











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La familia Caricaceae consta de seis géneros y 35 especies (Fuentes y Santamaría, 2014); de éstas, Carica papaya L. es la especie más importante por el valor comercial y volumen de ventas del fruto Carica papaya, es una especie originaria del norte de Centroamérica y sur de México, donde existen poblaciones cultivadas y silvestres (Fuentes y Santamaría, 2014; Manshardt y Zee, 1994); muestra alta variación morfológica en características como forma, tamaño, color de epidermis, sabor, sólidos solubles totales en el fruto y porte de la planta (Singh y Kumar, 2010). La caracterización morfo-agronómica es un método tradicional y eficiente que se utiliza ampliamente para conocer la diversidad morfológica de las poblaciones (Castellen et al., 2007).
Debido al predominio actual de variedades mejoradas, las poblaciones nativas cultivadas como Cera Amarilla, Mamey, Zapote y Coco han dejado de tener importancia comercial, por lo que su tamaño poblacional se ha reducido, encontrándose solamente en huertos familiares tradicionales (Ávila et al., 2013, Com. Pers.)1, donde se utilizan básicamente para autoconsumo (Caballero, 1992) y los pobladores conservan el germoplasma, que es un reservorio de genes valiosos para los fitomejoradores.
Existen estudios que se han realizado para caracterizar la variabilidad de C. papaya en variedades tradicionales locales (Aikpokpodion, 2012; Asudi et al., 2010), variedades mejoradas (Singh y Kumar 2010) y ambas (Sompak et al., 2014), las cuales son de interés comercial. Coppens d’Eeckenbrugge et al. (2007), además de las variedades anteriores, incluyeron poblaciones nativas. Aunque dichos estudios contemplan un número reducido de poblaciones, éstos han contribuido a la planificación de estrategias de manejo, mejoramiento genético, conservación en los bancos de germoplasma y utilización sostenible de la diversidad (Ara et al., 2016).
Existe investigación que involucra a poblaciones de C. papaya nativas de México; ésta se ha realizado predominantemente en el sur de los estados de Yucatán y Baja California Sur. Al respecto, Fuentes y Santamaría (2014) reportaron que las plantas femeninas de las poblaciones silvestres de Yucatán miden 5 m de altura y producen en promedio 70 frutos con peso de 20 a 35 g. Soriano-Melgar et al. (2016), al evaluar 81 recolectas de papayos domesticados de Baja California Sur encontraron frutos de hasta 1.6 kg. Lo anterior evidencia que la diversidad morfológica in situ de poblaciones de papayo nativas de México está poco explorada y caracterizada en regiones específicas; por ello, existe la necesidad de estudiar la variación existente de C. papaya nativa en diferentes zonas agroecológicas de México.
Las poblaciones nativas de C. papaya de valor comercial que aún se conservan en México y se han adaptado a condiciones edafoclimáticas diversas, son de interés para ciertos agricultores, y las características fenotípicas de sus frutos son las de mayor importancia para los mercados consumidores (Wall y Tripathi, 2014). En México, la riqueza genética nativa se está aprovechando para generar variedades nuevas de papayo con alta productividad y plantas de porte bajo (Vázquez et al., 2014). El conocimiento de la variabilidad genética en papaya es imprescindible en el estudio de la estructura genética de las poblaciones nativas para proponer estrategias de conservación y aprovechamiento (Allendorf et al., 2012). En este contexto, el objetivo del presente estudio fue caracterizar in situ la variación morfológica, con énfasis en frutos, de 222 ejemplares de C. papaya nativos de México.
MATERIALES Y MÉTODOS
Área de estudio y material biológico
Durante el periodo de 2009 a 2013 se recolectaron frutos de 222 ejemplares de papayo nativos de 10 estados de las vertientes del Pacífico (Baja California Sur, Guerrero, Oaxaca y Chiapas) y del Golfo de México (Hidalgo, San Luis Potosí, Tamaulipas, Veracruz, Tabasco y Campeche). De cada planta se cosecharon de tres a seis frutos en madurez fisiológica. Al momento de la recolecta de frutos se registró la presencia de ramificación en la planta. Los frutos se guardaron en bolsas de papel, se etiquetaron con los datos de pasaporte correspondientes y se utilizaron para la caracterización.
