











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Mejorar progenitores de híbridos comerciales de maíz (Zea mays L.) es una práctica común en los programas de mejoramiento genético, pues llega a ocurrir que uno de ellos presente una característica agronómica desfavorable de importancia económica que limita el uso intensivo de sus híbridos. Existen varios métodos para mejorar líneas (Hallauer y Miranda, 1988), el más común es la retrocruza (Márquez, 1988), con variantes como el mejoramiento convergente (Gómez-Montiel et al., 2013) y selección visual por pedigrí aprovechando la variación intralínea (Gómez-Montiel et al., 2013; Tokatlidis, 2000).
El progenitor B-3A es una línea S3 de excelente aptitud combinatoria general (ACG) y específica (ACE) para rendimiento de grano, participa en los híbridos comerciales H-380A (Ramírez et al., 2015), H-381A y H-382A, pero tiene poca tolerancia al acame, y ésta es una limitante importante, ya que históricamente los incrementos en rendimiento de grano se han logrado aumentando la densidad de población, y esta tendencia continúa, pero una restricción es que el acame de las plantas se eleva al aumentar la densidad de población (Duvick, 2005; Egli 2015; Liu et al., 2012; Sher et al., 2017).
El acame de las plantas es un problema complejo y multifactorial, causado por factores genéticos, vientos fuertes, plagas de la raíz (De la paz et al., 2007; Lugo-García et al., 2012; Pérez et al., 2006), barrenadores del tallo, especialmente del género Diatrea spp. y Calendra sp. (Ortega, 1987), enfermedades de la raíz y tallo (Figueroa-Rivera et al., 2010), tamaño y forma de la raíz (Stamp y Kiel, 1992), uso de altas densidades de población (Liu et al., 2012; Sher et al., 2017), competencia por luz (Xue et al., 2016), y morfología del tallo (Peiffer et al., 2013; Robertson et al., 2015; Robertson et al., 2016).
En la selección de líneas para tolerancia al acame de tallo se utilizan métodos tan simples como el de kick test (patada), y el más usado es el del penetrómetro, que consiste en medir la resistencia del tallo a la penetración con una aguja metálica (Peiffer et al., 2013). Las nuevas metodologías consideran la estructura del tallo y han informado que medir la flexibilidad del tallo es más eficiente que el uso del penetrómetro (Robertson et al., 2016); recientemente Xue et al. (2017) propusieron varias opciones para mejorar la tolerancia al acame al sembrar en altas densidades de población.
Con base en los resultados positivos de la selección visual de líneas (Ordás et al., 2012), y el aprovechamiento de la variación intralínea (Gómez-Montiel et al., 2013; Tokatlidis, 2000), se consideró que el progenitor B-3A tenía aún variabilidad genética para mejorar su tolerancia al acame, sin modificar la aptitud combinatoria de rendimiento de grano al avanzar la endogamia en el proceso de selección, lo cual era posible, pues la correlación genética entre una línea S3 y su cruza de prueba es 0.93 (Bernardo, 1991), y la de ACG entre líneas seleccionadas en generaciones tempranas y avanzadas es alta (Carlone y Russell, 1989; Lile y Hallauer, 1994). El objetivo de esta investigación fue mejorar la tolerancia al acame del progenitor B-3A mediante el aprovechamiento de su variación intralínea a través de generaciones de autofecundación, manteniendo la aptitud combinatoria para rendimiento de grano de la línea original.
Materiales y métodos
El material genético consistió de sublíneas seleccionadas a partir de la línea S3 de maíz de grano amarillo B-3A, las cuales se cruzaron con las líneas probadoras B-4A y B-6A, cuyo origen genético está descrito en Ramírez et al. (2007), Ramírez et al. (2015) y Ramírez et al. (2017).
El mejoramiento de la línea B-3A para tolerancia al acame se hizo a través de generaciones sucesivas de autofecundación mediante selección recurrente simple sin recombinación. Se seleccionaron inicialmente 150 familias de autohermanos de la línea B-3A. En cada generación de autofecundación se seleccionó entre familias para enfermedades foliares y dentro de familias para tolerancia al acame; para ello, en cada planta autofecundada se aplicó un “jalón-sacudida”, que consiste en seleccionar sólo plantas autofecundadas erectas y con mazorca completamente sana que hayan resistido un jalón y una sacudida fuerte aplicada en los tres primeros entrenudos del tallo, sin que éste se haya doblado, inclinado o arrancado de raíz. La selección de sublíneas se hizo en los ciclos agrícolas primavera-verano (PV) y otoño-invierno (OI). En PV se seleccionó en condiciones de temporal en la localidad de San Miguel Cuyutlán, municipio de Tlajomulco, Jal., ubicada a 1350 metros sobre el nivel del mar (msnm), y en OI se seleccionó en condiciones de riego en la localidad de Santiago Ixcuintla, Nay., ubicada a 20 msnm.
