











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El cacao (Theobroma cacao L.) es uno de los principales recursos agrícolas en el sureste de México; se cultivan 59,675 ha y beneficia directamente a 41,000 familias (SIAP, 2018). Los estados de Tabasco, Chiapas, Guerrero y Oaxaca son los principales productores de cacao; en los dos primeros se concentra el 99 % de la producción nacional. En adición al rezago tecnológico, productivo y agroindustrial que enfrenta este producto prehispánico, a partir del ingreso a territorio nacional del hongo Moniliophthora roreri en 2005, la producción de cacao se ha visto afectada fuertemente con pérdidas que pueden superar el 75 % (Torres et al., 2011). Moniliophthora roreri causa la moniliasis del cacao e infecta únicamente frutos durante todo su desarrollo; son los frutos juveniles los más susceptibles (Griffith et al., 2003). El hongo induce hipertrofia, necrosis interna, madurez prematura y manchas necróticas externas denominadas ‘chocolate’; sobre la mancha crece un estroma blanco con masas de esporas altamente infectivas (Enríquez, 2004).
En México, los estudios sobre este hongo incluyen la confirmación del agente causal, la relación filogenética con aislados centroamericanos (Cuervo-Parra et al., 2011; Dorado et al., 2017; Phillips-Mora et al., 2006) y el efecto de la sombra sobre la incidencia y progreso de la enfermedad (Torres et al., 2011). Adicionalmente, se ha evaluado la eficacia de fungicidas y programas de manejo integrado (Ortíz-García et al., 2015; Torres-de la Cruz et al., 2019); sin embargo, la reciente tendencia a una reactivación productiva del cacao en México y la endemicidad de la enfermedad, con intensidad variable según las condiciones climáticas, obliga a revisar con mayor precisión el efecto de estas condiciones en el desarrollo epidémico y la implicación de diferentes flujos de fructificación durante un ciclo productivo de cacao, con el fin de establecer criterios epidemiológicos para un control efectivo.
En el presente estudio se plantean estos objetivos bajo la premisa de que, en cultivos tropicales, una epidemia asociada a una enfermedad no sistémica, con procesos de infección especializados en órganos vegetativos o productivos debe ser analizada bajo criterios fisiológicos y no únicamente bajo criterios temporales clásicos; es decir, se deben descomponer los efectos de flujos fenológicos en tasas o parámetros epidémicos que permitan establecer criterios efectivos de control y sincronizar su acción con la fase de producción de inóculo, determinante para una epidemia de alta intensidad. Este enfoque probó su viabilidad en el control efectivo de la escoba de bruja en mango (Mangifera indica L.), enfermedad causada por Fusarium spp., al detectar y controlar el principal proceso de infección en el segundo flujo vegetativo respecto al flujo floral (Mora-Aguilera et al., 2003; Noriega-Cantú et al., 1999).
En el sureste de México la producción de frutos de cacao es un proceso continuo durante casi todo el año, con traslape de flujos de producción y amarre de frutos. La especialización infectiva de M. roreri en frutos inmaduros garantiza la culminación de n-ciclos de patogénesis con intra e inter-flujos de producción, lo que permite el progreso de una epidemia aparentemente continua (Torres et al., 2011) e implica efectos epidémicos biológicamente vinculados con la producción de inóculo. Consecuentemente, el analizar una epidemia única e integrada al ciclo productivo anual afecta la comprensión de la estructura epidémica en términos de toma de decisiones para un control racional. Aunque previamente en Costa Rica, en un estudio con una visión agroecológica, se encontró efecto de la densidad de frutos de cacao sin discriminación de edad, así como la sombra arbórea en la ocurrencia de la enfermedad (Gidoin et al., 2014), el enfoque epidemiológico no se ha abordado con rigor para demostrar la importancia de la descomposición epidémica en subprocesos determinados por flujos de fructificación.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se efectuó en seis plantaciones de cacao en el estado de Tabasco y región norte de Chiapas, México; áreas que concentran más del 70 % de la producción en el país (Cuadro 1). La región de estudio tiene clima cálido húmedo con abundantes lluvias en verano, con predominancia de suelos vertisoles y gleysoles. La temperatura media anual es de 26 °C, con mínimos entre 20 y 22 °C en enero, y máximos entre 30 y 34 °C en mayo. La precipitación media anual es de 2432 mm, con periodos de lluvia que se extienden de junio a marzo, y periodo de sequía en abril y mayo (INEGI, 2017). La ubicación geográfica y altitud de cada sitio de muestreo se presentan en el Cuadro 1.
Cuadro 1 Localización de sitios experimentales y número de frutos totales evaluados por localidad para el estudio epidémico de la moniliasis del cacao en Tabasco y norte de Chiapas, México (Mayo 2011-2012).
