











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El mejoramiento genético es el arte y la ciencia que permite el desarrollo de cultivares superiores (Hallauer et al., 2010), el cual permite generar genotipos que se adapten a diferentes ambientes y que satisfagan las necesidades de la sociedad y de los productores.
El maíz (Zea mays L.) posee una variabilidad genética enorme, aun en razas previamente consideradas uniformes genéticamente, como Zapalote Chico de la región istmeña de Oaxaca, México (Cabrera-Toledo et al., 2019); sin embargo, el proceso de mejoramiento provoca la reducción severa de la base genética (Fernie et al., 2006); por esto, en la faja maicera de Estados Unidos de América la mayoría de los programas de mejoramiento se enfocaron al uso de líneas o variedades que se originaron a partir del entrecruzamiento de materiales élite ya existente (Goodman, 1999); en el caso de México, se generaron variedades mejoradas a partir de germoplasma de origen templado, tropical y subtropical del interés de los mejoradores en las áreas productoras de maíz, lo cual provocó una pérdida de variabilidad en estas regiones (De La Cruz et al., 2003). Es muy importante el conocimiento sobre la diversidad genética para hacer uso efectivo y eficiente del germoplasma con uso potencial para los programas de mejoramiento (Adebayo et al., 2015).
Las poblaciones nativas de maíz en México son una fuente valiosa de variantes alélicas, porque en el proceso de selección continua y dinámica por los agricultores las variedades de cada raza han adquirido características de adaptación para regiones agrícolas particulares (Prasanna, 2012). En este contexto, una alternativa para ampliar la variabilidad genética del maíz dentro de cada región agrícola es la introducción y uso de germoplasma de otras regiones; con ello, además de aportar genes nuevos y variedades diferentes a las locales, es factible incrementar la heterosis de rendimiento entre cruzas de germoplasma diferente (Mickelson et al., 2001), su tolerancia a plagas y enfermedades y a otros factores adversos (Hallauer y Carena, 2009).
No obstante, emplear germoplasma introducido de maíz en una región no es sencillo porque requiere de un proceso de adaptación a las condiciones ambientales donde será utilizado (Gouesnard et al., 1996). En este sentido, diferentes autores han demostrado que la selección masal recurrente es una metodología eficiente para adaptar un maíz de origen tropical a Valles Altos de México; de esta manera, se esperaría que en ciclos avanzados de selección se pueda explotar las varianzas genéticas aditiva y de dominancia, y que sus poblaciones resultantes puedan ser empleadas per se o en la formación de variedades e híbridos con mayor heterosis en rendimiento y de sus componentes (Gómez-Espejo et al., 2015; Pérez-Colmenarez et al., 2000).
Diferentes trabajos confirman las bondades de usar germoplasma adaptado a una región agrícola, particularmente, de materiales originarios del trópico adaptado a una región templada. Gómez-Espejo et al.(2015) observaron valores de heterosis altos para rendimiento entre maíz Chalqueño y Tepecintle (16 a 58.5 %) y entre la cruza Tepecintle original × Tepecintle adaptado (126 %); asimismo, Carrera-Valtierra y Cervantes-Santana (2002) observaron mayor heterosis al cruzar líneas tropicales de maíz con líneas locales de Valles Altos (90.36 a 164.86 %) comparado con cruzas entre líneas tropicales previamente adaptadas a Valles Altos (17.24 a 40.19 %). De manera similar, Palemón et al. (2012) obtuvieron heterosis de 10.6 % en promedio en cruzas intervarietales de germoplasma tropical adaptado a Valles Altos con material subtropical en la región semi cálida de Guerrero.