Después de siete generaciones de autofecundación de la línea B-3A se seleccionaron seis familias de un total de 150 familias de autohermanos, equivalente a 4 % de presión de selección, dentro de las cuales se seleccionaron 21 sublíneas S10. Con las sublíneas seleccionadas, en el ciclo OI 2012-2013, en la localidad de La Huerta, Jal. (350 msnm), se formaron cruzas de prueba con los probadores B-4A y B-6A, los cuales corresponden a la contraparte heterótica de B-3A (Ramírez et al., 2015; Ramírez et al., 2017).
En el ciclo PV de 2013, las cruzas de cada probador se evaluaron en ensayos uniformes individuales, pues sólo 14 cruzas fueron comunes con los dos probadores. Con el probador B-4A se evaluaron 15 sublíneas más la cruza original B-3A × B-4A, mientras que con B-6A se evaluaron 21 sublíneas más la cruza original B-3A × B-6A. Las 15 cruzas recobradas B-3A × B-4A se evaluaron en temporal en las localidades de Tepatitlán, Jal. (1810 msnm), Tlajomulco, Jal. (1350 msnm) y San José de Mojarras, Nay. (1084 msnm), y en punta de riego en Peña del Panal, municipio de Tarímbaro, Mich. (1897 msnm).
Las cruzas recobradas B-3A × B-6A se evaluaron en las mismas localidades, excepto que la localidad de Tepatitlán, Jal. se cambió por la de Iguala, Gro. (751 msnm). En ambos experimentos se usó el diseño experimental bloques completos al azar con dos repeticiones. La parcela experimental consistió de un surco de 4.0 m distanciado 0.8 m de sus adyacentes, con 25 plantas, equivalente a 78,125 plantas por hectárea. El manejo agronómico de los experimentos en cada sitio de prueba se hizo siguiendo las recomendaciones generadas para el cultivo de maíz por el Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias (INIFAP, 2017a; INIFAP, 2017b; INIFAP, 2017c, INIFAP, 2017d).
Las variables estudiadas fueron: a) rendimiento de grano en peso seco (kg ha-1), b) número de días a floración masculina, contados desde la siembra hasta que el 50 % de las plantas estuvieran liberando polen, c) número de días a floración femenina, contados desde la siembra hasta que el 50 % de las plantas exhibieron jilotes con estigmas expuestos con longitud mayor de 3.0 cm, d) altura de planta (cm), medida desde el ras del suelo hasta la punta de la espiga, e) altura de mazorca (cm), medida desde el ras del suelo hasta el nudo de la mazorca principal, f) porcentaje de acame de raíz, se calculó a partir del número de plantas acamadas de raíz dividida entre el número total de plantas de cada parcela (NTPL) y multiplicado por 100, y g) porcentaje de acame de tallo, calculado a partir del número de plantas acamadas de tallo dividido entre el NTPL y multiplicado por 100. Las plantas acamadas de raíz fueron aquellas desviadas 30o de la vertical o con cuello de ganso, mientras que las acamadas de tallo fueron plantas dobladas o quebradas abajo del nudo de la mazorca principal.
Se hicieron análisis combinados en cada grupo de cruzas y se consideró a las localidades y cruzas como efectos fijos y a las repeticiones como aleatorios. Al considerar que la variable acame de tallo estaba expresada en porcentaje, se hicieron análisis de varianza con los datos originales y transformados mediante la función arco seno propuesta por Snedecor y Cochran (1974), pero se presentan los originales porque el coeficiente de determinación (R2) fue muy similar en ambos análisis de la varianza. Se calcularon las correlaciones entre las variables de estudio y en la comparación de medias se aplicó la prueba Tukey y de Dunnett para comparar las medias de las cruzas recobradas con las originales B-3A × B-4A y B-3A × B-6A, respectivamente. Ambas pruebas se aplicaron al 0.05 de probabilidad de error. Además, se calculó la ACG para rendimiento de grano y acame de tallo, estimada por la diferencia entre el valor de la cruza y la media general de las cruzas (Sprague y Tatum, 1942), pero dado que se utilizaron líneas como probadores se considera como aptitud combinatoria específica (Márquez, 1988).
