











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El trigo (Triticum aestivum L.) es uno de los cultivos alimenticios de primera necesidad, se producen alrededor de 700 millones de toneladas en el mundo (FAO, 2018), en altitudes que van de 0 a 3000 msnm (Huerta y Singh, 2000). Puccinia graminis f. sp. tritici (Pgt), patógeno causante de la roya del tallo del trigo es un parásito biótrofo especializado, con una alta variación genética para virulencia, debido a mutaciones y recombinación genética (Singh et al., 2011a). El mejoramiento genético aplicado al trigo ha reducido los daños por enfermedades, lo que ha incrementado la productividad y el mejor uso de los insumos. Entre los futuros retos del fitomejoramiento destacan la escasez de agua para el riego, lluvias más erráticas, mayor fluctuación de las temperaturas (ondas de calor y heladas) y la gran variación de razas de roya que, en su conjunto, podrían acabar con la producción nacional (Rajaram et al., 1988).
En México, la roya del tallo del trigo, se controló genéticamente en 1955 gracias al gen Sr2 derivado de la variedad Hope (McIntosh et al., 1995), gen que incluso a la fecha es efectivo; sin embargo, la raza TTKSK comúnmente denominada Ug99 (Jin et al., 2007; 2008) ha sido devastadora, causando pérdidas del 50 al 100 % en el continente africano y se estima que podría llegar a México dentro de 5 a 7 años, debido a que existe la amenaza continua de la evolución del patógeno ante las condiciones climáticas cambiantes, por lo que es necesario contar con material genético con resistencia ante las posibles mutaciones del patógeno, ya que al menos el 75 % de las variedades recomendadas en el país actualmente basan su resistencia en los efectos del gen Sr2 (Singh et al., 2011b).
Para lograr resistencia durable a la roya del tallo es necesario conjuntar de cuatro a cinco genes de efectos menores con acción génica aditiva, lo que se conoce como resistencia cercana a la inmunidad (Singh et al., 2002). Los genes que confieren resistencia de desarrollo lento de la enfermedad (slow rusting) tienen un efecto que fluctúa de pequeño a moderado, lo que aumenta su eficacia a medida que se involucran más genes aditivos (Rajaram et al., 1988). La combinación de mayor número de genes diferentes de resistencia con efectividad parcial y total, proporciona resistencia durable y estable (Anderson, 2000). En diversos países se ha detectado germoplasma con buenos niveles de resistencia a esta enfermedad, mismo que una vez caracterizado, se podría incorporar a las líneas élite de México.
Valderrama (1975) reportó la variedad Bonza 63, desarrollada en Colombia, como fuente de resistencia a roya del tallo, alto rendimiento y características de calidad deseables para la industria panificadora. El Instituto Nacional de Investigaciones Agropecuarias de Ecuador, en 1974 generó la variedad Romero 73 con la característica de poseer alta resistencia a Pgt (Lalama, 1974). La variedad de trigo harinero Huites M95 que fue ampliamente sembrada en el norte de México, posee genes de resistencia a royas de la hoja y amarilla (Rodríguez et al., 2009). En estudios preliminares realizados en el Kenyan Agriculture Livestock Research Institute (KALRO) en Njoro, Kenya, se evaluaron los genotipos Bonza 63, Romero 73 y LC-150 contra la raza TTKSK, en los cuales se observó 0 y 10 % de infección, respectivamente; así mismo Cacuke, Apav-14 y Apav-92 presentaron susceptibilidad a Ug99.
En los últimos años se han utilizado Huites M95 y LC-150 por su resistencia a roya del tallo en los programas de mejoramiento genético de trigo en México, de tal manera que resulta importante conocer la herencia de su resistencia y la posibilidad de acumular genes de efecto aditivo, motivo por el cual en México se introdujo Cacuke, Romero 73 y Bonza 63 para identificar nuevas fuentes de resistencia. El objetivo de la presente investigación fue determinar la herencia de la resistencia a la severidad del daño causado por Puccinia graminis f. sp. tritici raza RTR en las progenies F3 de las cruzas entre los genotipos Huites M95 y LC-150 y los progenitores Cacuke, Bonza 63, Romero 73, Apav-14 y Apav-92.