En México existen reportes de programas de mejoramiento genético de maíz que han adaptado e incorporado germoplasma exótico para ampliar la base genética de sus materiales e incorporarles genes favorables (Gómez-Espejo et al., 2015); asimismo, se cuenta con 10 razas tropicales y subtropicales de maíz que fueron sometidas a selección masal visual para adaptación en Montecillo, Estado de México, donde prevalecen condiciones de clima templado (Pérez et al., 2002). En este contexto, los objetivos del presente trabajo fueron a) evaluar el rendimiento y sus componentes en poblaciones de maíz tropical y subtropical adaptado por selección a Valles Altos y en sus cruzas interraciales y, b) evaluar la heterosis de cruzas interraciales de maíz tropical y subtropical adaptado por selección a Valles Altos para identificar y seleccionar genotipos como fuente potencial para el mejoramiento genético del maíz en dicha región.
MATERIALES Y MÉTODOS
Material Vegetal
Se evaluaron siete poblaciones de maíz adaptado a Valles Altos mediante 20 ciclos de selección masal visual (Pérez et al., 2002): Pepitilla (P1), Tabloncillo (P2), Comiteco (P3), Celaya (P4), Vandeño (P5), Tepecintle (P6) y Nal-Tel (P8); dos poblaciones con 19 ciclos de selección masal: Tuxpeño (P7) y Zapalote Chico (P9); ocho cruzas formadas entre dos poblaciones adaptadas: F1(P1×P3), F1(P2×P1), F1(P4×P1), F1(P4×P3), F1(P7×P5), F1(P6×P5), F1(P7×P6) y F1(P9×P8); dos cruzas entre dos cruzas poblacionales: F1(P2P1×P4P3) y F1(P5P7×P6P7); una entre tres cruzas poblacionales: F1(P5P7×P6P7×P6P5); tres generaciones F2: F2(P9×P8), F2(P2P1×P4P3) y F2(P5P7×P6P7); dos generaciones F3: F3(P9×P8) y F3(P2P1×P4P3); una generación F4: F4(P9×P8) y cuatro testigos locales de maíz Chalqueño: híbrido Promesa (H-PROMESA), híbrido San Josecito (H-SJOSECITO), Criollo de San Juan Tezontla (CSJTEZONTLA) y Compuesto Xolache (COM.XOLACHE).
Sitio experimental
La evaluación agronómica se realizó en tres ambientes diferentes, dos ambientes en el Campo Experimental del Colegio de Postgraduados, Campus Montecillo, en Montecillo, Texcoco, Estado de México, en las coordenadas 19° 27’ 38 N., 98° 54’ 11” O a una altitud de 2250 m, donde se establecieron dos experimentos en lotes contrastantes en cuanto a características edáficas y de humedad, además de fecha de siembra diferente. El tercer lote de evaluación se ubicó en el Campo Agrícola Experimental San Martín de la Universidad Autónoma Chapingo, Texcoco, Estado de México, en las coordenadas 19° 29’ 05” N, 98° 53’ 11” O, a una altitud de 2250 msnm.
Manejo agronómico
La preparación del terreno en los tres ambientes consistió en un barbecho, un paso de rastra, y posteriormente se realizó el surcado. La siembra se realizó el 17 y 23 de mayo de 2017 para los ambientes de Montecillo y el 25 de mayo para el ambiente de Chapingo. En los tres ambientes se sembró a una densidad de población de 54,166 plantas ha-1. La dosis de fertilización fue 160N-80P-00K, se aplicó todo el P y la mitad del N a la siembra y el resto del N se aplicó al aporque 35 días después de la siembra (dds). Para el control de maleza se aplicó Gesaprim Calibre 90 GDA® (1 kg ha-1) combinado con Primagran®Gold (3 L ha1 ) a los 7 dds; posteriormente, a los 60 dds una mezcla de Gesaprim Calibre 90 GDA® más Gramoxone®.
Diseño experimental
Los genotipos se evaluaron en un diseño experimental de bloques completos al azar con tres repeticiones, donde la unidad experimental estuvo conformada por una parcela de dos surcos de 6 m de largo separados a 0.80 m y matas de dos plantas cada 50 cm, para obtener un total de 52 plantas por parcela. La siembra se hizo manual con pala, depositándose dos semillas por sitio y no se hizo aclareo de plantas.
