











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El daño mecánico en semillas y granos usualmente es producido durante el desgranado y el manejo realizado por máquinas transportadoras como tornillos y elevadores; éste se refleja como la falla o ruptura del producto bajo deformación cuando éstas son forzadas a pasar a través de canales de dimensiones fijas; o bien, cuando son sujetas a cargas de impacto o compresión excesivas (Mohsenin, 1986).
El daño mecánico en semillas repercute de manera directa sobre la calidad física y además puede ocasionar la pérdida o inestabilidad de diversos componentes celulares, lo que acelerará el detrimento fisiológico y sanitario de las mismas. En semillas de algodón (Gossypium), la ruptura de la cubierta seminal aumenta los ácidos grasos que ocasionarán una reducción en el porcentaje de germinación (Mohsenin, 1986). En algunos casos, los tejidos dañados mecánicamente podrían ocasionar un detrimento en la calidad fisiológica, aunque éstos no desempeñen una función metabólica. En este contexto, Cicero et al. (1998) y Carvalho et al. (1999) afirman que la ruptura en el endospermo puede producir una plántula anormal o afectar la calidad de la semilla al restringir la translocación de nutrientes hacia el eje embrionario; asimismo, la ruptura en semillas puede ser causa de una mayor invasión por hongos y otros patógenos. Un estudio realizado en trigo (Triticum spp.) mostró que el daño ocasionado en la capa de aleurona o en la testa permite mayor penetración de hongos (Mohsenin, 1986); por otro lado, en semillas de maíz se encontró que pequeñas grietas realizadas sobre el embrión, o la remoción del pedicelo, ocasionaron un incremento en el desarrollo de hongos y un decremento en la germinación (Tuite et al., 1985).
Diversos estudios resaltan la importancia de la evaluación de la compresión a nivel microscópico; (Mohsenin, 1986) confirmó que la ruptura celular inicial ocasionó discontinuidad en la curva fuerza-deformación y menciona que la mayor parte de las células localizadas a uno y otro lado de la línea de fractura aparentaron no tener daño; es decir, no mostraron ruptura celular; asimismo, dicho autor menciona que, en semillas de maíz, una ruptura en el endospermo puede derivarse de exceder la deformación elástica o inelástica de la capa de aleurona, lo que ocasiona rupturas visibles o invisibles al ojo humano. Mohsenin (1986) menciona que algunos términos usados hasta ese momento no habían sido completamente comprendidos y definidos, como la biocedencia, y sugiere que ésta pudiera definirse como el inicio de la ruptura celular. Los estándares de la ASAE (2005) no definen la biocedencia en función del fenómeno que ocurre en el especimen, sino en función del comportamiento de la curva carga-desplazamiento (o esfuerzo-deformación), donde se presenta un incremento en la deformación sin un incremento en la fuerza aplicada o de reacción. Al respecto, Lucas et al. (2000) mencionan que la fuerza de cedencia de un material es una propiedad verdadera de material, que representa el esfuerzo en el que inicia la deformación permanente en un tejido.
Diversos estudios en biología aseguran que la pared celular es uno de los principales elementos responsables de proporcionar resistencia mecánica a las células vegetales y su composición confiere tenacidad a los órganos vegetales (Lucas et al., 2000). La tenacidad, al menos en madera y acero, determina si ocurrirá una falla enseguida de una grieta rápida (Ashby y Jones, 2009) y puede definirse como la capacidad de un material de absorber energía antes de que ocurra la fractura (Callister, 1995). Además de la pared celular, diversos componentes, como las sustancias de reserva contenidas en el endospermo de las semillas, pueden modificar las propiedades mecánicas de un tejido. En este sentido, García-Lara et al. (2004) afirmaron que la pared celular confiere resistencia al ataque de organismos como los gorgojos, por lo que la semilla queda vulnerable luego de la ruptura o desprendimiento de ésta.