Materiales y métodos
Obtención de generaciones F1, segregantes F2 y familias F3
Los progenitores se sembraron en el vivero de cruzamientos del INIFAP-Campo Experimental Valle de México (CEVAMEX) y se realizaron las cruzas de los genotipos resistentes Huites M95 y LC-150 con los genotipos susceptibles Romero 73, Bonza 63, Cacuke, Apav-14 y Apav-92, de donde se originaron las poblaciones Huites M95 × Romero 73, Huites M95 × Bonza 63, Huites M95 × Cacuke, Huites M95 × Apav-14 y Huites M95 × Apav-92; LC-150 × Bonza 63 y LC-150 × Cacuke. La F1 se sembró en parcelas de cuatro surcos de 1.0 m de longitud, de manera espaciada en el ciclo P-V/2012 en el CEVAMEX, ubicado en Chapingo, México. Se cosecharon todas las plantas F1 de manera individual y de cada cruza se eligieron tres plantas para sembrar su semilla de manera espaciada y avanzarlas a la generación F2 en el ciclo O-I/2012-2013, en el INIFAP, Campo Experimental Bajío, Guanajuato, México. Se obtuvieron entre 98 y 118 familias F3 derivadas de los cruzamientos.
Manejo experimental e inoculaciones
Las familias F3 se sembraron en el CEVAMEX, en los ciclos P-V/2013 y 2014. Se utilizó una mezcla de genotipos susceptibles a la raza RTR como fuente dispersante de inóculo. Dicha mezcla se sembró en la periferia de las poblaciones en bordos con doble surco y en los espacios entre parcelas.
La raza de roya del tallo usada fue RTR cuya avirulencia/virulencia es: Sr 7a, 10,14, Dp2, H/Sr5, 6, 7b, 8a, 8b, 9a, 9b, 9d, 11, 17, 21, 36 (Singh, 1991). La inoculación se llevó a cabo mediante aspersores manuales a los 30, 35 y 37 días después de la siembra con una suspensión de urediniosporas en aceite mineral Soltrol® como vehículo de dispersión a una concentración de 1 × 106 urediniosporas mL-1. Las condiciones de temperatura (20 y 30 ºC) y humedad (mayor a 70 %) fueron favorables para el desarrollo de la enfermedad.
Registro de datos
La primera toma de datos se realizó una vez que el progenitor susceptible de cada cruza alcanzó de 90 a 100 % de infección en la hoja bandera y la segunda evaluación 7 d después. Se registró el porcentaje de infección de acuerdo con la escala modificada de Cobb (Peterson et al., 1948). En las familias heterocigóticas, identificadas por su segregación, se utilizó la misma escala y se registró el dato promedio de infección. Cada familia F3 se clasificó en cuatro grupos (Rajaram et al., 1988), Grupo 1: familias homocigóticas con una respuesta similar a la del progenitor resistente; Grupo 2: familias homocigóticas con una respuesta similar a la del progenitor susceptible; Grupo 3: familias heterocigóticas segregantes, hasta un porcentaje intermedio y Grupo 4: familias heterocigóticas segregantes en las que se agrupan todas las categorías, de plantas tan resistentes como el progenitor resistente, intermedias y tan susceptibles como el progenitor susceptible. La primera toma de datos permitió clasificar las familias en grupos, y la segunda se utilizó para corroborar los datos.
Análisis de datos y pruebas estadísticas
Las frecuencias esperadas de las familias F3 en planta adulta fueron bajo el supuesto de que la resistencia es condicionada por, al menos, un gen de efecto aditivo y que las frecuencias de las familias homocigóticas susceptibles determinan el número de genes de resistencia.
La frecuencia de familias susceptibles similares al progenitor susceptible, sirve de base para determinar el número de genes de acuerdo con la proporción esperada (Singh et al., 2001). Las familias susceptibles son más fáciles de identificar en campo bajo el supuesto de que la virulencia del patógeno es recesiva y que la resistencia en la planta es dominante, siguiendo la teoría del gen por gen (Roelfs y Groth, 1988). Las frecuencias esperadas en las familias F3 en los cuatro grupos de clasificación se calcularon bajo el supuesto de que la resistencia está condicionada por 2, 3, 4 y 5 genes de efecto aditivo (frecuencias de familias homocigóticas susceptibles de 1/16, 1/64, 1/256 y 1/1024, respectivamente) de acuerdo con el Cuadro 1. Si la resistencia estuviera controlada por dos genes, las frecuencias esperadas de familias homocigóticas susceptibles serían de 6.25 % (1/16); si ésta fuera condicionada por tres genes, la proporción sería de (1/64) y si estuviera controlada por cuatro genes, la proporción sería de 0.4 % (1/256).