Caracteres evaluados
Los caracteres evaluados fueron rendimiento de mazorca a humedad constante (REN, t ha-1), índice de prolificidad (IP, número de mazorcas/planta), longitud de mazorca (LM, cm), diámetro de mazorca (DM, cm), número de hileras por mazorca (NHM), número de granos por hilera (NGH) y peso de 200 granos (PDG, g). Los componentes del rendimiento LM, DM, NHM, NGH y PDG se midieron en una muestra de 10 mazorcas representativas en cada parcela. El IP se obtuvo al dividir el número de mazorcas cosechadas en la parcela entre el número total de plantas de la parcela. Para estimar el REN se obtuvo el rendimiento de mazorca por planta (RMP) al dividir el peso seco a humedad constante del total de mazorcas de la parcela entre el número de plantas cosechadas con competencia completa en la parcela; este valor se multiplicó por la densidad de plantas por hectárea (54,166) para obtener el REN (t ha-1).
Análisis estadístico
Para todos los caracteres se realizó análisis de varianza combinado de los tres ambientes, comparación de medias de Tukey (P ≤ 0.05) y un análisis de correlación fenotípica entre los caracteres. Estos análisis se hicieron mediante el paquete estadístico SAS® 9.0 (SAS Institute, 2002). La heterosis, con respecto al progenitor medio en los tres ambientes de forma combinada, se calculó con la fórmula H = [(F1 - PM) / PM] × 100, donde H: heterosis porcentual (%); F1: media fenotípica de la población F1; PM= (Pi + Pj)/2, media fenotípica del progenitor medio; Pi y Pj: media fenotípica del padre i y del padre j (Falconer y Mackay, 1996).
RESULTADOS Y DISCUSIÓN
Análisis de varianza
El análisis de varianza (Cuadro 1) detectó diferencias significativas (P ≤ 0.0001) entre los genotipos (GEN) para todos los caracteres, lo que puede atribuirse en parte a que los materiales genéticos de las cruzas son de origen geográfico diferente y, no obstante su adaptación por selección, presentan variación genética intrínseca entre ellos porque los genotipos adaptados pueden tener expresión fenotípica diferente bajo condiciones ambientales de Valles Altos (Gómez-Espejo et al., 2015). Por otro lado, el haber incluido genotipos productivos localmente adaptados como testigos también contribuyó a la significancia de esta fuente de variación.
El análisis de varianza también detectó significancia (P ≤ 0.0001) entre ambientes para IP, REN, LM, PDG y NGH, lo que indica que al menos un ambiente fue diferente al resto para estas características y esto se atribuye a diferencias ambientales con efectos contrastantes entre sitios; por ejemplo, la variación en la precipitación, además de las características edáficas diferentes donde se establecieron los genotipos, como textura del suelo, salinidad, y pH (López et al., 2017), lo cual provocó diferente retención de humedad y por consiguiente variabilidad en los procesos fisiológicos de las plantas.
La interacción genotipo × ambiente (Cuadro 1) resultó significativa (P ≤ 0.0001) para todos los caracteres, excepto para IP (P ≤ 0.0908) y DM (P ≤ 0.1515). Lo anterior indicó que las diferencias entre los genotipos no fueron consistentes a través de ambientes, excepto para IP y DM; es decir, los genotipos presentaron una respuesta diferente en su comportamiento agronómico a través de los ambientes de prueba debido a la existencia de variabilidad genotípica en las poblaciones evaluadas, que permitió una respuesta diferencial para el efecto de cada ambiente (García et al., 2002). Los coeficientes de variación mostraron un intervalo de 3.54 a 16.45 % (Cuadro 1), esto sugiere que el experimento se realizó bajo un buen manejo agronómico y experimental, lo cual minimizó el error experimental y validó los resultados obtenidos.
Cuadro 1. Cuadrados medios y significancia del análisis de varianza combinado del rendimiento y sus componentes de rendimiento en poblaciones y cruzas interraciales de maíz tropical adaptado a Valles Altos, ciclo primavera-verano 2017.