Variables morfológicas
Las variables cuantitativas que se midieron en el fruto fueron longitud (cm), diámetro (cm), relación longitud/diámetro, grosor de pulpa (cm), diámetro de la cavidad central (cm) y volumen comestible (cm3), mientras que las variables cualitativas incluyeron forma del fruto, forma del extremo peduncular, forma del extremo distal, prominencia de aristas, forma de la cavidad central y color de la pulpa. Adicionalmente se registró la ramificación de la planta en campo. Las variables se evaluaron de acuerdo con los descriptores propuestos por la UPOV (2008) para C. papaya, excepto el volumen comestible, propuesto por Aikpokpodion (2012), la forma del fruto propuesta en el catálogo de Dantas et al. (2000) y el color de pulpa de acuerdo con las tablas de colores de Munsell Color (Firm) (1977).
Análisis estadístico
Con los datos de las variables se realizó un análisis de varianza mediante el procedimiento GLM de SAS V.9.1 (SAS Institute, 2004). Posteriormente, se eliminaron variables de pares con coeficiente de correlación menor a 0.5, y con bajo aporte a la variación; con las variables restantes, se obtuvieron los valores y vectores propios mediante la matriz de correlaciones (Rawlings et al., 1988), y con la gráfica de Gabriel (1971) se visualizaron las correlaciones entre variables y su aporte a la variación global (Pla, 1986).
Con las variables seleccionadas se realizó un análisis de componentes principales (ACP), construido con la matriz de correlaciones, la cual se analizó con el método no paramétrico de agrupación Modeclus para estimar el número adecuado de agrupaciones y éstas distribuirlas en una gráfica biplot (Gabriel, 1971). Para las agrupaciones se efectuó un análisis de conglomerados con datos estandarizados, usando distancias euclidianas y el método de ligamiento promedio (UPGMA). El punto de corte de este análisis se definió con el estadístico de Pseudo F. Se aplicó una prueba de medias de Tukey (P ≤ 0.05) sobre los grupos de C. papaya obtenidos con el dendrograma con el paquete estadístico SAS V.9.1 (SAS Institute, 2004).
RESULTADOS Y DISCUSIÓN
Variación morfológica
El análisis de varianza detectó diferencias significativas (P ≤ 0.01) en todas las variables morfológicas entre los ejemplares evaluados (datos no mostrados), lo que demuestra la existencia de variación entre ellos. La ramificación de planta y siete de las características del fruto presentaron CV menor de 20 %, lo que sugiere poca variación aleatoria de estas características. Da Silva et al. (2008) señalaron que un CV con valores de 20 a 40 % y mayores de 40 % se consideran altos y muy altos, respectivamente. En el presente estudio, la forma de la cavidad central, diámetro de la cavidad central y prominencia de aristas presentaron CV altos (26.69, 20.35 y 21.55 %, respectivamente). Para el volumen comestible y forma del fruto, los CV fueron muy altos (66.59 y 50.02, respectivamente) y se deben a la variación amplia de estas dos variables entre los ejemplares por factores aleatorios (Figura 1). Dentro de las características consideradas como de mayor variación, el diámetro de la cavidad central y el volumen comestible coinciden con lo que reportaron Aikpokpodion (2012) y Oliveira et al. (2010), quienes señalaron alta variabilidad entre genotipos de papayo. Ara et al. (2016) indicaron que los valores de CV altos se deben a la variación inter-poblacional, en tanto que Oliveira et al. (2010) los atribuyen a la variación intra-poblacional.