| Municipio, Estado | Localidad | Latitud N | Longitud O | Altitud (msnm) |
Frutos evaluados |
|---|---|---|---|---|---|
| Paraíso, Tabasco | Oriente | 18° 21’ 06.9” | 93° 12’ 57.8” | 3 | 2077 |
| Cárdenas, Tabasco | Poblado C-28 | 18° 01’ 46.7” | 93° 29’ 42.0” | 3 | 2282 |
| Cunduacán, Tabasco | La Piedra | 18° 07’ 45.2” | 93° 11’ 52.4” | 14 | 1709 |
| Huimanguillo, Tabasco | Paredón | 17° 44’ 59.6” | 93° 23’ 57.8” | 7 | 1404 |
| Jalpa de Méndez, Tabasco | Galeana | 18° 10’ 33.4” | 93° 08’ 48.5” | 12 | 1802 |
| Pichucalco, Chiapas | Platanar | 17° 33’ 01.3” | 93° 18’ 90.9” | 76 | 1347 |
Diseño de la plantación
Se seleccionaron seis plantaciones comerciales tradicionales de cacao de 20 a 25 años de edad. Cada plantación estuvo constituida por la mezcla de híbridos de tipo Trinitario, susceptibles al hongo, con arreglo topológico en marco real (3.5 × 3.5 m) y densidad de 784 plantas/ ha. Los árboles empleados como sombra tuvieron una distribución irregular y correspondieron a moté (Erythrina americana), eritrina (Erythrina fusca), chipilcohíte (Diphysa robinoides) y samán (Samanea saman). En cada plantación se delimitó una parcela experimental de 180 árboles, en un área rectangular de 10 × 18 árboles de cacao; de los cuales, se evaluó la parcela central de 4 × 12 árboles. En todas las parcelas, el manejo de plantaciones consistió únicamente en control mecánico de maleza, dos veces en el ciclo productivo, y eliminación mecánica de brotes vegetativos basales.
Registro de la formación de frutos
Los frutos con medidas de 2 a 8 cm de longitud se registraron semanalmente del 18 de mayo de 2011 al 9 de mayo de 2012 para establecer la densidad de frutos potenciales para la infección natural de M. roreri y su relación con la incidencia de la enfermedad.
Evaluación de incidencia de la enfermedad
Theobroma cacao produce frutos de manera continua durante casi todo el año, por lo cual, en cada parcela, de mayo 2011 a abril 2012, se etiquetaron todos los frutos de 8 a 10 cm de largo, provenientes de seis flujos de producción y amarre de frutos que se presentaron durante un ciclo completo de producción de cacao. El número de frutos por parcela se obtuvo mediante un censo de los 48 árboles y el número de frutos totales evaluados por parcela varió de 1347 a 2077 (Cuadro 1). Semanalmente, frutos individuales se observaron para detectar síntomas y signos de M. roreri consistentes en manchas tipo chocolate y esporulación. Se registró el número de frutos enfermos y sanos para estimar la incidencia de enfermedad. Frutos necróticos con presencia de esporas se eliminaron mensualmente con el fin mantener el efecto principal del inóculo externo sobre la parcela experimental y reducir el efecto de reinfección.
Análisis temporal del progreso de la moniliasis del cacao
Se generaron gráficas por parcela del progreso temporal del porcentaje de incidencia, acumulada y no acumulada (incrementos absolutos), en función del tiempo (semanas) total del ciclo productivo. También se generaron gráficas en función al progreso de la enfermedad por cada flujo de fructificación (tiempo parcial). Se analizó la curva del progreso acumulado de incidencia en el ciclo productivo total; además, se analizaron las epidemias parciales por cada flujo de fructificación.
Para la curva de epidemia total y curvas parciales se estimaron la tasa de infección aparente (r) con la inversa del parámetro b del modelo de Weibull, área bajo la curva de progreso de enfermedad (ABCPE), incidencia inicial (Y 0) e incidencia acumulada final (Yf) (Campbell y Madden, 1990). Para comparar tasas de infección aparente se empleó el error estándar del estimador para la aplicación de la prueba de t (P = 0.05). Para estimar el efecto de inóculo primario y secundario en la forma de la curva (monocíclico versus policíclico), la epidemia total y parciales se ajustaron a los modelos epidemiológicos Gompertz, logístico y monomolecular en su forma no lineal. El coeficiente de determinación se empleó como criterio de bondad de ajuste. La tasa epidémica de estos modelos no se consideró para el análisis por su inviabilidad comparativa; por lo que el modelo de Weibull se usó para este fin.
Variables climáticas
En cada parcela, la temperatura y la humedad relativa se registraron cada 2 h mediante data loggers Hobo H8® (Onset Instruments, Pocasset, MA, EUA), los cuales se instalaron en el centro de cada parcela experimental a 2 m de altura. Los datos se recolectaron semanalmente. Con la finalidad de estimar condiciones favorables para el desarrollo de la enfermedad, se correlacionó la incidencia absoluta con intervalos de temperatura y humedad relativa, desfasadas a intervalos semanales, dentro del rango del periodo de incubación; para este fin, se calculó el número de horas/semana que cumplieron con las siguientes condiciones: temperatura a) < 19.9 °C; b) 20-26.9 °C; c) 27-29.9 °C; d) 30-34.9 °C, y e) > 35 °C; y humedad relativa a) < 59.9 %, b) 60-90 %, y c) > 90 %.
Análisis estadístico
Los cálculos estadísticos de la tasa de infección aparente (r), ABCPE y el ajuste de los datos de incidencia a modelos epidemiológicos se realizaron con procedimientos NLIN, SUMMARY y GLM de SAS® (SAS Institute, 2004). El ajuste a modelos epidemiológicos se evaluó mediante el coeficiente de determinación, significancia de parámetros y del modelo. La distribución de los datos se verificó con el estadístico de Kolmogorov-Smirnov. Debido a la falta de normalidad, se calculó el coeficiente de correlación de Spearman entre la incidencia, variables de clima y número de frutos, mediante el software estadístico IBM SPSS® 22.0. La relación lineal del número total de frutos con el ABCPE, tasa de infección aparente y Y f, se analizó con el coeficiente de correlación de Pearson (r), mediante el procedimiento PROC CORR de SAS®.
RESULTADOS
Flujos productivos y condiciones ambientales
La densidad de fructificación mostró dos periodos de mayor producción (Figura 1). El primer periodo se presentó del 29 de junio al 12 de octubre de 2011 y el segundo del 25 de enero al 28 de marzo de 2012. Por otro lado, la temperatura, humedad relativa y el número de frutos presentaron fluctuación temporal similar en todas las parcelas evaluadas, donde la temperatura fue la que presentó la menor variación interparcelaria (Figura 1).