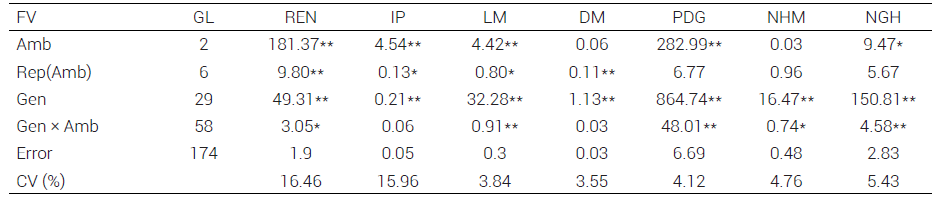
**:P ≤ 0.01
*:P ≤ 0.05, FV:
de variación, GL: grados de libertad, REN: rendimiento de mazorca, IP: índice de prolificidad, LM: longitud de mazorca, DM: diámetro de mazorca, PDG: peso de 200 granos, NHM: número de hilera por mazorca, NGH: número de granos por hilera, CV: coeficiente de variación.
Comparación de medias
La comparación de medias (Cuadro 2) mostró que la población Tuxpeño (P7) (11.71 t ha-1) y el testigo H-PROMESA (11.56 t ha-1) fueron estadísticamente superiores en REN. Este resultado es interesante porque indica que la población de Tuxpeño adaptada por selección masal a condiciones de Valles Altos tiene un rendimiento competitivo con el testigo superior; de acuerdo con Prasanna (2012), la población tropical Tuxpeño seleccionada presenta mayor número de alelos adaptativos, lo que la hace tener un mejor rendimiento per se y también podría generar híbridos como el F1(P7×P6) (10.74 t ha-1), que fue estadísticamente similar a los híbridos comerciales empleados como testigos. Zapalote Chico (P9) (3.89 t ha-1) y Nal-Tel (P8) (4.2 t ha-1) tuvieron rendimientos menores comparado con los demás genotipos, pero presentaron un ciclo vegetativo corto (68 y 73 días respectivamente), lo que representa una ventaja para Valles Altos, donde el periodo libre de heladas es corto, de esta manera permitirían ampliar el ciclo de siembras, inclusive se podría llegar a tener dos por año.
Cuadro 2. Comparación de medias del rendimiento y sus componentes a través de ambientes para las 11 mejores y cinco peores poblaciones de maíz y sus cruzas interraciales adaptadas a Valles Altos, ciclo primavera-verano 2017.
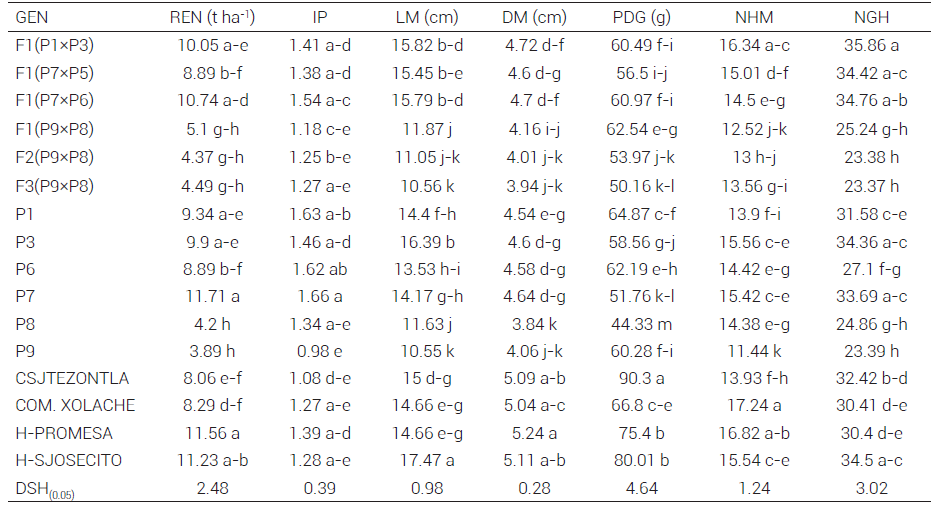
Medias con letras iguales dentro de la misma columna no son estadísticamente diferentes (Tukey, 0.05), GEN: genotipo, IP: índice de prolificidad, REN: rendimiento de mazorca, LM: longitud de mazorca, DM: diámetro de mazorca, PDG: peso de 200 granos, NHM: número de hilera por mazorca, NGH: número de granos por hilera.