Cuadro 1 Frecuencias esperadas (%) en las cuatro categorías en que se clasifica la F3 en cruzas susceptible por resistente, según el número de genes involucrados.
| No. de genes | Categoría | |||
|---|---|---|---|---|
| 1 LHRPR | 2 LHSPS | 3 Seg I | 4 Seg S | |
| 2 | 6.3 | 6.3 | 37.5 | 50 |
| 3 | 1.6 | 4.6 | 56.3 | 40.6 |
| 4 | 0.4 | 0.4 | 68 | 31.3 |
| 5 | 0.1 | 0.1 | 76.2 | 23.6 |
LHRPR: líneas homocigóticas como el progenitor resistente; LHSPS: líneas homocigóticas como el progenitor susceptible; Seg I: líneas segregantes intermedias, pero sin plantas completamente susceptibles; Seg S: líneas segregantes desde completamente resistentes hasta completamente susceptibles.
Se realizó una prueba de Ji-cuadrada (X2) con las frecuencias observadas y esperadas, con n-1 grados de libertad, donde n es el número de grupos de clasificación de familias F3 (Infante y Zárate, 1998).
Análisis molecular
Los progenitores se sembraron en condiciones de invernadero y 15 días después de la emergencia se cosecharon 2 g de tejido foliar en tubos (Micro Tube Strips of 8 Attached 1.1 mL de Micro Pack®) colocados en placas de 96 piezas (Colection Microtubes Cracked 10 × 96) las cuales se mantuvieron en congelación durante 2 d para posteriormente liofilizar. El ADN se extrajo mediante la técnica CTAB siguiendo los protocolos de Dreisigacker et al. (2013). Cada muestra de ADN se cuantificó con un espectrofotómetro NanoDrop 8000 (Thermo Scientific®) y se ajustó a 50 ng µL-1.
La caracterización génica se realizó con los marcadores SNP Lr46jf2-2, Lr46_SNP1622, CSLV67, Lr34_TCCIND, Lr68-2 y Sr2_ger9 3p; para los genes Lr46/Sr58, Lr67/Yr46, Lr34/Sr57, Lr68 y Sr2, respectivamente, los cuales confieren resistencia a royas en planta adulta.
Para la PCR se utilizaron 5 µL de ADN, mismos que se colocaron en microplacas de PCR de 384 pozos de la marca Midsci® (St. Louis, MO, USA), donde se dejaron para su secado. La mezcla de reacción se preparó con un volumen final de 5 µL, 2.5 µL de 2X KASPAR, 0.1 µL Assay Mix, 0.2 µL ddH2O por muestra, en el termociclador, con el programa Touchdown (TD) (1 ciclo: 94 ºC por 15 min, 11 ciclos: 94 ºC por 30 seg, 65 ºC por 1 min, 72 ºC por 30 seg, 26 ciclos: 94 ºC por 30 seg, 57 ºC por 1 min, 72 ºC por 30 seg, 1 ciclo: 72 ºC por 2 min y una extensión final a 72 ºC por 15 min); posteriormente, se realizó la lectura en un lector de placas de fluorescencia (Pherastar Plus de BMG Labtech® de Ortenberg, Alemania).
Resultados y discusión
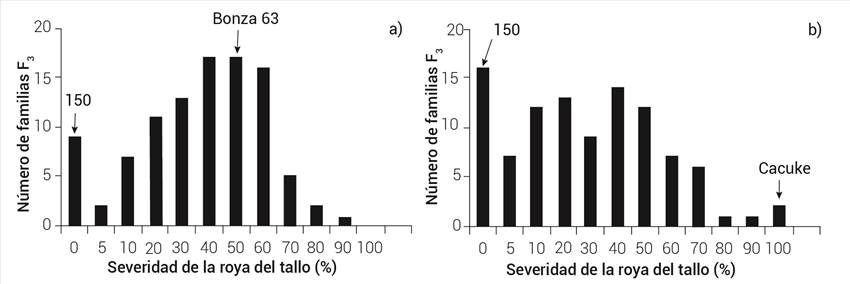
En los dos ciclos de evaluación no se observó una distribución discreta de clases que indica la presencia de genes de resistencia de efectos mayores o factores de resistencia de dominancia completa. En vez de esto, se observó una variación continua en la expresión de la resistencia de las familias F3, típica de un tipo de herencia poligénica (Figuras 1 y 2).