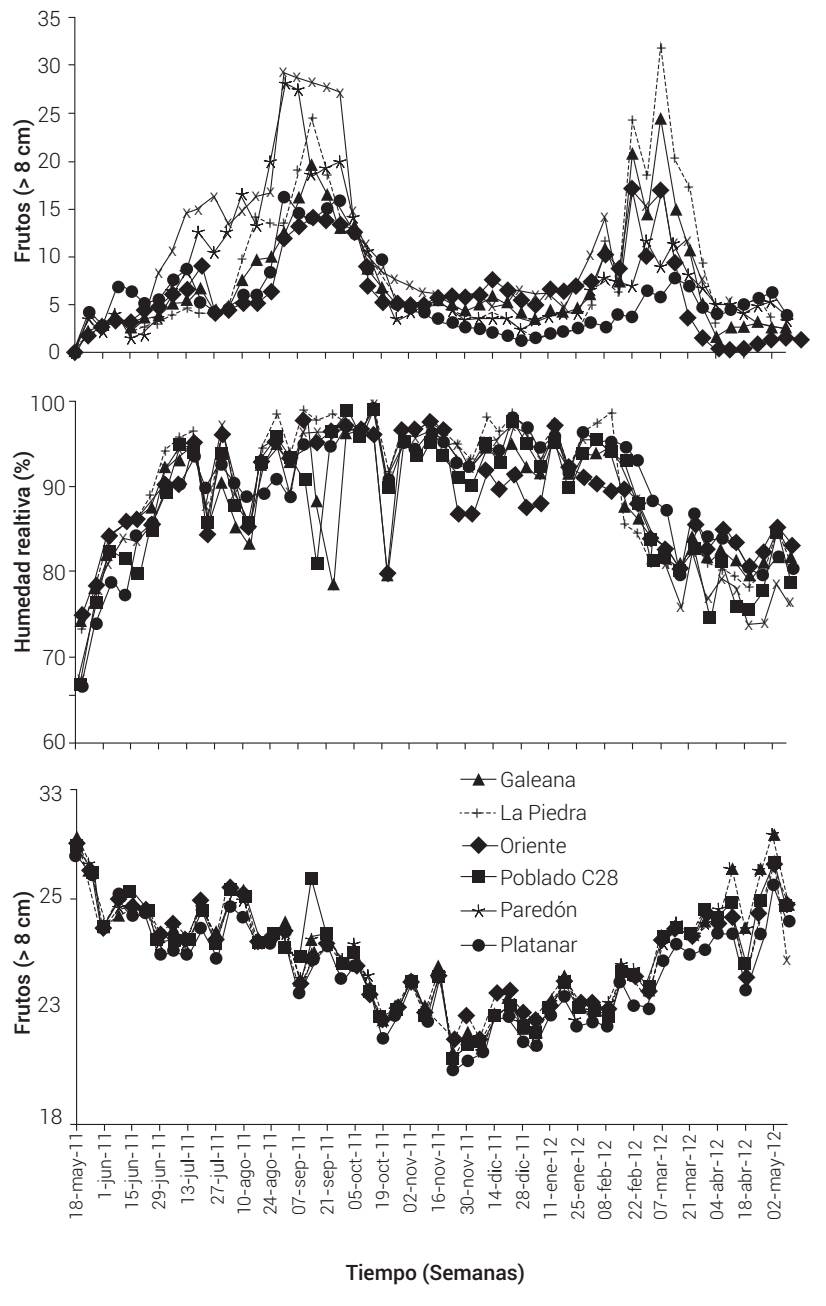
Variables de clima versus incidencia y número de frutos
La incidencia anual no acumulada o absoluta de frutos con moniliasis se relacionó positivamente con periodos de temperatura en el intervalo de 20 a 26.9 °C (r = 0.410.87, P < 0.05) y con periodos de humedad relativa mayor al 60 % (60-90 % y > 90 %) (r = 0.35-0.67, P < 0.05) (Cuadro 2). La mejor relación se presentó con un desfase de 28 a 35 d previo a la aparición del síntoma inicial (mancha chocolate). La incidencia absoluta también se relacionó positivamente con el proceso fenológico de amarre de frutos (r = 0.55-0.82, P < 0.05), 63 d antes de la aparición de síntomas (Cuadro 2). El periodo de mayor incidencia mostró relación positiva con el primer periodo de máxima producción de chilillos (r = 0.41-0.73) desfasados de 35 a 56 d (Cuadro 2). Por otro lado, el número de frutos totales por parcela correlacionó positivamente, sin desfase, con ABCPE (r = 0.85), tasa de infección aparente (r = 0.95) y Y f (r = 0.86).