Estructuración de la variación
Los tres primeros compontes principales (CP) explicaron el 77.4 % de la variación total entre los ejemplares estudiados (Cuadro 1). Las características asociadas al CP1 fueron longitud, diámetro, grosor de pulpa, volumen comestible y diámetro de la cavidad central del fruto, mientras que el extremo peduncular y la forma del fruto se relacionaron con el CP2. En el CP3, sólo la prominencia de aristas del fruto aportó significativamente a la variación (Cuadro 1). En esta investigación las características anteriores contribuyeron mayormente a la variación de C. papaya nativa de México y coinciden con las registradas en estudios similares, excepto la variable prominencia de aristas del fruto (Aikpokpodion, 2012; Asudi et al., 2010; Oliveira et al., 2012).
Cuadro 1 Vectores propios, valores propios y proporción de la variación explicada de los tres primeros CP de 10 características morfológicas de C. papaya nativa de México.
| Característica | CP1 | CP2 | CP3 |
|---|---|---|---|
| Longitud del fruto (cm) | 0.424 | 0.034 | -0.199 |
| Diámetro del fruto (cm) | 0.405 | -0.228 | 0.055 |
| Grosor de pulpa (cm) | 0.395 | -0.014 | -0.030 |
| Volumen comestible del fruto (cm3) | 0.390 | -0.040 | -0.253 |
| Diámetro de la cavidad central (cm) | 0.326 | -0.363 | 0.196 |
| Relación longitud/diámetro | 0.284 | 0.416 | -0.284 |
| Forma del fruto | 0.239 | 0.471 | -0.034 |
| Forma del extremo peduncular | -0.117 | 0.533 | 0.306 |
| Prominencia de aristas | 0.208 | -0.115 | 0.724 |
| Forma de la cavidad central | 0.211 | 0.345 | 0.390 |
| Valor propio | 5.1 | 1.7 | 0.9 |
| Variación explicada (%) | 51.3 | 16.8 | 9.3 |
| Variación acumulada (%) | 51.3 | 68.1 | 77.4 |
Con base en las variables cuantitativas del fruto, los ejemplares se dispersaron gradualmente en el CP1 (Figura 2). Manshardt (2014) señaló que el tamaño, la cavidad ovárica y el color de pulpa han sido las estructuras más favorecidas durante el proceso de domesticación. Así, al cambiar los hábitos de consumo, se fueron prefiriendo los frutos con peso superior a 1 kg y paralelamente se promovió el hermafroditismo. Este fenómeno se confirmó durante las recolectas del germoplasma de papayo nativo, al observar poblaciones trioicas como las del tipo Mamey (recolecta 87 de Guerrero) con frutos de 43 cm de longitud y volumen comestible de 5,660.1 cm3.
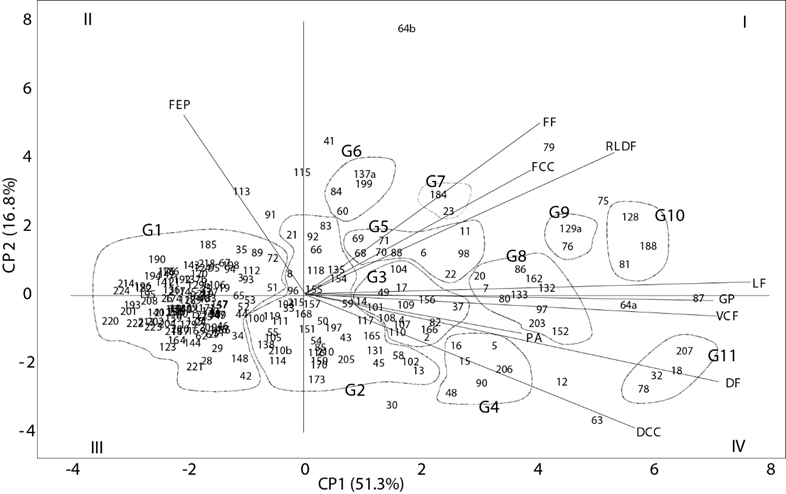
Figura 2 Biplot de 222 ejemplares de Carica papaya elaborado con los dos primeros CP de 10 características morfológicas. Para los acrónimos de las características ver Cuadro 2.