Resultados
Cruzas recobradas B-3A × B-4A
En el análisis de la varianza conjunto hubo diferencias estadísticas significativas entre ambientes (P ≤ 0.01) para todas las variables (Cuadro 1). Entre genotipos sólo hubo diferencias (P ≤ 0.05) en el número de días a floración masculina, mientras que la interacción genotipo × ambiente resultó significativa (P ≤ 0.05) para el porcentaje de acame de tallo, lo que indica que se tuvo una respuesta diferencial de las cruzas al acame en función del ambiente de evaluación.
Cuadro 1 Cuadrados medios y valores de probabilidad de cruzas recobradas formadas con sublíneas de B-3A y el probador B-4A.
| Factor de Variación | REND | FM† | FF† | AR | AT | AP | AM |
|---|---|---|---|---|---|---|---|
| Ambientes | 329,122,484 | 10,539 | 10,268 | 211.3 | 389.67 | 22,310 | 2452 |
| Probabilidad de F | ≤ .0001 | ≤ .0001 | ≤ .0001 | 0.03 | ≤ .0001 | 0.0008 | 0.018 |
| Genotipo | 912,078.3 | 1.63 | 2.24 | 9.47 | 135.7313 | 112.3328 | 42.8 |
| Probabilidad de F | 0.4897 | 0.057 | 0.08 | 0.97 | 0.22 | 0.24 | 0.78 |
| Genotipo × Ambiente | 993,820.7 | 0.72 | 1.50 | 13.8 | 182.2049 | 64.09253 | 71.2 |
| Probabilidad de F | 0.4063 | 0.72 | 0.33 | 0.95 | 0.02 | 0.86 | 0.29 |
| CV (%) | 10.4 | 1.3 | 1.5 | 273.2 | 54.1 | 3.4 | 6.1 |
| R2 | 0.95 | 0.998 | 0.997 | 0.53 | 0.92 | 0.93 | 0.77 |
†En el análisis de varianza no se incluyó la localidad de Tlajomulco. REND: rendimiento de grano; FM: días a floración masculina; FF: días a floración femenina; AR: porcentaje de acame de raíz; AT: porcentaje de acame de tallo; AP: altura de planta; AM: altura de mazorca.
La media de rendimiento de grano más alta (12,701 kg ha-1) de las cruzas recobradas B-3A × B-4A se obtuvo en la localidad de Peña del Panal, y le siguieron las localidades de Tlajomulco, Tepatitlán y San José de Mojarras (Cuadro 2), con diferencias significativas (P ≤ 0.05) entre ellas. Las cruzas tuvieron ciclo de madurez intermedio-tardío, la media más alta de acame de raíz se registró en Peña del Panal (5.5 %) y fue estadísticamente superior al resto de las localidades, mientras que la media de acame de tallo más alta se obtuvo en San José de Mojarras (52.9 %) y fue estadísticamente (P ≤ 0.05) diferente al resto de localidades, y entre estas últimas no hubo diferencias significativas (Cuadro 2).
Cuadro 2 Rendimiento de grano y características agronómicas de cruzas recobradas formadas con sublíneas de B-3A y el probador B-4A, en las localidades de evaluación.
| Localidad | REND | FM† | FF† | AR | AT | AP | AM |
|---|---|---|---|---|---|---|---|
| Peña del Panal, Mich. | 12,701 a | 84.0 a | 84.8 a | 5.5 a | 0.8 b | 270 c | 131 b |
| Tlajomulco, Jal. | 10,500 b | 1.0 b | 2.6 b | 307 a | 139 a | ||
| Tepatitlán, Jal. | 8881 c | 87.0 b | 87.0 b | 0.4 b | 2.5 b | 286 b | 121 c |
| San José de Mojarras, Nay. | 5096 d | 54.2 b | 54.9 c | 0.0 b | 52.9 a | 245 d | 121 c |
| DSH (0.05) | 638 | 0.6 | 0.7 | 3.1 | 6.7 | 6 | 5 |
†No se tomaron datos de FM y FF en la localidad de Tlajomulco. Medias con letras iguales no son estadísticamente diferentes (Tukey 0.05). REND: rendimiento de grano (kg ha-1); FM: días a floración masculina; FF: días a floración femenina; AR: porcentaje de acame de raíz; AT: porcentaje de acame de tallo; AP: altura de planta (cm); AM: altura de mazorca (cm); DSH: diferencia significativa honesta.