Cuadro 2 Correlación de Spearman entre la incidencia absoluta de moniliasis del cacao e intervalos de temperatura y humedad relativa desfasados entre 28 y 35 d, y con el número de frutos (producción y amarre) desfasados 63 d en seis parcelas comerciales de Tabasco y norte de Chiapas. Ciclo productivo 2011-2012.
| Localidad, Estado | Correlación de Spearman (Rho) | ||||
|---|---|---|---|---|---|
| Intervalo de temperatura | Intervalo de humedad | Producción de frutos |
Máximo amarre de frutos |
||
| 20-26.9 °C | 60-90 % | > 90 % | |||
| Galeana, Tabasco | 0.77 | 0.58 | 0.55 | 0.80 | 0.66 |
| La Piedra, Tabasco | 0.60 | 0.52 | 0.38 | 0.72 | 0.70 |
| Oriente, Tabasco | 0.77 | 0.40 | 0.61 | 0.70 | 0.53 |
| Poblado C-28, Tabasco | 0.50 | 0.35 | 0.43 | 0.82 | 0.63 |
| Paredón, Tabasco | 0.87 | 0.48 | 0.67 | 0.55 | 0.73 |
| Platanar, Chiapas | 0.41 | 0.57 | 0.54 | 0.72 | 0.41 |
Análisis de epidemia total y ajuste a modelos epidemiológicos
Se conformaron seis epidemias integradas al ciclo anual productivo, una por cada parcela estudiada (Figura 2). En todas las epidemias el inicio de enfermedad (Y 0) e incidencia final (Yf) se observaron en la semana correspondiente al 27 de junio de 2011 y 9 de mayo de 2012, respectivamente. Considerando el ciclo anual, el progreso de la enfermedad ocurrió durante 42 semanas (10 meses) y la mayor incidencia se presentó entre los meses de octubre a diciembre. Las epidemias se caracterizaron por una conformación sigmoidal atípica con fase asintótica inferior y superior cortas, de 5 a 7 y 2 semanas respectivamente, y un débil punto de inflexión aproximadamente a las 22 semanas, correspondiente al mes de diciembre (Figura 2).
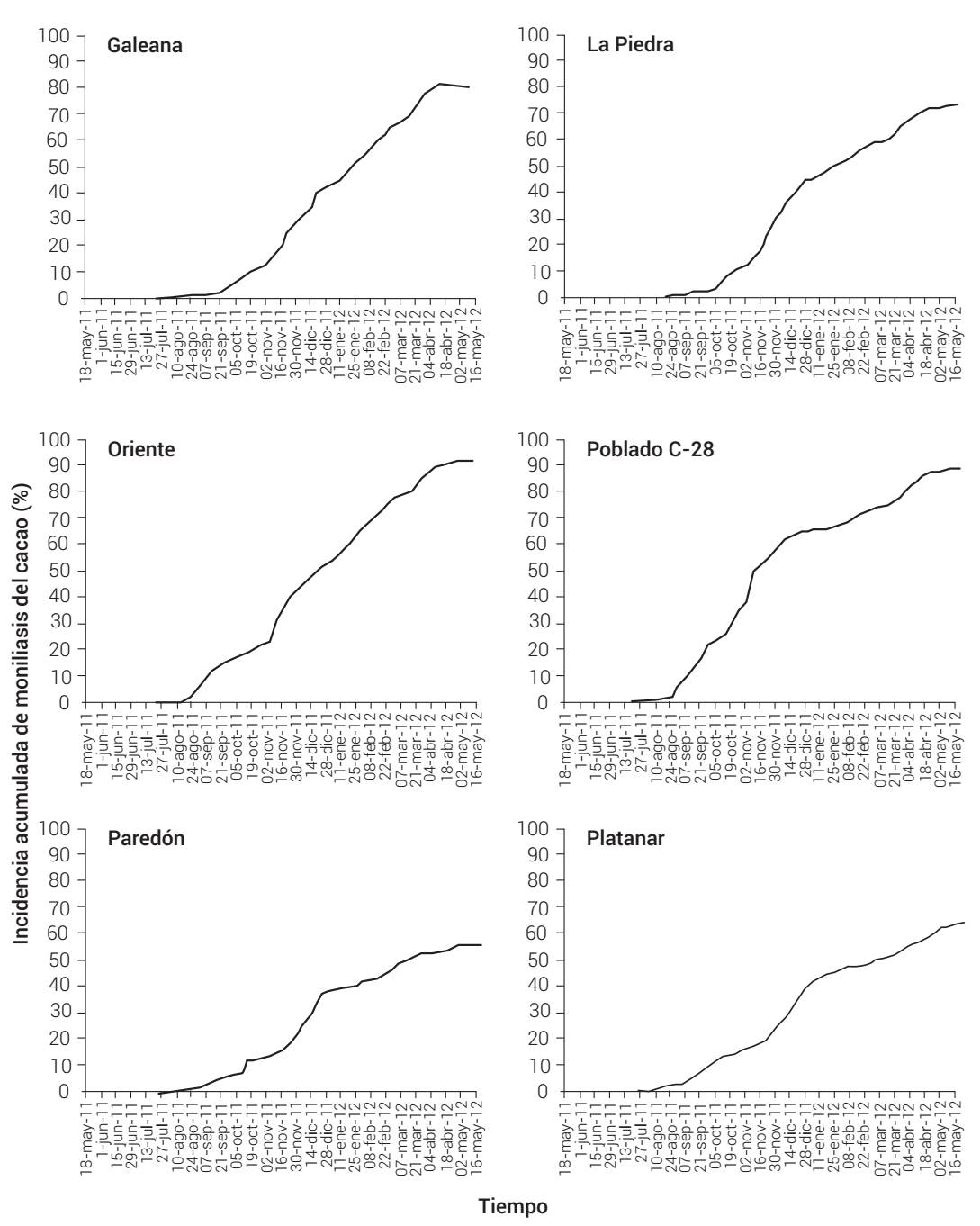
Figura 2 Curvas epidémicas del progreso temporal acumulado de incidencia de moniliasis del cacao causada por Moniliophthora roreri en seis plantaciones comerciales tradicionales de Tabasco y norte de Chiapas, México. Ciclo productivo 2011-2012.
La Yf de la enfermedad fluctuó de 54.49 a 88.73 %, con una incidencia promedio de 75.8 % para la región (Cuadro 3). La menor Y f se presentó en Paredón, Huimanguillo, Tabasco, mientras que la mayor Yf se obtuvo en la parcela Oriente del Municipio de Paraíso (Cuadro 3, Figura 2).