De acuerdo con el análisis de agrupación Modeclus, a un radio en la hiper-esfera de exploración R = 0.7 en el plano de los dos primeros componentes principales, los ejemplares se dispersaron y formaron 11 grupos, distintos fenotípicamente entre ellos.
El Grupo 1 integró 109 ejemplares silvestres que presentaron frutos de dimensiones pequeñas en longitud, diámetro, relación longitud/diámetro, grosor de pulpa, diámetro de cavidad central y volumen comestible de 6.7 cm, 4.6 cm, 1.5 cm, 0.7 cm, 3.4 cm y 31.3 cm3, respectivamente. Cualitativamente, los frutos mostraron forma redonda, cavidad ovárica redonda y aristas pronunciadas ligeramente; además, estos ejemplares silvestres pertenecen exclusivamente a poblaciones dioicas; es decir, las plantas hermafroditas están ausentes, lo cual también han documentado Chávez-Pesqueira y Núñez-Farfán (2016) en México. De las 109 recolectas del Grupo 1,101 proceden de la vertiente del Golfo de México y ocho del Pacífico.
Todos los ejemplares presentaron frutos con dimensiones similares; sin embargo, el CP2 ubicó a los del Pacífico en el Cuadrante II porque presentaron variación en la forma del fruto, la cual fue redonda, oval y elíptica. Las ocho recolectas del Pacífico evidencian que las poblaciones silvestres se encuentran raramente en esta vertiente, ya que la mayoría presenta frutos de tamaño intermedio a grande (Figura 2); esto indica, probablemente, que las barreras montañosas de la Sierra de Juárez y Sierra Madre del Sur han evitado la dispersión de poblaciones externas a ese lugar (Chávez-Pesqueira y Núñez-Farfán, 2016). La pérdida de este germoplasma silvestre también se debe a la destrucción de su hábitat para fines agropecuarios, como se observó directamente durante la recolección de los frutos. La escasez de este material también podría asociarse a una erosión genética baja de la especie (IBPGR, 1986). El Grupo 1 no concentró ejemplares de papayo de Baja California Sur; ésto podría atribuirse a que, en dicho estado, las poblaciones existentes fueron introducidas, pues no se tienen registros de poblaciones silvestres (Badillo, 1971; Soriano-Melgar et al., 2016).
En la parte central del plano (Figura 2) se ubicó el Grupo 2 con 46 ejemplares. Los frutos mostraron dimensiones intermedias en las variables cuantitativas; los valores promedio fueron 13.9 cm de longitud, 8.0 cm de diámetro, 1.8 en la relación longitud/diámetro, 1.5 cm de grosor de pulpa, 5.1 cm de diámetro de cavidad central y 293.7 cm3 de volumen comestible. Los frutos tuvieron forma elíptica y cavidad ovárica angular. Este grupo incluyó papayos tanto de tipos silvestres como de domesticados presentes en huertos familiares, con excepción de las recolectas 92 de Guerrero, 105 y 119 de Oaxaca, que se localizaron en hábitat silvestre. Resalta también la recolecta 13 de Baja California Sur que tiene similitud fenotípica con las silvestres de huertos familiares del Grupo 2, aunque ésta es un genotipo domesticado.
El Grupo 3 concentró 15 recolectas de papayo que presentaron frutos con longitud de 19.9 cm, diámetro de 9.8 cm, volumen comestible de 694.1 cm3, de forma elíptica y extremo peduncular hundido. El Grupo 4 integró seis ejemplares con una longitud promedio de 24.6 cm, diámetro de 13.3 cm, volumen comestible de 1203.5 cm3, forma globular y extremo peduncular hundido en el fruto. Se podría inferir que los ejemplares de ambos grupos son domesticados, ya que se observaron plantas hermafroditas durante la recolecta, además de diferentes colores de pulpa como el rojo, donde destaca la recolecta 206 de Campeche (tipo Mamey). Todos los ejemplares se localizaron en huertos familiares.