De acuerdo con Mohsenin (1986), aún se requieren estudios que permitan entender la mecánica de la falla de tejidos, entre las que se encuentra la ocasionada por compresión, en cuyo caso se desconoce si ésta ocurre mediante la ruptura o separación celular. Cabe señalar que hasta el momento no se han realizado suficientes estudios recientes que permitan comprender a cabalidad ese fenómeno. El mismo autor menciona que la limitada información está basada en estándares arbitrarios derivados de algunas pruebas subjetivas, más que en medidas físicas bien definidas. En el mismo tenor, Van Zeebroeck et al. (2007) indicaron de manera categórica que la relación entre las propiedades micro y macromecánicas permanecen incompletas; su trabajo corresponde a una revisión bibliográfica enfocada al tema, lo que implica una oportunidad para explorar los fenómenos asociados al daño mecánico en la micro y macroestructura de semillas.
En la presente investigación se estudió la falla de los tejidos de la semilla de maíz al ser sometida a compresión axial y su relación con la separación o ruptura celular, por lo que el objetivo planteado fue determinar la proporción de ruptura y separación celular en compresión de ruptura en semilla de maíz en función de la textura de endospermo y contenido de humedad, así como determinar el efecto de la compresión de ruptura sobre la calidad fisiológica de la semilla. Se desea probar las hipótesis siguientes: a) en compresión de ruptura, la proporción de separación es superior a la proporción de ruptura celular, y b) la proporción de ruptura celular en el endospermo harinoso es mayor que en el cristalino.
MATERIALES Y MÉTODOS
Material genético
Se utilizaron semillas de maíz de variedades con endospermo harinoso (Cacahuacintle), semi-dentado (híbrido HS-2) y cristalino (Palomero). El híbrido HS2 y la variedad Cacahuacintle se produjeron en el año 2014 en el Rancho Santa Rosa, Texcoco, México, y en el Campo Experimental del Campus Montecillo, Colegio de Postgraduados, Texcoco, México, respectivamente. La semilla de maíz Palomero se adquirió en 2014 en una casa comercial.
Niveles de humedad
Se evaluaron dos niveles de humedad, 12 y 20 % (± 0.10 %, base húmeda); para lograr estos porcentajes de humedad, las semillas se humedecieron de 2 a 2.5 % por encima de la humedad deseada con agua destilada y posteriormente fueron secadas a temperatura ambiente hasta obtener el contenido de humedad deseado; posteriormente, las semillas se mantuvieron por no más de 4 h en frascos herméticos hasta el momento de la compresión. Después de la compresión, las semillas fueron deshidratadas bajo condiciones ambientales por 24 h, y luego por 20 a 24 h más en estufa a 30 ºC; finalmente, las semillas fueron expuestas a temperatura ambiente hasta alcanzar el porcentaje de humedad deseada y se almacenaron en frascos cerrados herméticamente hasta su evaluación.
Aplicación de compresión
Las semillas se sometieron a compresión justo hasta el momento en que se detectó la ruptura de tejido. Cada semilla se colocó en posición de descanso, con el embrión hacia arriba entre superficies planas y pulidas (ASAE, 2005), la velocidad de desplazamiento de la cruceta fue de 1.0 mm min-1. El equipo utilizado fue una máquina de ensayo universal INSTRON modelo 3385H (INSTRON®, Norwood, MA, USA) con una célula de carga de 50 kN. El desplazamiento de la cruceta se detuvo en cuanto se observó el cambio de pendiente característico en la curva fuerza-desplazamiento o cuando se detectó ruido ocasionado por la ruptura de la semilla; en este último caso, se utilizó un micrófono en contacto con el soporte de la semilla (plato inferior) conectado a una computadora con una amplificación de sonido de 36 dB y supresión digital de ruidos y eco acústico. El volumen de las bocinas se ajustó a un nivel suficientemente audible y se observó el gráfico de nivel de sonido para evitar descartar sonidos de baja intensidad. Luego de efectuar la compresión, la humedad de la semilla se redujo de acuerdo con la metodología mencionada anteriormente, para evaluarse a los 45 y 60 días posteriores.
Diseño del experimento
Para evaluar el efecto ocasionado por los tratamientos se utilizó un diseño experimental completamente al azar con tres factores de estudio: tipo de endospermo (harinoso, semi-dentado y cristalino), contenido de humedad (12 y 20 %) e intensidad de compresión (ruptura y testigo).