Cuadro 3 Parámetros epidemiológicos del progreso de la incidencia acumulada durante el ciclo total productivo y ajuste a modelos epidemiológicos de la moniliasis del cacao (Moniliophthora roreri) en seis localidades del estado de Tabasco y norte de Chiapas, México. Ciclo productivo 2011-2012.
| Epidemia | ABCPE | Tasa | Yf | Modelo | R2 | |
|---|---|---|---|---|---|---|
| r | R2 | |||||
| Oriente | 12,937.12 | 0.005 | 0.99 | 88.73 | Logístico | 0.99 |
| La Piedra | 10,262.49 | 0.004 | 0.98 | 75.66 | Gompertz | 0.99 |
| Poblado C-28 | 14,132.83 | 0.006 | 0.97 | 85.45 | Logístico | 0.99 |
| Paredón | 8186.08 | 0.003 | 0.95 | 54.49 | Gompertz | 0.98 |
| Galeana | 8508.85 | 0.005 | 0.99 | 86.94 | Gompertz | 0.97 |
| Platanar | 9617.30 | 0.003 | 0.97 | 63.77 | Logístico | 0.96 |
ABCPE: área bajo la curva del progreso de la enfermedad, r: tasa de infección aparente de la enfermedad estimado con el modelo de Weibull, Y f: incidencia final acumulada, R2: coeficiente de determinación.
Los valores de intensidad de las epidemias anuales estimados con ABCPE fluctuaron entre 8186 en Paredón y 14,132 en Poblado C-28 (Cuadro 2). Estas mismas parcelas también tuvieron los valores más bajos y altos de tasa de infección aparente (r) con 0.003 y 0.006, respectivamente. Las formas de las curvas de epidemias anuales de tres localidades se ajustaron al modelo logístico y tres se ajustaron al modelo Gompertz (Cuadro 2, Figura 2).
Análisis de epidemias parciales y ajuste a modelos epidemiológicos
El análisis parcial epidémico por flujo de fructificación permitió obtener seis sub-epidemias por ciclo productivo y localidad. Todas las sub-epidemias presentaron traslape debido a la sucesión de etapas de fructificación del cacao (Figura 3). La Y f de cada sub-epidemia fue variable, con tendencia creciente en tiempo; así, en todas las parcelas estudiadas el flujo de fructificación 1 (G1) mostró la menor Y f. También se observó un incremento gradual del valor de este parámetro hasta G5 o G6 (Cuadro 4). De forma similar, los valores de ABCPE de cada sub-epidemia fueron variables; sin embargo, en todas las parcelas evaluadas G1 y G5 mostraron la menor y mayor intensidad, respectivamente, excepto en Galeana (Cuadro 4).
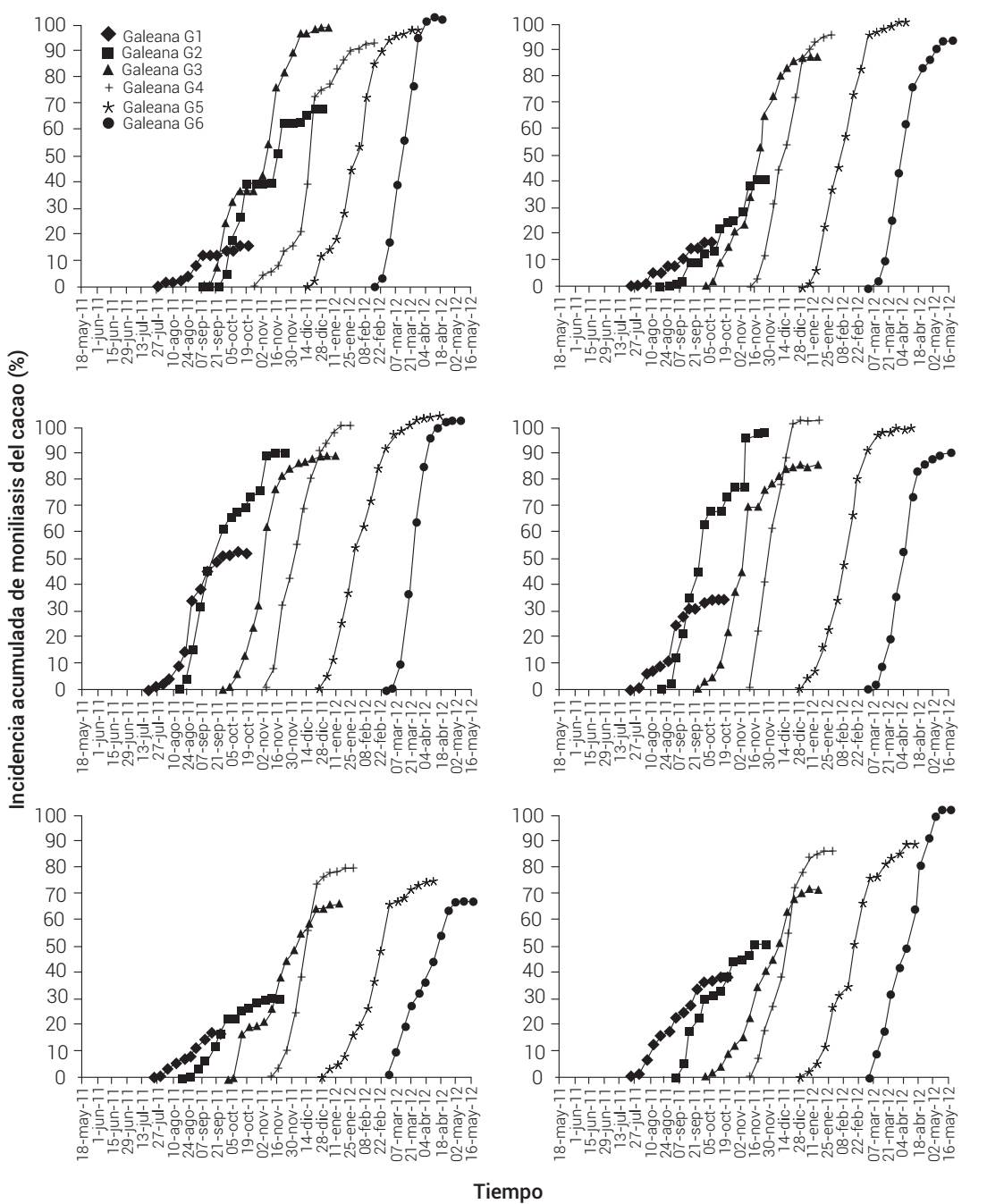
Figura 3 Curvas de incidencia acumulada del progreso temporal de seis sub-epidemias de moniliasis del cacao (Moniliophthora roreri) asociadas a seis etapas sucesivas de fructificación del cacao en seis localidades del estado de Tabasco y norte de Chiapas. Ciclo productivo 2011-2012. Gn: flujo de fructificación-n.