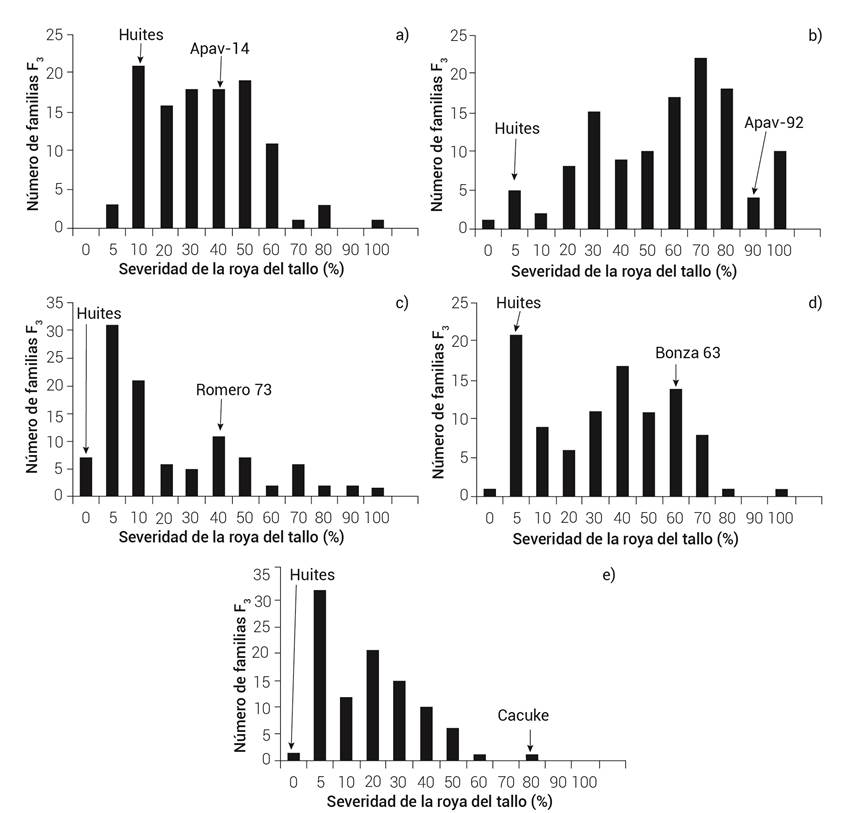
Figura 1 Distribución de familias F3 de las cruzas Huites M95 × Apav-14 (a), Huites M95 × Apav-92 (b), Huites M95 × Romero 73 (c), Huites M95 × Bonza (d) y Huites M95 × Cacuke (e).
El progenitor Apav-92 alcanzó el 100 % de severidad, mientras que los progenitores resistentes Huites M95 y LC-150 mostraron los niveles más bajos de severidad a la raza RTR con 10 y 0 %, respectivamente (Cuadro 2).
Cuadro 2 Porcentaje de infección en hoja bandera de los progenitores a Pgt raza RTR cuando el testigo susceptible alcanzó el 100 % de severidad en Chapingo, México 2013 y genes de planta adulta (APR) de resistencia evaluados mediante la metodología mediante SNPs.
| Progenitor | Respuesta (%) | Gen APR† |
|---|---|---|
| Huites M95 | 10 | Sr58 |
| Apav-92 | 100 | - |
| Apav-14 | 40 | Sr2, Sr58 |
| Romero 73 | 40 | Sr57 |
| Bonza 63 | 70 | Sr58 |
| Cacuke | 80 | Sr57, Sr2, Sr58 |
| LC-150 | 0 | Sr58 |
†APR: adult plant resistance: genes de planta adulta
El análisis por marcadores de Huites M95 y LC-150 indicó, la presencia del gen Sr58 (Cuadro 2), lo cual explica los menores niveles de infección, mientras que Cacuke con tres genes detectados alcanzó infección del 80 % (Cuadro 2). Los resultados indican que en los dos primeros progenitores existen genes no identificados que incrementan la efectividad del Sr58.
Genética de los cruzamientos con el progenitor Huites M95
Las frecuencias observadas y esperadas, el número de genes que condicionan la resistencia y la prueba de X2 en las cruzas se observan en el Cuadro 3. En el cruzamiento de Huites M95×Apav-92 la proporción de 5:103:11 (resistentes: segregantes: susceptibles) indicó que la resistencia de Huites M95 en planta adulta está condicionada por dos genes de efecto aditivo, ya que Apav-92 no posee genes de resistencia. Con las frecuencias observadas en plantas adultas de las cruzas Huites M95×Apav-14 de 20:81:8, Huites M95×Romero 73 de 76:21:1, Huites M95×Bonza 63 de 88:9:1 y Huites M95×Cacuke de 25:73:0, se corrobora que el progenitor resistente Huites M95 posee dos genes de efecto aditivo, mientras que los progenitores Romero 73, Bonza 63 y Cacuke contribuyen con un solo gen de efecto aditivo en sus progenies.