Variables evaluadas
Proporción del área fracturada en la microestructura
Para exponer el área fracturada a la observación en el microscopio, las semillas sometidas a ruptura se abrieron con el auxilio de unas pinzas. Los fragmentos de semilla se montaron sobre portaobjetos de latón adheridos con cinta adhesiva conductiva de carbono, se sometieron a vacío por 10 min y se bombardearon en un ionizador de oro/paladio (Modelo JFC-1100, JEOL Ltd., Tokio, Japón) por un periodo de 5 min; finalmente, se observaron en un microscopio electrónico de barrido (Modelo JSM6390, JEOL Ltd., Tokyo, Japón) operado a 10 kV. Se capturaron imágenes en aumentos de 10, 150, 300, 600 y 1,200X. Para determinar la proporción de ruptura y separación celular en la microestructura del endospermo se analizaron cinco micrografías por cada tratamiento con superficie de muestra equivalente a 1.37 mm2 ; para determinar esas proporciones en la capa de aleurona, se analizaron 21 micrografías con acercamiento de 300 a 1000X. En tejido embrionario y escutelo se tomaron dos y cuatro micrografías, respectivamente.
Conductividad eléctrica
Se realizó de acuerdo con la metodología descrita por la ISTA (2005) para evaluar conductividad eléctrica en chícharo (Pisum sativum), aunque se utilizaron tres repeticiones de 30 semillas. Para determinar la conductividad eléctrica se utilizó un conductímetro (Marca Oakton® Modelo WD-35607, Vernon Hills, Illinois, EUA). Los resultados se expresaron en μS cm-1 g-1.
Germinación estándar
La prueba de germinación se llevó a cabo mediante la metodología entre papel recomendada por la ISTA (2005). Se evaluaron tres repeticiones de 30 semillas para cada tratamiento.
RESULTADOS Y DISCUSIÓN
Proporción del área de fractura en la microestructura
Endospermo
El análisis realizado a la microestructura de endospermo observada en microscopía electrónica de barrido mostró que la fractura tisular en semillas comprimidas estuvo compuesta principalmente por ruptura celular y en menor proporción por separación celular (Cuadro 1). El endospermo, excluyendo la capa de aleurona, está conformado por células que almacenan carbohidratos, lípidos y proteínas (Ritchie et al., 2000); por consiguiente, la ruptura celular puede representar únicamente algunos cambios en los compuestos de almacenamiento, especialmente si tiene intercambio de gases directamente con la atmósfera, pues el oxígeno podría desnaturalizar algunos compuestos como las proteínas y los lípidos al ser expuestos a radicales libres, ocasionando efectos de envejecimiento (Schwember y Bradford, 2011; Smith y Berjak, 1995), además de la exposición de las reservas al ataque de patógenos (Mycock y Berjak, 1995).
Cuadro 1. Comportamiento medio de la proporción de área con ruptura celular y proporción de área con separación celular en células de endospermo, de capa de aleurona, embrión y escutelo de semillas de maíz sometidas a compresión de ruptura.

Medias con letra distinta en cada columna difieren estadísticamente (Tukey, 0.05). † Las células referidas como endospermo para este cuadro excluyen a las células de aleurona. ††Las células de aleurona son células de endospermo nucleadas y metabólicamente activas. ¶ Excluye el escutelo.
Aunque la pared celular brinda integridad y regularmente confiere resistencia mecánica (Lucas et al., 2000), en el caso de las células del endospermo, las paredes son muy delgadas, desde 310 nm de espesor en un par de paredes adyacentes (Figura 5D) hasta 1500 nm para la pared de la capa de aleurona (Figura 6B) y no confieren resistencia en comparación con la pared celular de otros tejidos (Watson, 2003). Es probable que, debido a ello, la proporción de ruptura celular fue mayor que la separación celular. Es común que en estudios realizados donde se provocan fracturas de manera artificial, al observar la microestuctura del endospermo, éste presente una alta proporción de granos de almidón expuestos como lo señalan GaytánMartínez et al. (2006) y Narváez et al. (2007b), quienes muestran imágenes de células del endospermo donde se observan los granos de almidón, lo que apoya los hallazgos del presente estudio.
La humedad tuvo efecto significativo (P ≤ 0.08) sobre la proporción de área de endospermo con pared celular equivalente al área con separación celular y con la complementaria proporción de ruptura celular; el tipo de endospermo no tuvo efecto significativo sobre dicha proporción. Con la humedad de 12 % se tuvo una mayor proporción de área con pared celular, en comparación con 20 % de humedad (Cuadro 2); no obstante, en ambos casos el porcentaje resultó muy bajo (inferior a 20 %), lo que indica que la pared celular se volvió frágil con la hidratación, como sucede en algunos casos (Ziegler, 1995), pero la mayoría de las veces su capacidad de deformación, tenacidad y resistencia se incrementa con la humedad (Mohsenin, 1986).