Cuadro 4 Área bajo la curva de progreso de la enfermedad, incidencia final acumulada, tasa de infección aparente, modelos epidémicos con mejor bondad de ajuste y correlación de incidencia absoluta (Y i) con respecto a intervalos de temperatura y humedad relativa de seis sub-epidemias de moniliasis del cacao (Moniliophthora roreri) en seis localidades de Tabasco y norte de Chiapas, México. Ciclo productivo 2011-2012.
| Localidad (Estado) |
Sub-epidemia | ABCPE | Yf | Tasa | Modelos | Correlación (Rho) Yi vs: | |||
|---|---|---|---|---|---|---|---|---|---|
| r | R2 | Mejor ajuste | r2 | T (°C) 20-26.9 |
HR (%) 60-90 |
||||
| Galeana (Tabasco) | G1 | 597 | 14.2 | 0.022c | 0.91 | Monomolecular G | 0.92 | 0.80 | 0.66 |
| G2 | 3690 | 42.2 | 0.018a | 0.91 0.96 0.91 0.91 | Gompertz | 0.95 | 0.79 | 0.8 | |
| G3 | 5816 | 69.0 | 0.019b | 0.91 | Logístico | 0.93 | 0.79 | 0.59 | |
| G4 | 4948 | 77.9 | 0.017a | 0.91 | Logístico | 0.98 | 0.89 | 0.88 | |
| G5 | 5109 | 90.7 | 0.019b | 0.94 | Logístico | 0.99 | 0.76 | 0.94 | |
| G6 | 2978 | 96.8 | 0.033d | 0.99 | Logístico | 0.97 | 0.54 | 0.60 | |
| La Piedra (Tabasco) | G1 | 531 | 16.5 | 0.022d | 0.92 | Monomolecular | 0.97 | 0.67 | 0.94 |
| G2 | 1397 | 39.7 | 0.015a | 0.95 | Gompertz | 0.97 | 0.79 | 0.66 | |
| G3 | 4506 | 84.4 | 0.019b | 0.93 | Logístico | 0.99 | 0.60 | 0.75 | |
| G4 | 3626 | 92.1 | 0.026e | 0.93 | Logístico | 0.99 | 0.64 | 0.81 | |
| G5 | 5192 | 97.5 | 0.021c | 0.93 | Logístico | 0.98 | 0.77 | 0.73 | |
| G6 | 3553 | 90.8 | 0.023d | 0.94 | Logístico | 0.99 | 0.62 | 0.81 | |
| Oriente (Tabasco) | G1 | 2198 | 50.4 | 0.023b | 0.91 | Gompertz | 0.93 | 0.54 | 0.6 |
| G2 | 4724 | 85.8 | 0.025c | 0.97 | Monomolecular | 0.97 | 0.49 | 0.83 | |
| G3 | 5212 | 85.8 | 0.020a | 0.94 | Logístico | 0.99 | 0.05 | 0.62 | |
| G4 | 4184 | 96.8 | 0.031d | 0.99 | Logístico | 0.97 | 0.82 | 0.82 | |
| G5 | 6719 | 100.0 | 0.023b | 0.99 | Logístico | 0.97 | 0.67 | 0.49 | |
| G6 | 3670 | 98.6 | 0.035e | 0.99 | Logístico | 0.98 | 0.82 | 0.56 | |
| Poblado C-28 (Tabasco) | G1 | 1569 | 32.8 | 0.023c | 0.91 | Monomolecular | 0.93 | 0.40 | 0.66 |
| G2 | 4640 | 82.6 | 0.021b | 0.97 | Gompertz | 0.97 | 0.71 | 0.40 | |
| G3 | 5361 | 94.6 | 0.022c | 0.99 | Gompertz | 0.99 | 0.84 | 0.41 | |
| G4 | 3680 | 99.2 | 0.046e | 0.99 | Logístico | 0.96 | 0.92 | 0.60 | |
| G5 | 5476 | 95.9 | 0.020a | 0.93 | Logístico | 0.98 | 0.69 | 0.74 | |
| G6 | 3334 | 86.5 | 0.028d | 0.93 | Logístico | 0.99 | 0.80 | 0.63 | |
| Paredón (Tabasco) | G1 | 414 | 17.0 | 0.018a | 0.96 | Gompertz | 0.97 | 0.71 | 0.47 |
| G2 | 1448 | 29.6 | 0.022c | 0.92 | Monomolecular | 0.96 | 0.91 | 0.68 | |
| G3 | 3519 | 65.2 | 0.019b | 0.95 | Gompertz | 0.97 | 0.60 | 0.40 | |
| G4 | 3296 | 73.2 | 0.021c | 0.92 | Gompertz | 0.98 | 0.40 | 0.64 | |
| G5 | 3766 | 77.7 | 0.018ab | 0.94 | Gompertz | 0.98 | 0.48 | 0.41 | |
| G6 | 2668 | 66.2 | 0.026d | 0.91 | Gompertz | 0.98 | 0.76 | 0.56 | |
| Platanar (Chiapas) | G1 | 1679 | 35.5 | 0.021d | 0.94 | Monomolecular | 0.99 | 0.63 | 0.90 |
| G2 | 2244 | 46.8 | 0.030e | 0.97 | Monomolecular | 0.97 | 0.56 | 0.74 | |
| G3 | 3104 | 67.5 | 0.017a | 0.94 | Gompertz | 0.99 | 0.65 | 0.77 | |
| G4 | 3335 | 81.3 | 0.027d | 0.92 | Gompertz | 0.99 | 0.73 | 0.76 | |
| G5 | 4442 | 83.3 | 0.018b | 0.95 | Gompertz | 0.99 | 0.65 | 0.70 | |
| G6 | 3527 | 95.8 | 0.026d | 0.95 | Logístico | 0.97 | 0.41 | 0.57 | |
†Tasas con la misma letra, dentro de cada localidad, son estadísticamente iguales (P = 0.05). ABCPE: área bajo la curva del progreso de la enfermedad, Y f : incidencia final acumulada, R: tasa de infección aparente de la enfermedad/modelo de Weibull, R2 : coeficiente de determinación, T: temperatura, HR: humedad relativa, Rho: Correlación de Spearman, Y i : incidencia absoluta al tiempo-i.
