (m3) de fuste de la especie, basta con dividir la biomasa estimada (kg) por la densidad básica (490 kg m-3).
Modelos para predicción de biomasa en otras especies
En otras especies, como Picea obovata Ldb., Altanzagas et al. (2019) reportaron una R2 para biomasa de fuste de 0.971 % con el uso del mismo modelo, con 22 árboles y diámetros de 5.6 a 43.7 cm. En Pinus cembroides Zuc. y P. halepensis Mill. se reportaron R2 de 0.74 y 0.81, respectivamente, en 50 árboles para biomasa de fuste (Marroquín et al., 2018). Otro ejemplo de ello, es en la especie A. religiosa, donde el modelo explicó 92.8 % de la biomasa de fuste en una muestra de 10 árboles con diámetros de 12 a 105 cm (Flores-Nieves et al., 2011). En A. religiosa, Razo-Zárate et al. (2013) reportaron que el diámetro normal explicó 98 %, mientras que Avendaño et al. (2009) demostraron que éste explicó 99.3 % de la biomasa total. Es importante mencionar que el modelo no siempre arroja buenos ajuste; por ejemplo, en P. halepensis el diámetro basal sólo explica 77.0 % de la biomasa total (Montoya et al., 2018).
Los errores y la pendiente (β1) del modelo en Pseudotsuga menziesii (Mirb.) Franco
Para este estudio se tuvo un coeficiente de variación de 32.7 % y error estándar de 209.37 kg, este último fue mayor a lo reportado por Návar (2009) el cual fue de 72.86 kg, lo cual resulta lógico por las diferencias en el tamaño de los árboles muestreados. Feller (1992) y Espinosa y Perry (1987) reportaron un error estándar superior a 0.25 en escala logarítmica, en el presente estudio se obtuvo un valor de 0.19 en esa misma escala. Los coeficientes del modelo (β0 = -2.8732 y β1 = 2.4237) mostraron significancia estadística con P < 0.0001 (Cuadro 3). En los valores originales de las variables el valor estimado de β0 es dependiente del valor mínimo de la variable y, por lo que difiere en cada estudio; β1 define la razón de incremento de y por cada unidad de x, lo que los hace comparables. En modelos potenciales linealizados, St. Clair (1993) obtuvo para P. menziesii una β1 = 2.27, levemente menor a lo obtenido en este estudio, Návar (2009) reportó un valor de 2.59, pero en esta misma especie se han obtenido valores superiores a 2.8 (Espinosa y Perry, 1987; Feller, 1992; Gower et al., 1987), pero estos coeficientes ya no poseen la misma interpretación que tienen en un modelo lineal simple sin trasformación.
La pendiente (β1) del modelo en otras especies
En P. cembroides y P. halepensis se han reportado valores de β1 para biomasa de fuste de 1.80 y 1.51 (Marroquín et al., 2018). Flores-Nieves et al. (2011) reportaron que este valor llega a ser de 2.75 en biomasa de fuste de A. religiosa, pero en Picea obovata el valor de β1 fue de 2.59 (Altanzagas et al., 2019). El valor del parámetro β1 de P. menziesii es similar al de A. religiosa en los estudios de Avendaño et al. (2009) y Razo-Zárate et al. (2013), quienes reportaron valores de 2.5 para biomasa total. En P. halepensis se reportó el valor más bajo (β1 = 0.80), a pesar de ser árboles de grandes dimensiones.
Diferencias de estimación de biomasa entre modelos de Pseudotsuga menziesii (Mirb.) Franco
Las diferencias de biomasa entre modelos muestran que la categoría de 10 cm llega a subestimar otros modelos desde -32.4 % (St. Clair, 1993) hasta -46.3 % (Espinosa y Perry, 1987); sin embargo, en la categoría de 25 cm el modelo de Gower et al. (1987) sobreestimó hasta en 73.5 % más que el modelo presentado aquí. Las predicciones de biomasa que se asemejaron a las obtenidas de este modelo las presentó Feller (1992) y las distantes fueron las de Gower et al. (1987) (Cuadro 4).
Cuadro 4 Comparación de ecuaciones generadas para biomasa de fuste en Pseudotsuga menziesii (Mirb.) Franco.
| CD | Estudios realizados | ||||||||||
| Presente estudio |
Feller (1992) | Dif. (%) | St. Clair (1993) | Dif. (%) | Gower et al. (1987) | Dif. (%) | Espinosa y Perry (1987) | Dif. (%) | Návar (2009) | Dif. (%) | |
| 10 | 14.99 | 15.64 | 4.3 | 10.13 | -32.4 | 18.45 | 23.1 | 8.05 | -46.3 | 13.52 | -9.8 |
| 25 | 138.11 | 125.08 | -9.4 | 135.07 | -2.2 | 239.57 | 73.5 | 122.08 | -11.6 | 144.97 | 5.0 |
CD: categoría diamétrica (cm), Dif.: diferencia con respecto al presente estudio (%).