Al comparar las medias por genotipo, sólo hubo diferencias significativas (P ≤ 0.05) en el número de días a floración masculina, donde, de acuerdo con la prueba de Dunnet, la cruza CR (3A × 4A)-9 fue significativamente más tardía (76 d) (P ≤ 0.05) que la original B3A × B-4A (74 d), y no hubo diferencias significativas en rendimiento de grano entre las cruzas recobradas y la original, y las diferencias entre la mejor y la peor cruza oscilaron entre -10 y 5 %, respectivamente. En acame de tallo no hubo significancia entre las cruzas, pero es de resaltar que las cruzas CR(3A × 4A)-8 y CR(3A × 4A)-6 tuvieron valores de acame total, raíz más tallo, de 7 y 9 %, respectivamente, valores inferiores al 18 % de la cruza original B3A × B-4A (Cuadro 3).
Cuadro 3 Cruzas recobradas con sublíneas de B-3A usando como probador la línea B-4A. Promedio de cuatro localidades.
| Genealogía | REND | FM† | FF† | AR | AT | AP | AM |
|---|---|---|---|---|---|---|---|
| CR(3A × 4A)-12 | 9886 | 74 | 75 | 2 | 14 | 271 | 126 |
| CR(3A × 4A)-10 | 9602 | 76 | 77 | 2 | 12 | 281 | 131 |
| CR(3A × 4A)-8 | 9545 | 75 | 76 | 0 | 7 | 281 | 128 |
| CR(3A × 4A)-6 | 9511 | 75 | 76 | 3 | 6 | 282 | 127 |
| CR(3A × 4A)-11 | 9495 | 75 | 76 | 2 | 14 | 280 | 130 |
| CR(3A × 4A)-9 | 9485 | 76 | 77 | 1 | 11 | 279 | 131 |
| CR(3A × 4A)-13 | 9395 | 75 | 76 | 3 | 18 | 279 | 130 |
| CR(3A × 4A)-5 | 9369 | 75 | 75 | 1 | 13 | 276 | 124 |
| CR(3A × 4A)-2 | 9288 | 75 | 76 | 0 | 16 | 275 | 130 |
| CR(3A × 4A)-7 | 9243 | 76 | 76 | 4 | 14 | 280 | 130 |
| CR(3A × 4A)-4 | 9121 | 75 | 75 | 1 | 18 | 276 | 126 |
| CR(3A × 4A)-3 | 9055 | 75 | 76 | 2 | 17 | 269 | 126 |
| CR(3A × 4A)-1 | 9048 | 75 | 76 | 3 | 18 | 278 | 129 |
| CR(3A × 4A)-14 | 8800 | 75 | 76 | 2 | 21 | 276 | 128 |
| CR(3A × 4A)-15 | 8482 | 75 | 75 | 2 | 20 | 274 | 125 |
| Media de cruzas recobradas | 9288 | 75 | 76 | 1.7 | 14.6 | 277 | 128 |
| Cruza original B3A × B-4A | 9388 | 74 | 74 | 2.5 | 16.1 | 279 | 129 |
| Media general | 9295 | 75 | 76 | 1.7 | 14.7 | 277 | 128 |
| CV (%) | 10.4 | 1.3 | 1.5 | 273.2 | 68.9 | 3.4 | 6.1 |
| DSH (0.05) | 1727 | 2.0 | 2.4 | 8.4 | 18 | 17 | 14 |
| Probabilidad de F | 0.4897 | 0.057 | 0.082 | 0.97 | 0.22 | 0.24 | 0.78 |
†Los valores de FM y FF corresponden al promedio de las localidades de Tepatitlán, Santa María del Oro y Peña del Panal. REND: rendimiento de grano; FM: días a floración masculina; FF: días a floración femenina; AR: porcentaje de acame de raíz; AT: porcentaje de acame de tallo; AP: altura de planta; AM: altura de mazorca; ns: no significativo; DSH: diferencia significativa honesta.
Hubo una asociación negativa entre los efectos de ACE del rendimiento de grano y el acame de tallo (Figura 1), cuyo coeficiente de correlación fue negativo (r = -0.70) y significativo (P ≤ 0.05), donde la cruza original tendió a ubicarse en la media de las cruzas recobradas. Asimismo, la correlación entre acame de tallo y altura de planta fue negativa (r = -0.55) y significativa (P ≤ 0.05).