El modelo epidémico con mejor bondad de ajuste y la tasa de infección aparente fue variable en todas las subepidemias y localidades (P ≤ 0.05) (Cuadro 4). En Oriente, Poblado C-28 y Paredón, la sub-epidemia con mayor tasa fue la G6. En cinco localidades cinco de las seis subepidemias se ajustaron a funciones sigmoides (Gompertz o logístico). En cuatro localidades la G1 se ajustó al modelo monomolecular y en las dos restantes al Gompertz (Cuadro 4).
Variables de clima versus incidencia y número de frutos en epidemias parciales
Todas las sub-epidemias se relacionaron positivamente con intervalos de temperatura entre 20 y 26.9 °C, y con intervalos de humedad relativa mayor a 60 % (Cuadro 4). Al igual que en las epidemias anuales, esta asociación presentó un desfasamiento de 28 a 35 d antes de la aparición de la mancha tipo chocolate.
DISCUSIÓN
La producción de cacao en México se ha afectado fuertemente a partir del ingreso de M. roreri en 2005 (Phillips-Mora et al., 2006; Torres et al., 2011); sin embargo, los estudios no fueron extensivos para cuantificar el efecto epidémico. Este trabajo evidenció que la moniliasis tuvo una incidencia regional promedio de 75.8 % asociada al ciclo productivo anual en la principal área cacaotera del país, lo cual muestra la capacidad endémica del hongo, al igual que en otros países como Costa Rica, Ecuador y Colombia, donde se han reportado pérdidas superiores al 80 % (Hidalgo et al., 2003). El potencial parasítico del M. roreri se favoreció, pero no se limitó, por periodos predominantes con temperaturas entre 20 y 26.9 °C y humedad relativa mayor al 60 % (P ≤ 0.05), condiciones prevalentes en el trópico. Estos resultados tienen concordancia con varios estudios relacionados con la patogénesis de la enfermedad. Porras y Galindo (1985) reportaron que la esporulación de M. roreri depende de la humedad y la temperatura (20 a 27 °C). De acuerdo con Leandro-Muñoz et al. (2017), M. roreri requiere temperaturas de 20 a 27 °C y humedad entre 80 y 100 % para la germinación de esporas y penetración al tejido del hospedante.
La relación positiva de la incidencia de moniliasis con la emisión de frutos y la correlación positiva del número de frutos totales con incidencia final (Y f ), ABCPE y tasa de infección aparente demuestran la importancia de la densidad poblacional de frutos en la intensidad epidémica. Previamente, Gidoin et al. (2014) encontraron que la cantidad de frutos de cacao es importante para explicar la incidencia de la enfermedad; sin embargo, estos autores no demostraron el efecto directo de esta variable sobre parámetros epidémicos ni su implicación en el incremento de inóculo primario y secundario debido a la ausencia de trazabilidad de fructificación. En esta investigación, el marcado de frutos permitió discriminar frutos entre flujos productivos para el análisis y parametrización independiente de sub-epidemias en adición a la epidemia total anual.
La asociación de la incidencia absoluta de la epidemia anual con el proceso fenológico de amarre de frutos presentó un desfasamiento de 65 d; así también, la asociación de incidencia absoluta con el primer periodo de máxima producción total de chilillos (frutos jóvenes) se presentó con un desfase de 35 a 56 d. Lo anterior se explica por el largo periodo de incubación de M. roreri. Evans (1981) y Porras y Enríquez (1998) demostraron que la aparición de los síntomas de moniliasis ocurre de 40 a 75 días después de la infección. Los valores encontrados en esta investigación también concuerdan con la fase asintótica inicial de 5 a 7 semanas (35-49 d) antes del primer incremento en la curva epidémica; en consecuencia, la emisión y cantidad de frutos puede ser empleada en modelos de alerta temprana en conjunción con horas favorables semanales de temperatura y humedad relativa.