El Grupo 5 presentó nueve recolectas de C. papaya. El tamaño y volumen comestible del fruto fueron muy similares a los del Grupo 4, pero se diferenciaron por la forma de pera y extremo peduncular achatada del fruto. Dentro de este grupo se encontró la recolecta 22 de Veracruz (tipo Mamey). El Grupo 6 reunió cuatro recolectas; las dimensiones y volumen comestible del fruto fueron similares a las del Grupo 4, distinguiéndose de éste por la forma de pera y extremo peduncular puntiagudo. Los ejemplares de ambos grupos, así como los del 3 y 4 podrían ser domesticados, debido a que se asociaron con la presencia de plantas hermafroditas y varios colores de pulpa como el rojo (Manshardt, 2014).
El Grupo 7 concentró dos ejemplares de huertos familiares; los frutos mostraron similitud en dimensiones y volumen comestible con aquellos del Grupo 3, pero diferente por ostentar frutos en forma de clava y extremo peduncular redondo. La recolecta 23 de Veracruz (Mamey) destacó por ser hermafrodita con frutos de pulpa roja, característica que permite su comercialización regional.
En la parte final del plano del CP1 se posicionaron el Grupo 8 con 10 recolectas, Grupo 9 con dos, Grupo 10 con tres y el Grupo 11 con cuatro. Los frutos presentaron mayores dimensiones en longitud, diámetro, grosor de pulpa, diámetro de cavidad central y volumen comestible que en el resto de los grupos, incluyendo las variedades Zapote, Mamey y Cera Amarilla, que presentan variación en sexos de la planta, formas y colores de pulpa del fruto. En estos grupos se posicionaron aquellas recolectas con un grado de domesticación más avanzado; además, se observaron plantas hermafroditas. Los frutos se recolectaron en huertos familiares y se comercializan local o regionalmente, sobre todo los de Cera Amarilla, Mamey y Zapote que dominaron el mercado nacional hasta el año 1990 (Soriano-Melgar et al., 2016) como consecuencia de la introducción de la variedad Maradol a México en 1978.
Los ejemplares de los Grupos 3 al 11 que se encontraron en huertos familiares se distribuyeron ampliamente en los Cuadrantes I y IV (Figura 2), ya que mostraron gran dispersión morfológica entre sí. Estas recolectas ubicadas en esos cuadrantes podrían considerarse como germoplasma del complejo papayo de acuerdo con sus dimensiones, formas del fruto y procedencias diversas. Posiblemente dicho germoplasma nativo ha tenido un manejo de selección mínimo; por tanto, la variación que existe pudiera deberse a los distintos usos locales que se remontan desde la época prehispánica (Manshardt y Zee, 1994) hasta la actualidad (Jiménez, 2011).
La forma del fruto también fue determinante en la discriminación entre recolectas. Segun Nakasone y Paull (1998), los agricultores seleccionan esta característica conforme a las preferencias de los mercados consumidores de papaya. Moore (2014) afirmó que los tipos sexuales están determinados genéticamente, aunque el ambiente también modifica la expresión sexual y al tipo de flor que originará formas de fruto diferentes. Plantas con flores femeninas producen formas esféricas a ovoides, y las hermafroditas producen frutos de forma piriforme o alargada con ciertas variaciones, dependiendo de la variedad y de otros factores (Nakasone y Paull, 1998). Los ejemplares de los Grupos 4, 5, 6, 7, 8, 9, 10 y 11 tuvieron valores de relación longitud/diámetro del fruto superiores a 2.0; este valor está asociado con frutos alargados (Chan y Ooi, 1975).