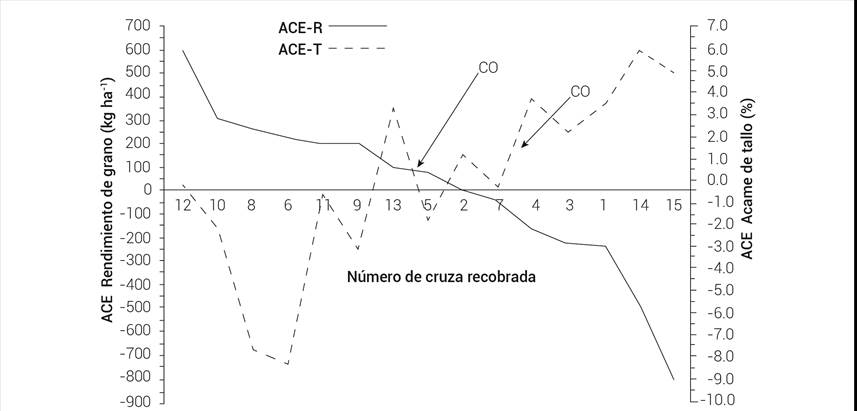
Figura 1 Aptitud combinatoria específica para rendimiento de grano (ACE-R) y porcentaje de acame de tallo (ACE-T) de cruzas recobradas formadas con sublíneas de B-3A y el probador B-4A. CO: cruza original B-3A × B-4A.
Cruzas recobradas de B-3A × B-6A
En el análisis de varianza conjunto hubo diferencias estadísticas significativas para todas las variables entre ambientes, excepto para porcentaje de acame de raíz (Cuadro 4). Entre genotipos hubo diferencias significativas en el número de días a floración masculina y femenina, y altura de planta y de mazorca; el rendimiento de grano y porcentaje de acame no tuvieron significancia. La interacción genotipo × ambiente resultó significativa (P ≤ 005) para el número de días a floración masculina y femenina, porcentaje de acame de tallo y altura de planta y de mazorca (Cuadro 4).
Cuadro 4 Cuadrados medios y valores de probabilidad de cruzas recobradas formadas con sublíneas de B-3A y el probador B-6A.
| Factor de Variación | REND | FM† | FF† | AR†† | AT†† | AP | AM |
|---|---|---|---|---|---|---|---|
| Ambientes | 512,643,515 | 10904 | 10725 | 68.05 | 54541 | 23253 | 3740 |
| Probabilidad de F | 0.0003 | <.0001 | <.0001 | 0.10 | 0.001 | 0.001 | 0.012 |
| Genotipos | 1,535,486 | 1.73 | 2.661 | 3.55 | 179.6 | 183.3 | 73.67 |
| Probabilidad de F | 0.2184 | 0.06 | 0.016 | 0.65 | 0.22 | 0.01 | 0.034 |
| Genotipo x Ambiente | 1,144,389 | 1.77 | 2.661 | 2.16 | 313.3 | 137.3 | 80.55 |
| Probabilidad de F | 0.5856 | 0.03 | 0.005 | 0.99 | 0.0017 | 0.03 | 0.002 |
| CV (%) | 13.4 | 1.5 | 1.7 | 202.7 | 46.7 | 3.4 | 4.9 |
| R2 | 0.94 | 0.997 | 0.996 | 0.564 | 0.94 | 0.92 | 0.84 |
†En el análisis de varianza no se incluyó la localidad de Tlajomulco. ††En el análisis de la varianza no se incluyó la localidad de Iguala. REND: rendimiento de grano; FM: días a floración masculina; FF: días a floración femenina; AR: porcentaje de acame de raíz; AT: porcentaje de acame de tallo; AP: altura de planta; AM: altura de mazorca.
El promedio de rendimiento de grano fue significativo (P ≤ 0.05) entre localidades y el más alto se obtuvo en Peña del Panal (12,868 kg ha-1) y le siguieron las localidades de Tlajomulco, Iguala y San José de Mojarras (Cuadro 5). El número de días a floración masculina y femenina entre ambientes resultó significativo (P ≤ 0.05) y el más alto se registró en Peña del Panal, seguido de Iguala y San José de Mojarras. En acame de tallo las diferencias fueron significativas entre ambientes; el promedio más alto se registró en San José de Mojarras (65.6 %), seguido por Tlajomulco (9.5 %) y Peña del Panal (0.6 %). En altura de planta hubo significancia (P ≤ 0.05) entre ambientes, y el promedio más alto se registró en Tlajomulco, seguido por Iguala, Peña del Panal y San José de Mojarras; sin embargo, la tendencia en altura de mazorca fue diferente, ya que el promedio más alto se registró en Tlajomulco, seguido por Peña del Panal y San José de Mojarras con diferencias significativas (P ≤ 0.05), lo cual confirma la interacción genotipo × ambiente significativa para estas características (Cuadros 4 y 5).