Cuadro 2. Comportamiento medio de la proporción de área de endospermo con pared celular en semillas de maíz sometidas a compresión de ruptura a humedad de 12 y 20 % (b. h.).

Medias con letra distinta difieren estadísticamente (Tukey, 0.08).
En la Figura 1D, correspondiente a semillas de la variedad con endospermo semi-dentado (HS-2) comprimidas a 12 % de humedad, se observan células con ruptura exponiendo los granos de almidón, el límite de la pared celular y el aspecto de las fracturas celulares. En la Figura 1C se observó un patrón sobre la superficie fracturada que pudiera representar las trayectorias de los esfuerzos principales, el cual está asociado a la forma y orientación de las células de endospermo duro; dicho patrón es casi perpendicular a la dirección en que se aplicó la compresión (Figuras 1A, B y C).

Figura 1. Micrografías electrónicas de barrido de fracturas internas en una semilla de maíz semi-dentado (HS-2) con 12 % de humedad sometida a compresión: A) Posición de la semilla al momento del ensayo; la flecha indica la dirección de desplazamiento del plato de compresión (dpc), s: zona del escutelo, es: endospermo suave, ed: endospermo duro. B) Micrografía electrónica perpendicular (90º ↺) al recuadro en A donde se muestra superficie de la fractura. C) Detalle del recuadro en B (45º ↻), donde se aprecia la orientación de las células periféricas de endospermo, p: pericarpio, ca: capa de aleurona. D) Detalle del recuadro en C, donde se aprecian ga: granos de almidón, lc: límite celular, fc: una fractura celular y pc: porción de la pared celular.
En la Figura 2D, correspondiente a la variedad con endospermo cristalino (Palomero), comprimido a 12 % de humedad, se observan células con pared y células con gránulos de almidón expuesto, ubicadas en áreas de endospermo duro, cerca de la zona central de la semilla. En este último grupo de células, el desprendimiento de pared no fue completo sino parcial en cada célula, como lo observado en las Figuras 1C, 1D y 4D; esto puede deberse al arreglo celular, ya que en estas micrografías las células están alternadas sobre la superficie, mientras que en la Figura 2F las fronteras celulares se ubican sobre el mismo plano.
En las Figuras 2E y F, correspondientes a la variedad con endospermo cristalino (Palomero) comprimido a 12 % de humedad, se aprecian total o parcialmente nueve células separadas, lo cual se deduce por la presencia de los contornos celulares y la ausencia de granos de almidón sobre la superficie observada, así como por la presencia de una cubierta continua que corresponde a la superficie interna o externa de la pared celular.

Figura 2. Micrografías electrónicas de barrido de fracturas internas en una semilla de maíz cristalino (Palomero) con 12 % de humedad sometida a compresión: A) Posición de la semilla al momento del ensayo, el embrión se ubica en la parte superior. B) Micrografía donde se muestra la superficie de la fractura. C) Aumento de B en la zona de interés. D) Recuadro “a” en B, en la zona de endospermo duro, donde se observan ga: granos de almidón y pc: pared celular. E) Recuadro “b” en B, en la zona de endospermo duro, hay ausencia de granos de almidón expuestos, por lo que se asume que la superficie corresponde a pared celular. F) Contornos de las células del área en E.
En la Figura 3B, correspondiente a la variedad con endospermo cristalino (Palomero) comprimido a 12 % de humedad, se observa una envoltura fina o restos de ella alrededor de los amiloplastos, que corresponde a la membrana lipoproteica característica de los fenotipos conocidos como cerosos y palomeros (Bewley y Black, 1978); los amiloplastos de estas superficies están rodeados también por unos gránulos más pequeños (Figura 3D) que pudieran ser también de almidón, proteína u otras sustancias (Bewley y Black, 1978); al respecto, NarváezGonzález et al. (2007a) mencionan que en razas de maíz palomero predominan los gránulos de almidón pequeño (< 12 μm), de forma hexagonal y rodeados de una densa matriz proteica.