Con respecto al índice de prolificidad, la población P7 fue significativamente (P ≤ 0.0001) mayor (1.66 mazorcas/planta) y le siguieron las poblaciones Pepitilla (P1) y Tepecintle (P6) con 1.63 y 1.62 mazorcas/planta, respectivamente. Martínez et al. (1992) indicaron que el índice de prolificidad responde de manera positiva al proceso de selección; de manera similar, Biasutti et al. (2004) y Montenegro et al. (2002) encontraron que a mayor prolificidad el rendimiento potencial de grano incrementa, esto permitió inferir que las poblaciones P7, P1 y P6 tuvieron respuesta positiva a la selección de esta característica.
La LM fluctuó de 10.55 a 17.47 cm. El testigo H-SJOSECITO fue significativamente (P ≤ 0.0001) mejor con respecto a las demás poblaciones estudiadas (17.47 cm), seguido por la población Comiteco (P3) (16.39 cm). Hernández y Esquivel (2004) reportaron valores similares para cruzas de maíz criollo de Valles Altos; en cambio, Carrera y Cervantes (2006) observaron que cruzas dobles de maíz tropical y subtropical presentan mayor longitud de mazorca que los híbridos para Valles Altos. La población P3 podría ser una opción adecuada para mejorar esta característica porque presentó 16.4 cm para LM, el cual es superior estadísticamente comparado con el resto de las poblaciones adaptadas.
Para PDG el genotipo local criollo CSJTEZONTLA de la raza Chalqueño fue estadísticamente (P ≤ 0.0001) superior (90.3 g) con respecto al resto de las poblaciones, lo cual concuerda con lo reportado por Antonio et al. (2004), quienes encontraron que variedades de maíz criollo de la raza Chalqueño fueron superiores al híbrido comercial H-139 porque presentó un PDG de 102.6 g. En contraste, en el presente estudio las poblaciones tropicales y subtropicales adaptadas y sus cruzas fueron las de menor peso de grano en comparación con los testigos; esto se podría explicar por lo reportado por Cázares-Sánchez et al. (2015), quienes indicaron que para maíz tropical el porcentaje de almidón en el grano fluctuó de 57.54 a 69.92 %, lo cual genera menor peso en comparación con maíces originarios de regiones templadas como el Chalqueño. En relación con el NHM, el testigo COM.XOLACHE fue estadísticamente (P ≤ 0.0001) superior a los demás genotipos con 17.24 hileras (P ≤ 0.05), lo que concuerda con lo reportado por Wellhausen et al. (1952), quienes observaron que la raza Chalqueño presenta mayor NHM comparado con los maíces tropicales y subtropicales.
Por otro lado, la F1 de la cruza simple Pepitilla × Comiteco (P1 × P3) fue significativamente mejor para NGH con 35.86, el cual es superior al valor promedio de diferentes híbridos experimentales de maíz para Valles Altos observado por Espinosa-Calderón et al. (2012). En cuanto al DM, H-PROMESA presentó 5.24 cm y fue superior estadísticamente (P ≤ 0.0001) a las demás poblaciones; en contraste, el material Nal-Tel (P8) fue el de menor DM (3.84 cm). Estos resultados coincidieron con los obtenidos por Martínez-Lázaro et al. (2006), quienes observaron que el híbrido Promesa presentó 0.5 cm más que los híbridos de maíz de cruza simple de Valles Altos para DM.
Los resultados anteriores indican que la selección masal aplicada por varios ciclos de selección a poblaciones de maíz tropicales introducidas en Valles Altos propició una adaptación adecuada de dichos materiales a dicha región, medida con base en el rendimiento y sus componentes de las poblaciones evaluadas (Gómez-Espejo et al., 2015). En la presente investigación los resultados comprueban que la generación de mayor diversidad fue de gran importancia porque se podría ampliar la base genética de las variedades locales para mejorar el rendimiento de maíz de Valles Altos (Badu-Apraku et al., 2015; Edmeades et al., 2017).