Cuadro 5 Rendimiento de grano y características agronómicas de cruzas recobradas formadas con sublíneas de B-3A y el probador B-6A en las localidades de prueba.
| Localidad | REND | FM† | FF† | AR†† | AT†† | AP | AM |
|---|---|---|---|---|---|---|---|
| Peña del Panal, Mich. | 12868 a | 84.2 a | 84.7 a | 2.4 | 0.6 c | 271.0 b | 132 b |
| Tlajomulco, Jal. | 7833 b | 0.5 | 9.5 b | 304.4 a | 143 a | ||
| Iguala, Gro. | 7295 b | 59.4 b | 60.3 b | 276.3 b | 121 c | ||
| San José de Mojarras, Nay. | 4705 c | 55.0 c | 55.7 c | 0.1 | 65.6 a | 248.4 c | 129 b |
| DSH (0.05) | 614 | 0.5 | 0.6 | 1.0 | 6.0 | 5.3 | 4 |
†En el análisis de varianza no se incluyó la localidad de Tlajomulco. ††En el análisis de la varianza no se incluyó la localidad de Iguala. Medias con letras iguales no son estadísticamente diferentes (Tukey 0.05). REND: rendimiento de grano; FM: días a floración masculina; FF: días a floración femenina; AR: porcentaje de acame de raíz; AT: porcentaje de acame de tallo; AP: altura de planta (cm); AM: altura de mazorca (cm); ns: no significativo; DSH: diferencia significativa honesta.
Al comparar las medias de genotipos, sólo hubo diferencias significativas en el número de días a floración masculina y femenina y altura de planta y de mazorca, pues con base en la prueba de Dunnett, la cruza CR(3A × 6A)-17 fue significativamente (P ≤ 0.05) más tardía (68 d) que la original B-3A × B-6A (66 d), y la cruza CR(3A × 6A)-9 fue significativamente diferente (P ≤ 0.05) en altura de planta (287 cm) y mazorca (142 cm) que la original B-3A × B-6A, que tuvo valores de 270 y 131 cm, respectivamente, y no hubo diferencias estadísticas entre genotipos para rendimiento de grano y porcentaje de acame de tallo, pero es de resaltarse que sobresalió la cruza CR(3A × 6A)-10, en la cual se combinaron un mayor rendimiento de grano (9145 kg ha-1) con menor porcentaje de acame total (11.7 %) que el registrado en la cruza original B-3A × B-6A, que fueron de 8153 kg ha-1 y 20.5 %, respectivamente (Cuadro 6).
Cuadro 6 Rendimiento de grano de cruzas recobradas formadas con sublíneas de B-3A usando como probador la línea B-6A. Promedio de cuatro localidades.
| Genealogía | REND | FM† | FF† | AR†† | AT†† | AP | AM |
|---|---|---|---|---|---|---|---|
| CR(3A × 6A)-10 | 9145 | 66 | 66 | 0.0 | 11.7 | 272 | 129 |
| CR(3A × 6A)-11 | 8915 | 66 | 66 | 0.0 | 19.3 | 278 | 130 |
| CR(3A × 6A)-12 | 8751 | 66 | 67 | 0.0 | 27.5 | 277 | 131 |
| CR(3A × 6A)-6 | 8679 | 66 | 67 | 1.3 | 32.7 | 276 | 131 |
| CR(3A × 6A)-8 | 8490 | 66 | 66 | 2 | 30.3 | 281 | 134 |
| CR(3A × 6A)-15 | 8410 | 67 | 68 | 1.3 | 25.0 | 278 | 134 |
| CR(3A × 6A)-5 | 8304 | 66 | 66 | 1.3 | 22.7 | 271 | 130 |
| CR(3A × 6A)-1 | 8265 | 66 | 68 | 1.7 | 28.8 | 278 | 131 |
| CR(3A × 6A)-7 | 8222 | 66 | 67 | 0.7 | 25.7 | 279 | 130 |
| CR(3A × 6A)-20 | 8198 | 66 | 67 | 1.3 | 18.8 | 269 | 129 |
| CR(3A × 6A)-16 | 8117 | 66 | 67 | 1.3 | 19.8 | 279 | 135 |
| CR(3A × 6A)-13 | 8108 | 67 | 67 | 0.7 | 25.3 | 275 | 132 |
| CR(3A × 6A)-14 | 8090 | 67 | 67 | 0.7 | 29.7 | 279 | 133 |
| CR(3A × 6A)-18 | 8042 | 66 | 68 | 1.3 | 23.7 | 269 | 130 |
| CR(3A × 6A)-4 | 8022 | 66 | 67 | 1.2 | 32.7 | 273 | 129 |
| CR(3A × 6A)-2 | 7898 | 66 | 67 | 0.0 | 27.5 | 277 | 133 |
| CR(3A × 6A)-3 | 7690 | 66 | 67 | 0.7 | 30.7 | 268 | 129 |