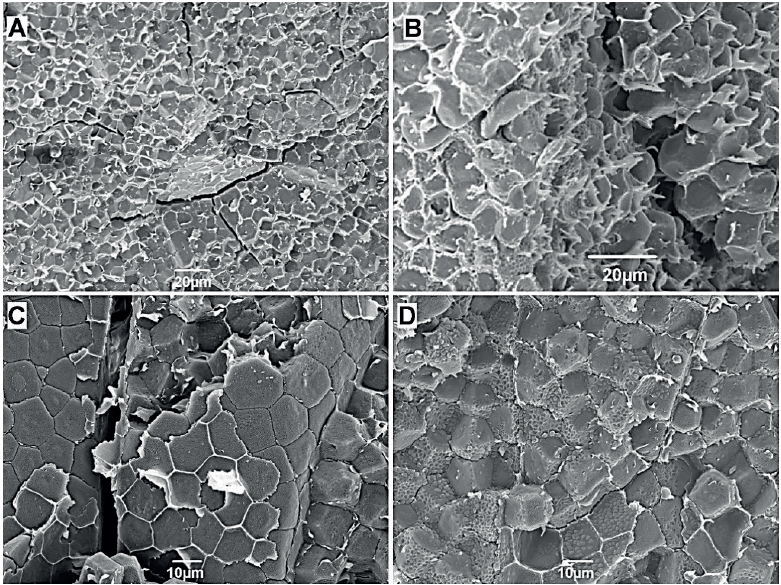
Figura 3. Micrografías electrónicas de barrido de fracturas internas en una semilla de maíz cristalino (Palomero) con 12 (A y B) y 20 % (C y D) de humedad sometidas a compresión: A) Recuadro “c” en la Figura 2B. B) Recuadro “d” en la Figura 2B, nótese la estructura que rodea a los granos de almidón correspondiente a la matriz proteica que le da al endospermo la apariencia compacta típica del endospermo cristalino. C) Células con restos de pared celular y granos de almidón con restos de la matriz de proteína. D) Granos de almidón rodeados de una densa matriz proteica.
En la Figura 4D, correspondiente a la variedad con endospermo harinoso (Cacahuacintle) comprimido a 12 % de humedad, se observan superficies correspondientes a límites celulares (superficie interna o externa de pared celular) y superficies con granos de almidón expuestos. Nótese la ausencia de la matriz proteica (Figura 3D) y de la capa cerosa que envuelve a los gránulos de almidón (Figura 3B) características de las variedades cristalinas. Las células se aprecian intactas, aunque en la mayoría de los casos sin la pared celular.

Figura 4. Micrografías electrónicas de barrido con el detalle de la superficie de fractura en la zona cercana al pericarpio en una semilla de maíz harinoso (Cacahuacintle) con 12 % de humedad sometida a compresión: A) El embrión se ubica en la parte superior, el recuadro muestra el área de fractura provocada durante el ensayo de compresión, el resto se fracturó al separar la semilla. B) Aumento del recuadro en A. C) Aumento de B, donde se aprecian contornos intactos de célula con o sin pared celular. D) Aumento de C, lc: límites celulares o pared celular y ga: granos de almidón.
En la Figura 5B, correspondiente a la variedad con endospermo semi-dentado (HS-2) comprimido a 12 % de humedad; se observan células de almidón suave con una alta frecuencia de ruptura celular, representada por la proporción grande de granos de almidón expuestos (Figura 5C), se aprecian también las paredes celulares cuyo espesor es de 155 nm, 310 nm por cada par de paredes adyacentes (Figura 5D); se observaron también células con endospermo duro en la periferia, donde los granos de almidón están rodeados por una matriz proteica (Figura 5B)

Figura 5. Micrografías electrónicas de barrido de fracturas internas en una semilla de maíz semi-dentado (HS-2) con 12 % de humedad sometida a compresión: A) Vista panorámica. B) Recuadro “a” en A, que muestra el endospermo duro con granos de almidón rodeados de una densa matriz proteica. C) Recuadro “b” en A, que muestra endospermo suave, se observan los límites celulares, los granos de almidón y la ausencia de pared celular sobre esta superficie. D) Endospermo suave que muestra paredes celulares al centro y tres granos de almidón de la célula superior y dos de la inferior
Aleurona, embrión y escutelo
A nivel microestructural, las fracturas de la aleurona presentaron principalmente ruptura celular y en menor proporción separación celular (Cuadro 1). Sobre la superficie de la fractura de semillas del híbrido HS-2 y del Palomero, en células de la capa de aleurona, se observó la presencia de unas fibrillas que corresponden a cristales de carbonato de calcio (Figura 6A). A pesar de que el grosor de la pared celular en células de la capa de aleurona (1.5 μm) fue superior al grosor de pared celular de otras células del endospermo (0.15 μm), la ruptura celular superó a la separación celular. Es probable que las sustancias cementantes confieran una mayor resistencia a la proporcionada por la pared celular, lo que ocasiona que en la ruptura de tejido ocurriera mayor proporción de ruptura que de separación celular, similar a lo que ocurre durante la molienda, donde la ruptura celular en la capa de aleurona es común (Watson, 2003).