El modelo generado (Cuadro 3; Figura 4A) es válido y confiable para la predicción de la biomasa de fuste en P. menziesii al usar el diámetro normal en el intervalo indicado, y se debe aplicar en condiciones similares en las que se encuentra la especie (Flores-Nieves et al., 2011; Razo-Zárate et al., 2013). Puesto que se ha evidenciado la sub y sobreestimación de biomasa con los diferentes modelos en relación con modelo obtenido (Cuadro 4; Figura 4B), ésto da pauta para sugerir la inclusión de la altura en este mismo modelo a fin de mejorar las estimaciones, sin omitir el debido cumplimiento de los supuestos de un modelo de regresión.
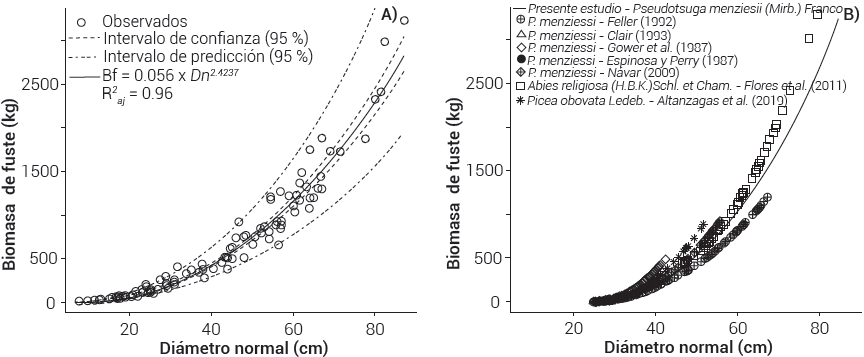
Figura 4 A) valores observados y ajustados del modelo para Pseudotsuga menziesii (Mirb.) Franco, y B) comparado con otros estudios en biomasa de fuste.
A pesar de que se ha demostrado que la cubicación por el método de Pressler sobreestima ligeramente el volumen real, no es estadísticamente diferente, según se detalla a continuación: Lima et al. (2016) evaluaron nueve métodos de cubicación en 98 árboles de P. taeda L. de entre nueve y 20 años de edad e indicaron que todos los métodos de cubicación sobreestiman el volumen de árboles en relación con el volumen real, pero a los 20 años de edad, el método de Pressler es de los cuatro más precisos, incluso para las edades de nueve y 13 años, los volúmenes obtenidos por métodos de Huber, Pressler y Sectional Standard presentaron desviaciones menores en comparación con el volumen real; por su parte, Valencia-Manzo et al. (2017) compararon el volumen real de 134 árboles de P. patula Schl. et Cham. var. longepedunculata Look con el volumen estimado por el método de Pressler y declararon que las diferencias que visualmente presenta la ecuación de Pressler (sobrestimación), en relación con el volumen real, son tan pequeñas que la prueba t-Student determinó que estadísticamente no hay diferencia entre ambos tipos de volumen (Pr > t < 0.05). En un estudio similar, Curto et al. (2019) compararon el volumen de Pressler con el volumen real de Eucalyptus grandis y encontraron que no hay diferencia significativa entre los promedios de los volúmenes estimados por los diferentes métodos en relación al volumen real, según la prueba de Dunnett (P ≤ 0.05).
Conclusiones
El uso de métodos indirectos para la cuantificación de volumen en especies forestales es una forma práctica y de bajo costo que puede ser empleada de manera confiable. La ecuación de Pressler presume ser sencilla y rápida para la cubicación de fuste de árboles en pie de la especie Pseudotsuga menziesii Franco, ya que sólo se requiere medir el diámetro normal y la altura del punto directriz para obtener el volumen y convertir a biomasa, lo que da pauta para incrementar significativamente el tamaño de muestra para generar modelos robustos. El cumplimiento de los supuestos del modelo de regresión: normalidad, homogeneidad de varianzas, independencia de los errores, significancia estadística de los estimadores, identificación y análisis de observaciones atípicas, potenciales e influyentes, aseguran que las predicciones de los modelos de alométricos sean confiables. El diámetro normal demostró ser un buen predictor, ya que explica hasta en 98 % la biomasa de fuste de P. menziesii.