| CR(3A × 6A)-9 | 7686 | 66 | 68 | 0.5 | 21.5 | 287 | 142 |
| CR(3A × 6A)-21 | 7653 | 66 | 67 | 0.3 | 31.2 | 273 | 128 |
| CR(3A × 6A)-19 | 7581 | 66 | 67 | 0.7 | 30.2 | 270 | 130 |
| CR(3A × 6A)-17 | 7440 | 68 | 69 | 2.7 | 20.2 | 272 | 130 |
| Media de cruzas recobradas | 8176 | 66 | 67 | 0.9 | 25.5 | 275 | 131 |
| B-3A × B-6A | 8153 | 66 | 67 | 2.5 | 20.5 | 270 | 131 |
| Media general | 8175 | 66 | 67 | 1 | 25.2 | 275 | 131 |
| CV (%) | 13.4 | 1.5 | 1.7 | 202.7 | 46.7 | 3.4 | 4.9 |
| DSH (0.05) | 2040 | 2.2 | 2.5 | 4.4 | 25.5 | 17 | 12 |
| Probabilidad de F | 0.22 | 0.6 | 0.016 | 0.65 | 0.22 | 0.01 | 0.034 |
†En el análisis de varianza no se incluyó la localidad de Tlajomulco. ††En el análisis de la varianza no se incluyó la localidad de Iguala. REND: rendimiento de grano Kg ha-1; FM: días a floración masculina; FF: días a floración femenina; AR: porcentaje de acame de raíz; AT: porcentaje de acame de tallo; AP: altura de planta (cm); AM: altura de mazorca (cm); ns: no significativo; DSH: diferencia significativa honesta.
Los efectos de ACE del rendimiento de grano y los valores del acame de tallo tuvieron una asociación negativa (Figura 2), con un coeficiente de correlación negativo (r = -0.32), pero no significativo, y sólo la correlación entre el rendimiento de grano y número de días a floración femenina fue negativa (r = -0.57) y significativa (P ≤ 0.05). Se observó que la media de rendimiento de la cruza original B-3A × B-6A se ubicó casi en la media de las cruzas recobradas.
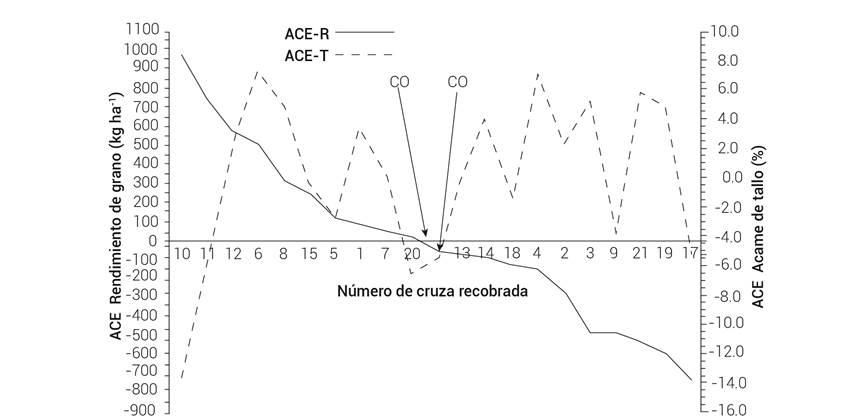
Discusión
Las diferencias observadas en rendimiento de grano entre ambientes se deben a que las cruzas recobradas y originales se evaluaron en ambientes contrastantes en altitud y humedad, ya que oscilaron desde 751 msnm (Iguala, Gro.) hasta 1847 msnm (Peña del Panal, Mich.), y comprendieron sistemas de temporal y punta de riego. El acame de tallo fue más importante que el de raíz y, consistentemente, en San José de Mojarras se registró el porcentaje de acame más alto (Cuadros 1, 2, 4 y 5), debido a que en la etapa R5 hubo ataque de Fusarium spp. en el tallo; esta enfermedad es una de las más importantes en Nayarit, que constituye un sitio ideal para seleccionar para tolerancia al acame de tallo.
No hubo diferencias estadísticas en rendimiento de grano entre las cruzas originales B-3A × B-4A y B-3A × B-6A y sus cruzas recobradas formadas a partir de sublíneas S10 de B-3A (Cuadros 1, 3, 4 y 6); este resultado confirma que la aptitud combinatoria en las líneas de maíz se mantiene desde las primeras generaciones de autofecundación (Carlone y Russell, 1989; Lile y Hallauer, 1994; Sprague, 1946; Wellhausen y Wortman, 1954), y está en concordancia con las correlaciones genéticas de 0.93 y 0.97 entre la línea y su cruza de prueba, evaluadas en las generaciones S3 y S4, respectivamente (Bernardo, 1991). Por tanto, los programas de mejoramiento genético de maíz con recursos limitados y sin acceso a tecnologías más caras de hibridación, como los dobles haploides, pueden formar híbridos comerciales de maíz competitivos mediante líneas S3 ó S4 de probadas ACG y ACE, pero con una buena estrategia en el mantenimiento de las líneas parentales (Chávez et al., 2005).