La variación morfológica amplia detectada en este estudio resalta la importancia de conservar in situ o ex situ germoplasma de papayo nativo. También se evidenció que el proceso de selección y mejoramiento genético consciente o inconsciente por los pobladores de las diferentes regiones ha cambiado la estructura genética de las poblaciones debido a características fenotípicas de interés centradas en el fruto; por tanto, los propietarios de los huertos deben conservar este germoplasma, pero con medidas de manejo a corto plazo y de esta manera contribuir a la reducción de los cambios genéticos desfavorables a largo plazo (Allendorf et al., 2012).
La amplia diversidad morfológica se debe tanto a la variación inter como a la intra poblacional; además, se asocia con la polinización cruzada, recombinación meiótica y mutación, seguida por una selección intensiva de genotipos en comunidades rurales aisladas y en constante intercambio de material genético (Asudi et al., 2010). El papayo es un cultivo de polinización cruzada en el que el hábito de la floración y fructificación varía, lo que genera una gran diversidad en forma, tamaño, calidad, sabor y color del fruto (Singh y Kumar, 2010). La polinización cruzada ha causado cambios morfológicos en cada población, lo que causa pérdida de identidad en las variedades de papayo y posiblemente la formación de nuevos genotipos (Oliveira et al., 2012).
Los resultados de este estudio apoyan la teoría de que el sur de México es centro de origen de C. papaya (Badillo, 1971; Fuentes y Santamaría, 2014) por la amplia diversidad encontrada, y probablemente centro de domesticación (Fuentes y Santamaría, 2014; Manshardt y Zee, 1994).
Relaciones entre grupos de papayos
Los agrupamientos de los ejemplares de C. papaya presentaron continuidad en el análisis de conglomerados, principalmente por dimensiones y formas del fruto. Se definieron, de acuerdo con la Pseudo F, tres grupos fenotípicos a una distancia euclidiana de 0.95 unidades (Figura 3). Todas las características presentaron diferencias significativas (P ≤ 0.01) entre grupos (Cuadro 2).