Las curvas epidémicas anuales se ajustaron a los modelos logístico y Gompertz; estos modelos describen epidemias policíclicas, las cuales originan curvas de progreso sigmoidales (Campbell y Madden, 1990); sin embargo, las curvas anuales exhibieron una condición sigmoidal atípica, principalmente por una débil definición del punto de inflexión, el cual se observa en las subepidemias definidas por el flujo de fructificación ajustadas por los mismos modelos (Figuras 2 y 3). Esta condición atípica se puede atribuir a las variaciones en los ritmos de amarre de frutos, los cuales incrementan la intensidad de la epidemia de manera diferencial, lo que justifica su análisis particionado.
El análisis parcial de las epidemias anuales, definidas por flujos productivos, evidenció un incremento gradual epidémico en todos los parámetros estimados con un inicio monocíclico, seguido de eventos policíclicos. En epidemias monocíclicas el patógeno exhibe un único proceso infeccioso en un periodo corto, mientras que en policíclicas, el hongo tiene la capacidad de producir nuevo inóculo infectivo y la epidemia progresa con incrementos graduales intermediada con una fase explosiva o exponencial (Campbell y Madden 1990). Esta multiplicidad de ciclos es favorecida por la permanencia de frutos infectados adheridos al árbol, los cuales pueden proveer inóculo para infecciones continuas durante el ciclo productivo (Hernández-Gómez et al., 2012; Torres et al., 2011).
La predominancia de epidemias policíclicas entre el tercer y sexto flujo productivo es relevante para el establecimiento de estrategias de manejo que incluyan la reducción del inóculo secundario para retrasar o disminuir la intensidad de la tasa epidémica (Campbell y Madden, 1990); así, la eliminación periódica de frutos infectados en etapa pre-esporulativa y la cosecha oportuna de frutos puede tener un impacto negativo sobre el inóculo secundario (Soberanis et al., 1999). De forma similar, la predominancia de sub-epidemias monocíclicas es un indicativo de un efecto predominante del inóculo primario en los primeros flujos de fructificación (G1 y G2), lo cual justifica acciones que reduzcan el inóculo primario y que retrasen la epidemia al afectar el parámetro Y o (Campbell y Madden, 1990); en este caso, la eliminación de frutos momificados o la eliminación total de frutos en época de baja producción (denominada purga) puede resultar en disminución del inóculo primario (Leandro-Muñoz et al., 2017; Torres et al., 2011).
El análisis de la estructura epidémica de la moniliasis reveló que durante un ciclo productivo existe en realidad una serie sucesiva de epidemias encriptadas, con incremento gradual de intensidad, debido al aporte de inóculo de sub-epidemias previas; razón por la cual esta epidemia puede ser de alta intensidad y requerir un programa de manejo continuo. Por otro lado, la reinfección dentro de una epidemia está garantizada con la liberación, hasta por siete meses, de grandes cantidades de inóculo que puede producir un fruto infectado (Enríquez, 2004); por consiguiente, el control de este hongo requiere acciones complementarias a las dirigidas a reducir Y o y que actúen directamente sobre los parámetros de intensidad epidémica (i.e. r, ABCPE, Y f ) mediante acciones sanitarias que incluyan la protección de frutos por flujos reproductivos, dando prioridad a los frutos de 1 a 2 meses de edad por su mayor susceptibilidad. En este caso, el uso de fungicidas de contacto o sistémicos, eficaces contra la moniliasis, puede coadyuvar en la protección de frutos (Bateman et al., 2005; Leandro-Muñoz et al., 2017, Torres de la Cruz et al., 2019).
La influencia inductiva de temperaturas cálidas y alta humedad relativa sobre la intensidad epidémica sugieren la importancia de manejar el microclima de la plantación de cacao con medidas como poda de aireación, eliminación de renuevos, control de arvenses, drenaje efectivo y el establecimiento de una sombra uniforme y moderada (Bateman et al., 2005; Gidoin et al., 2014; Ortíz-García et al., 2015); sin embargo, estas medidas pueden ser limitadas debido a la plasticidad climática encontrada en este trabajo, particularmente por los bajos requerimientos de humedad del hongo para infectar (> 60 %), por lo que deben complementarse con la inclusión de sistemas de alertas tempranas.
CONCLUSIONES
El estudio de la estructura epidémica de la moniliasis del cacao en el sureste de México en función de flujos productivos y eventos climáticos reveló que la enfermedad ocurrió durante 10 meses consecutivos, con una incidencia promedio de 75.8 % y mayor incidencia entre octubre y diciembre. La enfermedad fue favorecida por periodos de temperaturas de 20 a 26.9 °C, por humedad relativa mayor al 60 % y por la disponibilidad de frutos susceptibles. La epidemia anual progresa a través de una sucesión de subepidemias determinadas por flujos de fructificación que se comportan como una enfermedad policíclica. La dinámica de la moniliasis por flujos productivos evidenció una fase monocíclica inicial dependiente del inóculo primario, y fases policíclicas posteriores dependientes de ciclos de los inóculos primario y secundario. La mayor intensidad epidémica se alcanzó en el quinto flujo productivo (eneromarzo) determinado por la densidad de frutos. Las estrategias de manejo deben enfocarse a operar sobre inóculo inicial (Y o ) y parámetros de intensidad epidémica (i.e. r, ABCPE). Este trabajo constituye el primer reporte que demuestra las implicaciones de flujos productivos en la estructura epidémica de la moniliasis del cacao.