Con respecto a la selección para acame de tallo, no hubo diferencias estadísticas entre las cruzas recobradas y la original (P ≤ 0.22) (Cuadros 1 y 4), lo que indica que el método del “jalón-sacudida” no fue eficiente para seleccionar plantas con tolerancia al acame de tallo en generaciones avanzadas de líneas S3, debido probablemente a que éstas de inicio presentan mucha uniformidad (F = 0.875), o posiblemente la varianza aditiva para acame (plantas tolerantes) era muy poca entre y dentro de familias. No obstante, las cruzas recobradas CR(3A × 4A)-8, CR(3A × 4A)-6 y CR(3A × 6A)-10 fueron las más prometedoras porque combinaron media de rendimiento de grano mayor que el testigo y acame de tallo hasta 100 % menor que las cruzas originales (Cuadros 3 y 6), lo que indica que hubo sublíneas superiores en acame, pero en muy baja frecuencia.
Si se desea mejorar la tolerancia al acame de tallo en líneas S3, o con endogamia superior, e incrementar la frecuencia de líneas tolerantes, es mejor introgresar germoplasma de un donador reconocido por su buena ACG para rendimiento y tolerancia al acame de tallo, usar el método de retrocruza y aplicar técnicas cuantitativas en la selección como las usadas por Peiffer et al. (2013) y Robertson et al. (2016), aun cuando en la selección visual haya resultado alentadora (Gómez-Montiel et al., 2013; Ordás et al., 2012; Tokatlidis, 2000).
La correlación entre el rendimiento de grano y el acame de tallo fue mayor en las cruzas recobradas de B-3A × B-4A (-0.70) que en B-3A × B-6A (-0.32), probablemente porque en las cruzas recobradas de B-3A × B-6A los daños al tallo se presentaron en etapas más tempranas que en las cruzas recobradas de B-3A × B-4A; al respecto, Scauflaire et al. (2011) informaron que las infecciones por Fussarium spp. al tallo y mazorca se inician en algunas plantas desde la antesis y ésta avanza conforme se alcanza la etapa de madurez, lo que puede infectar hasta 100 % de las plantas.
Con respecto al mejoramiento del rendimiento de grano en maíz, estudios sobre la relación fuente-demanda indican que se ha logrado poca ganancia genética en el número de granos por mazorca y el número de mazorcas por planta, por lo que, los incrementos históricos en rendimiento de maíz han estado más asociados con incrementos en la densidad de población, tolerancia a factores adversos y prácticas de cultivo (Duvick 2005; Egli, 2015), lo que muestra que a pesar de la complejidad que implica mejorar para tolerancia al acame, dicho mejoramiento debe hacerse por la relación tan estrecha entre el acame y las altas densidades de población (Liu et al., 2012; Sher et al., 2017; Xue et al., 2017).
El comportamiento de los valores de la ACE del rendimiento de grano de las cruzas recobradas fue similar en ambos probadores, y en ambos casos la cruza original se ubicó casi en la media de la población (Figuras 1 y 2), por lo que en el avance endogámico la línea tuvo variaciones pequeñas positivas y negativas en capacidad de combinación, que aunque no fueron significativas, indican que en la selección de sublíneas, éstas deben evaluarse en cruzas para identificar las de mayor ACE y evitar perder aptitud combinatoria para rendimiento de grano con respecto a la línea original. Otro aspecto importante del comportamiento centrista de la cruza original se relaciona con el mantenimiento de las líneas progenitoras S3, pues éstas podrían mantenerse alternando una o dos generaciones de autofecundación, para purificar la línea de posibles contaminantes, con un incremento de semilla mediante cruzas fraternales, para así mantener la aptitud combinatoria de la línea y recuperar su vigor mediante los tamaños de muestra propuestos por Chávez et al. (2005) o Carlone y Russell (1988).
Conclusiones
La selección a partir de una línea S3 para tolerancia al acame en generaciones sucesivas de autofecundación no fue un método eficiente, pues en la generación S10 sólo dos sublíneas tuvieron 100 % menos acame que la cruza original. La aptitud combinatoria de la línea B-3A para rendimiento de grano se mantuvo de la generación S3 a la S10, ya que no hubo diferencias estadísticas significativas entre el rendimiento de grano de la cruza original y el de las recobradas.