Heterosis
La heterosis promedio para todos los caracteres evaluados con respecto al progenitor medio, en los tres ambientes, fluctuó de -5.30 a 6.53 % (Cuadro 3) y resultó positiva para rendimiento (5.68 %) con valores individuales de 4.3 a 26.19 %, ésto significó un aumento promedio de 0.48 t ha-1 en el rendimiento de las cruzas, como fue el caso de la F1(P9×P8) que presentó la mayor heterosis (26.19 %) y REN de 5.1 t ha-1, superior a la media de Valles Altos de 2.79 t ha-1 (SIAP, 2019). Estos valores se asemejan a los reportados por Escorcia-Gutiérrez et al. (2010) y Palemón et al. (2012). La heterosis alta de la cruza F1(P9×P8) se explica por los bajos rendimientos que presentaron los progenitores, como lo sugirieron De la Cruz-Lázaro et al. (2010).
Cuadro 3. Heterosis (%) estimada en 11 cruzas de maíz de origen tropical y subtropical adaptado a Valles Altos en tres ambientes de evaluación, ciclo primavera-verano 2017.
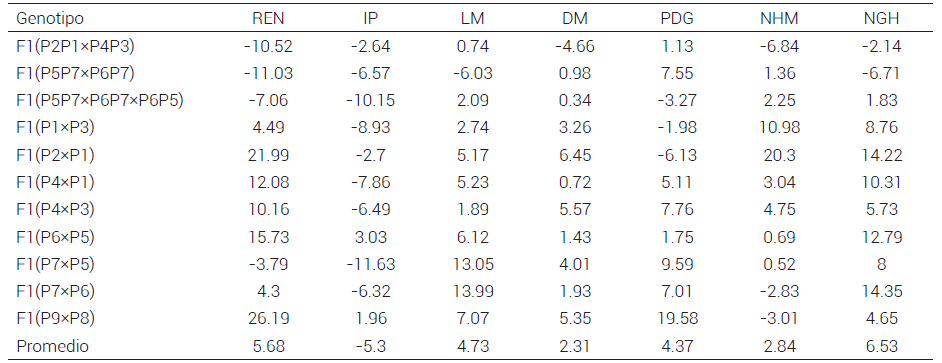
IPíndice de prolificidad, REN: rendimiento de mazorca, LM: longitud de mazorca, DM: diámetro de mazorca, PDG: peso de 200 granos, NHM: número de hilera por mazorca, NGH: número de granos por hilera.
Para el índice de prolificidad (IP), sólo las cruzas F1(P6×P5) y F1(P9×P8) presentaron heterosis positiva individual (3.0 % y 2.0 %); en cambio, la heterosis promedio del carácter fue negativa (-5.3 %), lo que indica que el IP no exhibe heterosis, aunque en la mayoría de las cruzas simples esta aparente pérdida de prolificidad de mazorca por planta se compensó con heterosis positivas para el rendimiento de mazorca y sus componentes.
El carácter LM presentó una heterosis promedio de 4.73 %, con una variación de -6.03 % a 13.99 %; la F1(P5P7×P6P7) presentó heterosis negativa para este carácter (-6.03 %); en cambio, las cruzas F1(P7×P5) (13.05 %) y F1(P7×P6) (13.99 %) fueron las de mayor heterosis. Este resultado pudo deberse a que en la cruza participa P7 como progenitor en común en ambas cruzas poblacionales, lo que generó loci comunes para longitud de mazorca, y por lo tanto, una menor divergencia genética y expresión del carácter (De la Cruz et al., 2003).
Para DM se obtuvo una heterosis promedio de 2.31 %. La cruza que presentó mayor heterosis fue F1 (P9×P8) con 5.35 %. Estos resultados fueron similares los reportados por Amiruzzaman et al. (2010), quienes observaron heterosis de -12.16 a 8.78 % para DM en cruzas de líneas de maíces de origen tropical y subtropical.