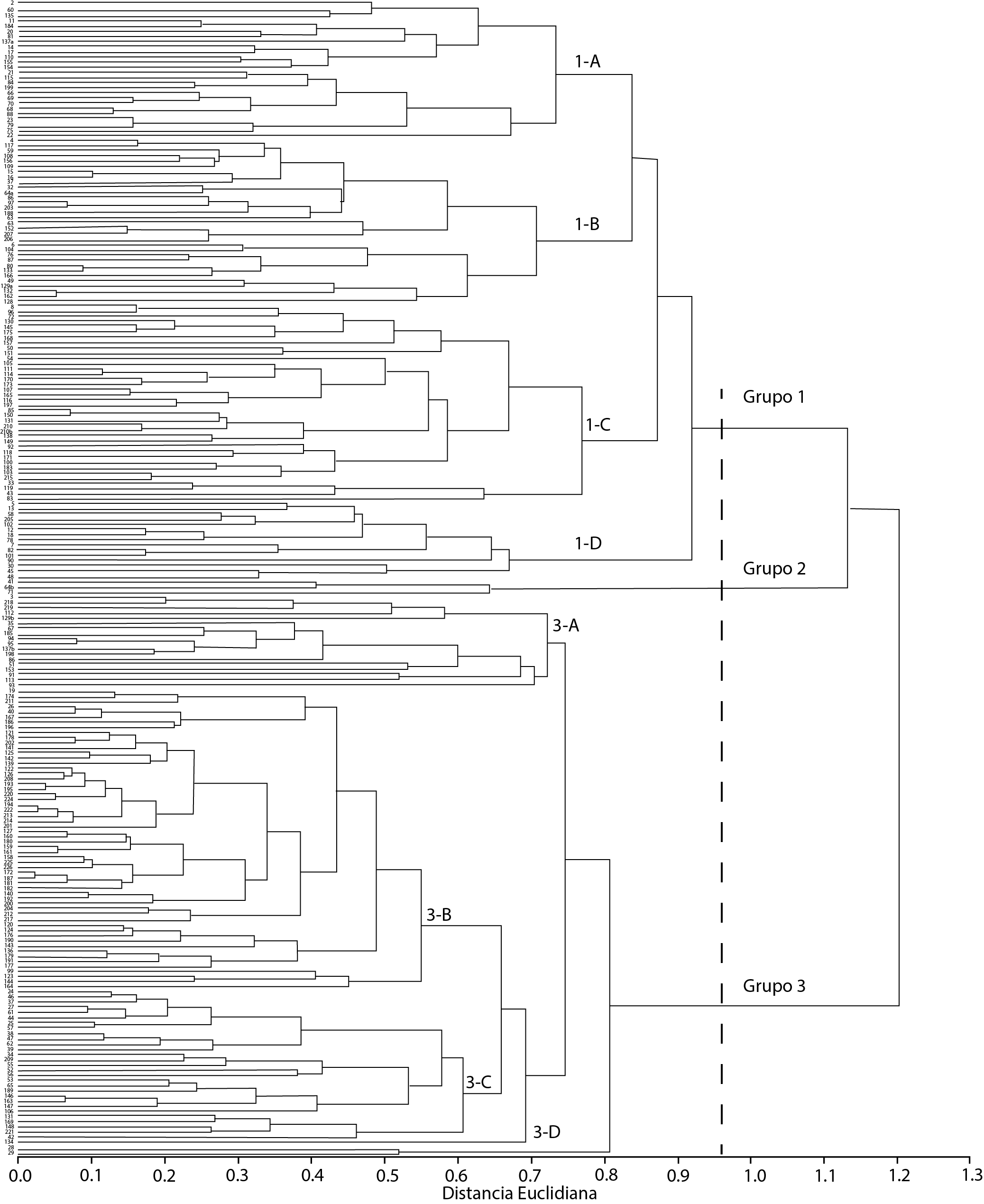
Figura 3 Dendrograma de 222 ejemplares de C. papaya generado con distancias euclidianas con base en 10 características morfológicas y el método de agrupamiento UPGMA.
Cuadro 2 Medias de las características morfológicas del fruto de C. papaya nativa de México, en grupos formados por el dendrograma.
| Grupo | Número de ejemplares | LF (cm) | DF (cm) | RLDF | GP (cm) | DCC (cm) | VCF (cm3) | FF | FEP | PA | FCC |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 111 | 21.0 a | 9.8 a | 2.1 b | 2.1 b | 5.8 a | 1034.6 a | 8.1 b | 1.5 b | 1.6 ab | 2.2 b |
| 2 | 3 | 10.5 b | 4.0 b | 3.4 a | 3.2 a | 2.0 b | 77.2 b | 14.7 a | 3.7 a | 2.0 a | 3.3 a |
| 3 | 108 | 6.6 b | 4.5 b | 1.5 c | 0.7 c | 3.4 b | 30.5 b | 3.5 b | 1.9 b | 1.1 b | 1.3 c |
Medias con letras iguales en cada columna no son estadísticamente diferentes (Tukey, 0.01). LF: longitud de fruto; DF: diámetro de fruto; RLDF: relación longitud/diámetro; GP: grosor de pulpa; DCC: diámetro de cavidad central; VCF: volumen comestible; FF: forma de fruto; FEP: forma del extremo peduncular; PA: prominencia de aristas; FCC: forma de la cavidad central.
El Grupo 1 se formó por 111 ejemplares que provienen de huertos familiares, y que se distribuyeron en cuatro subgrupos con similitud en el diámetro de cavidad central (Cuadro 2). Los frutos presentaron colores de pulpa de verde amarilla (5GY9/12) a roja (10R6/16). El Subgrupo 1-A exhibió frutos en forma de pera, con una relación longitud/diámetro de 2.4; el 1-B, frutos en forma de pera, pero con los valores altos (2.6) en la relación longitud/diámetro y la longitud (29.4 cm); el 1-C mostró valores intermedios tanto en las dimensiones como en la forma, al ser elíptica, con 1.7 de relación longitud/diámetro; en el 1-D, la forma del fruto fue globular, alcanzó el mayor diámetro (12.7 cm) y 1.8 de relación longitud/diámetro.