La compresión aplicada no ocasionó fractura en tejido embrionario (embrión y escutelo), y aunque la semilla se fracturó artificialmente a partir de la fractura ocasionada sobre tejido de endospermo, no se apreció daño o fractura a nivel tisular ni celular en el tejido embrionario y escutelo (Figuras 6C y D), por lo que no se analizó estadísticamente la proporción de ruptura y separación celular para estos tejidos; en el Cuadro 1 se incluyeron dichos valores (todos de cero) sólo como referencia. Uno de los factores que protege al embrión de las fracturas propagadas desde el endospermo es la capacidad de deformación que permite disipar la energía que ocasiona la fractura, lo que podría atribuirse a su mayor elasticidad (Ashby y Jones, 2009; Callister, 1995; Tipler y Mosca, 2005).

Figura 6. Micrografías electrónicas de barrido de fractura en células de una semilla de maíz semi-dentado (HS-2) a 12 % de humedad (A y C) y harinoso (Cacahuacintle) a 20 % de humedad (B y D): A) p: pericarpio, a: capa de aleurona, cc: cristales de carbonato de calcio. B) Célula de aleurona, n: núcleo y t: punteaduras. C) Eje embrionario, sin presentar daño. D) Superficie del escutelo, que muestra células sin daño.
Calidad fisiológica
La variedad con endospermo cristalino obtuvo el mayor valor de conductividad eléctrica (CE), usualmente asociado a un mayor grado de deterioro. La CE fue inferior con humedad de 20 %, lo cual se atribuye a una pérdida inicial de solutos durante la hidratación previa en semillas comprimidas a 20 % (Bewley y Black, 1994). El valor de CE en semillas comprimidas fue casi el doble del correspondiente al testigo (Figura 7A).
La mayor germinación estándar se obtuvo en la variedad con endospermo semi-dentado (HS-2), seguido del harinoso (Cacahuacintle) sin diferencia significativa en semillas sin comprimir. La germinación fue significativamente superior en semillas con 12 % de humedad y en semillas sin comprimir (Figura 7B); cabe resaltar que la fuerza aplicada a las semillas con humedad de 12 % y 20 % no presentó diferencia significativa, con valores de 312.5 y 313.8 N respectivamente; el efecto final sobre la germinación se atribuyó a que la mayor humedad al momento de recibir la carga de compresión ocasionó una mayor actividad fisiológica y deterioro posterior al ensayo de compresión. El deterioro de la semilla sometida a ruptura se asocia tanto a la reducción en la capacidad de la translocación de nutrientes o señalización, ocasionado por la separación celular, y a la desnaturalización de los componentes celulares, ocasionado por la ruptura celular.

CONCLUSIONES
En la microestructura del endospermo y de la aleurona se observó separación y, predominantemente, ruptura celular. La compresión aplicada no ocasionó daño sobre tejido embrionario, pero el daño ocasionado por fractura en el endospermo estuvo asociado con una menor germinación. La proporción de ruptura celular no fue diferente entre el endospermo harinoso y cristalino, pero sí estuvo influenciada por la humedad de la semilla. El deterioro de la semilla sometida a ruptura se asocia con la reducción en la capacidad de la translocación de nutrientes o señalización, ocasionado por la separación celular, así como con la desnaturalización de los componentes celulares ocasionada por la ruptura celular.