El NGH y peso de grano son componentes de rendimiento importantes, por lo que se busca que la heterosis para estos caracteres sea positiva. El carácter NGH presentó una fluctuación de -6.71 a 14.35 %, donde la cruza F1(P7×P6) presentó la mayor heterosis (14.35 %), ésto significó que la combinación de Tuxpeño con Tepecintle mostró diversidad para esta característica. Así, De la Cruz et al. (2003) obtuvieron heterosis de 1 a 24 % para NGH en cruzas de maíz de poblaciones exóticas y adaptadas × exóticas; por lo anterior, se puede inferir que las poblaciones de este trabajo adaptadas a Valles Altos presentaron divergencia genética para NGH, lo que favoreció al rendimiento.
La heterosis de PDG fluctuó entre -6.13 y 19.58 %. La cruza con mayor heterosis fue F1(P9×P8) (19.58 %), ésto indicó que, aunque los progenitores son de semilla pequeña y de rendimiento menor, poseen características que impactan positivamente el rendimiento de grano cuando se cruzan y podrían ser empleadas para el mejoramiento genético. Amiruzzaman et al. (2010) observaron resultados similares al evaluar cruzas entre líneas de maíz divergente.
El carácter NHM tuvo una heterosis de -6.84 a 20.30 %, donde la cruza F1 (P2×P1) presentó la mayor heterosis. Esto contrastó con lo reportado por Malik et al. (2004), quienes identificaron heterosis de hasta 49.6 % con respecto al progenitor medio en líneas de origen templado, subtropical y tropical.
Los resultados obtenidos demostraron que las poblaciones adaptadas tienen gran potencial para ser empleadas per se, o bien como progenitores en cruzas simples; por ejemplo, las cruzas F1(P2×P1), F1(P6×P5), F1(P7×P6) y F1(P9×P8) mostraron el mayor incremento para rendimiento y sus componentes; ésto también permitiría ampliar la variabilidad morfológica de poblaciones de Valles Altos.
Correlación
El análisis de correlación fenotípica mostró correlación positiva entre la mayoría de los caracteres evaluados, excepto en el carácter IP. En el caso del REN, los componentes que correlacionaron de manera significativa (P ≤ 0.0001) y positiva fueron IP (r = 0.67), LM (r = 0.64), DM (r = 0.65) y NGH (r = 0.64), lo cual indicó que los genotipos con mayor índice de prolificidad, mayor longitud y diámetro de mazorca, y más NGH presentaron rendimiento superior tales como P7, H-PROMESA y H-SJOSECITO. Al respecto, diferentes autores observaron correlación positiva entre REN, NGH, DM y LM (Rafiq et al., 2010; Rani et al., 2017), por lo que estos componentes son características que deben tomarse en cuenta en el mejoramiento del maíz para Valles Altos.
CONCLUSIONES
Las poblaciones de maíz tropical y subtropical adaptadas a Valles Altos presentaron características sobresalientes para rendimiento y sus componentes en las condiciones agronómicas de Valles Altos, por lo que podrían ser empleadas en el mejoramiento genético de maíz, especialmente aquella de la raza Tuxpeño. Debido a su ciclo vegetativo precoz, la cruza de las poblaciones adaptadas de Zapalote Chico y Nal-Tel serían una alternativa de producción para regiones que presentan un ciclo agrícola reducido o para aquellas sin restricciones de humedad, porque se podría obtener más de un ciclo de siembra por año. El rendimiento correlacionó de manera positiva con índice de prolificidad, longitud de mazorca, diámetro de mazorca y número de granos por hilera; por ésto, se debe considerar tales componentes en la selección para rendimiento del maíz. Las cruzas interraciales de maíz presentaron heterosis positiva en la mayoría de caracteres evaluados, sus progenitores podrían ser una opción viable para emplearse en el mejoramiento genético de componentes de rendimiento de variedades locales de Valles Altos.