Cuadro 3 Distribución y frecuencias observadas y esperadas de familias F3 de las cruzas con los progenitores Huites y LC-150.
| Cruza | Total fam. F3 | Res O | Res E | Seg O | Seg E | Susc O | Susc E | Núm. genes | X² |
|---|---|---|---|---|---|---|---|---|---|
| Apav-92 × Huites M95 | 119 | 5 | 4.20 | 103 | 86.55 | 11 | 9.24 | 2 | 3.614 |
| Apav-14 × Huites M95 | 109 | 20 | 18.34 | 81 | 74.31 | 8 | 7.33 | 3 | 0.813 |
| Huites M95 × Romero 73 | 98 | 76 | 77.55 | 21 | 21.42 | 1 | 1.02 | 3 | 0.039 |
| Huites M95 × Cacuke | 98 | 25 | 25.51 | 73 | 74.48 | 0 | 0 | 3 | 0.039 |
| Huites M95 × Bonza 63 | 98 | 88 | 89.79 | 9 | 9.18 | 1 | 1.02 | 3 | 0.039 |
| LC-150 × Bonza 63 | 98 | 8 | 8.16 | 81 | 82.65 | 0 | 0 | 3 | 0.036 |
| LC-150 × Cacuke | 98 | 15 | 15.30 | 82 | 83.67 | 1 | 1.02 | 3 | 0.039 |
Res O: resistentes observados; Res E: resistentes esperados; Seg O: segregantes observados; Seg E: segregantes esperados; Susc O: susceptibles observados; Susc E: susceptibles esperados. 2 gl, α = 0.05; X²t = 5.99.
Es posible que los progenitores Huites M95 y Cacuke tengan por lo menos un gen en común, debido a que no se observaron familias homocigóticas susceptibles, además de que al analizar la distribución de frecuencias de los niveles de la enfermedad (Figura 1), las familias tienden a la resistencia, lo que evidencia la presencia de tres genes de efecto aditivo, dos de Huites M95 y uno de Cacuke. Por otra parte, en el análisis genético, el progenitor Huites M95 posee el gen Sr58 y Cacuke los genes Sr2, Sr57 y Sr58 (Cuadro 2). En los cruzamientos de Huites M95 × Romero 73 y Huites M95 × Bonza 63, el nivel de infección de la enfermedad se relacionó con lo obtenido por Harder (1999), quien demostró que la presencia de dos a tres genes de efecto aditivo contribuye a un mayor nivel de resistencia.
De acuerdo con el análisis genético de los progenitores, se detectó la presencia del gen Sr57 en Romero 73 y del gen Sr2 en Cacuke. Los marcadores morfológicos también son importantes para confirmar la presencia de ciertos genes; por ejemplo, el gen Sr57 en la cruza Huites M95 x Romero 73 se detectó por medio de necrosis en la punta de la hoja, lo cual coincide con el estudio de Rouse et al. (2014) quienes afirmaron que el gen Sr57 proviene del progenitor Romero 73 y confiere resistencia a Pgt. El gen Sr2 en la cruza Huites M95 × Cacuke se detectó a través de la presencia de un melanismo en la base de las espiguillas y en los entrenudos de los tallos mejor conocido como Pseudo-black chaff, el cual coincide con lo demostrado por Faris et al. (2008), quienes observaron altos niveles de resistencia a la roya del tallo en las familias derivadas de las cruzas con el progenitor Cacuke.
Genética de los cruzamientos con el progenitor LC-150
En el cruzamiento de LC-150 × Bonza 63 se observó que Bonza 63 es susceptible en campo ya que alcanzó hasta 70 % de severidad; sin embargo, el progenitor LC-150 mostró 0 %. Al analizar las familias se observó que existió una proporción de 8:81:0, con ninguna familia susceptible y con una distribución que tiende a un mayor número de familias segregantes, lo que indica la presencia de dos genes de efecto aditivo provenientes del progenitor LC-150, el cual presenta el gen Sr58 y un gen de Bonza 63; sin embargo, existen familias con niveles de severidad del 0 %, lo cual puede deberse a que tienen un gen en común o a que existe segregación transgresiva.