El Grupo 2 estuvo integrado por tres recolectas de papayo, con valor intermedio de longitud del fruto (Cuadro 2). Se distinguió de los Grupos 1 y 3 por mostrar mayor relación longitud/diámetro (3.4) y grosor de pulpa (3.2 cm), cavidad ovárica pequeña (2.0 cm de diámetro) y forma oblonga-aperada. Los colores de pulpa fueron de 2.5Y8/10 a 7.5YR7/12 (de amarilla a amarilla-anaranjada).
El Grupo 3 incluyó 108 recolectas subdivididas en cuatro subgrupos que mostraron semejanza en la longitud y diámetro del fruto con 6.6 cm y 4.5 cm, respectivamente (Cuadro 2). El color de pulpa de 5GY9/12 a 7.5Y9/8 (de verde-amarilla a amarilla) fue más frecuente en comparación con la amarilla-anaranjada (7.5YR7/12). Dentro de este grupo, 76 % de los ejemplares se localizaron en varios tipos de vegetación, en los que destaca la selva alta perennifolia, aunque algunos de ellos se situaron en los acahuales (Helianthus spp.) y el 24 % se encontró en huertos familiares. Al respecto, Caballero (1992) señaló que es posible encontrar este tipo de poblaciones en los huertos, ya que sus frutos contribuyen a la alimentación familiar. También Chávez-Pesqueira y Núñez-Farfán (2016) encontraron germoplasma silvestre en la vegetación de acahuales. En este Grupo 3, el dendrograma no logró diferenciar, dentro de tipos silvestres, los recolectados en vegetación natural de aquellos de huertos familiares. Este resultado tiene coherencia con las observaciones de Coppens dʼEeckenbrugge et al. (2007), quienes mediante marcadores morfológicos e izoenzimáticos caracterizaron germoplasma silvestre proveniente de áreas naturales y de huertos familiares, y tampoco encontraron diferencias.
El Subgrupo 3-A presentó principalmente frutos de forma elíptica al mostrar 1.7 de relación longitud/diámetro, con cavidad central tipo angular; el 3-B presentó valores intermedios (1.5) de relación longitud/diámetro, correspondiente a la forma redonda; el 3-C y 3-D mostraron forma oval al tener 1.2 de relación longitud/diámetro, pero ambos discreparon en la forma del extremo peduncular y prominencia de aristas.
Badillo (1971) y Manshardt y Zee (1994) encontraron en germoplasma silvestre frutos con longitud de 2 a 10 cm y 1.5 a 6 cm de diámetro, de forma redonda, pulpa amarilla o amarilla-anaranjada, pulpa escasa y cavidad central reducida; características similares a las encontradas en el Grupo 3 en el presente estudio.
CONCLUSIONES
La variabilidad morfológica de los ejemplares de C. papaya nativos de México descritos muestra importantes diferencias en la forma, dimensiones y tipo del extremo peduncular en fruto. Estas características del fruto son las que más contribuyen a la variabilidad morfológica entre el germoplasma evaluado; sin embargo, con base en las dimensiones del fruto se detectó un patrón gradual que podría estar asociado con el proceso de domesticación. Esto revela revela que los pobladores de las diferentes regiones probablemente se enfocaron en la obtención de frutos grandes con mucha pulpa comestible y diversidad de colores de pulpa. Paralelamente se pudo haber favorecido la presencia de plantas hermafroditas que no están presentes en las poblaciones silvestres estudiadas. Se identificaron tres grupos con ejemplares de papayo fenotípicamente diferente entre ellos. Se observó una continuidad en dimensiones y forma del fruto entre y dentro de grupos definidos en el análisis de conglomerados.