En LC-150 × Cacuke se presenta una distribución similar a la de la cruza LC-150 x Bonza 63, el cual tiende a la resistencia, y con sólo una familia homocigótica susceptible como el progenitor susceptible, lo que comprueba la presencia de dos genes de LC-150 y uno de Cacuke.
Singh et al. (2000) afirmaron que los programas de mejoramiento genético trabajan continuamente para incorporar resistencia genética a los genotipos sobresalientes contra las royas. Al cruzar una variedad susceptible por una resistente, en su progenie es posible determinar tanto el tipo de acción génica como el número de genes que confiere la resistencia.
Riede et al. (1995) estudiaron la herencia de la resistencia a la roya del tallo en la variedad Waldron, mediante el análisis de la progenie F3 derivada del cruzamiento con otras 13 líneas, de las cuales seis contribuyeron con un gen de resistencia y las otras siete, con dos. Ghazvini et al. (2012) realizaron estudios de resistencia similares al de la presente investigación, con el objeto de identificar nuevas fuentes de resistencia a la raza TTKSK (Ug99) derivadas de la especie silvestre Aegilops triuncialis, donde se detectó la presencia de dos genes dominantes en estudios genéticos en poblaciones F3 inoculadas con Pgt raza MCCF, donde la segregación de las familias fue 8:7:1, lo que confirmó la presencia de dos genes dominantes. Por su parte, Hiebert et al. (2012) estudiaron la genética de la resistencia en plántula a Ug99 en variedades de trigo canadienses Peace y AC Cadillac en poblaciones segregantes F2:3, donde se encontró la presencia de un gen dominante involucrado en la resistencia en ambas poblaciones.
Singh et al. (2001) demostraron que la presencia de más de tres genes de efecto aditivo en royas es deseable en todo programa de fitomejoramiento; en este sentido, la presencia de hasta tres genes de resistencia con efectos aditivos en el germoplasma aquí analizado, es una ventaja per se, ya que podría considerarse como fuente genética para la formación de futuras variedades.
Para acumular más de tres genes aditivos queda la posibilidad de generar el entrecruzamiento o la combinación entre la variedad Huites M95 o la LC-150 con otras variedades que posean cierto grado de resistencia. Además, se ha demostrado que no existen efectos negativos en el rendimiento (Huerta y Singh, 2000; Villaseñor-Espín et al., 2009).
Existen cuatro genes que están asociados a la quemadura de la punta de la hoja, los cuales son: Lr34/Yr18/Sr57/Pmxx/Ltn1; Lr46/Yr29/Sr58/Pmxx/Ltn2; Lr67/Lyr46/Sr55/Pmxx/Ltn3 y Lr68, identificados originalmente para resistencia a roya de la hoja, y años más tarde se comprobó que el mismo gen es efectivo para otros patógenos del trigo como roya amarilla, roya del tallo, mildiu, entre otros, es decir tiene efectos pleiotrópicos. En el análisis génico molecular Huites M95 resultó positivo para el marcador Cslv46 y negativo para los otros marcadores, lo que indica que Huites posee el gen Sr58 de roya del tallo y Ltn2 que ocasiona la quemadura de la punta de la hoja (Herrera-Foessel et al., 2012; Herrera-Foessel et al., 2014; Krattinger et al., 2016; Rinaldo et al., 2017).
Un análisis más detallado de la resistencia a la roya del tallo en poblaciones derivadas de Huites M95 y LC-150 podría proporcionar a los fitomejoradores de trigo información valiosa para el desarrollo de variedades con niveles deseables de resistencia a royas, que incluye a Ug99; sin embargo, esta resistencia debe ser evaluada y confirmada fenotípicamente contra dicha raza de roya del tallo en específico.
Conclusiones
La variedad Huites M95 posee dos genes de efecto aditivo, Sr58 y otro mas no caracterizado, que confieren resistencia en su progenie a la roya del tallo, mientras que Apav-14, Bonza 63, Romero 73 y Cacuke aportan sólo un gen de efecto aditivo al recombinarse con Huites M95. En la genética de LC-150 se presentaron dos genes de efecto aditivo sumados al efecto de un gen de Bonza 63 y Cacuke. El progenitor Cacuke, a pesar de poseer los genes Sr2, Sr57 y Sr58, mostró menor nivel de resistencia. A través de marcadores SNPs no fue posible detectar los genes menores Sr2 y Sr57 que están confiriendo mayor resistencia en progenitores de cruzas de Romero 73 con el progenitor Huites M95; sin embargo, Huites M95 y LC-150 tienen en común el gen Sr58.

