











 nova página do texto(beta)
nova página do texto(beta) Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroduction
Golden Delicious is the most cultivated apple (Malus domestica Borkh.) variety in Chihuahua, México (SAGARPA 2010). Flavor is the main quality attribute of apples from Chihuahua (Bismark, 2002; Olivas et al., 2007). Volatile compounds are essential and confer a complex combination of taste and odor (Defilippi et al., 2009).
More than 300 different volatile compounds have been identified in apples to date (Dixon and Hewett, 2000). The main precursors of volatile compounds in apple are fatty acids which are catabolized through β-oxidation and lipoxygenase (LOX) pathway (Pérez and Sanz, 2008), that produce straight chain aldehydes, alcohols, and esters. Aldehydes are predominant in immature apples (De Pooter et al., 1987), whereas alcohols and esters prevail in ripe fruits (Flath et al., 1967). Alcohol biosynthesis involves enzymes such as alcohol dehydrogenase (ADH; EC 1.1.1.1) and lipoxygenase (LOX; EC 1.13.11.12) (Defilippi et al., 2005; Echeverría et al., 2004a). Availability of alcohols is a limiting factor for ester biosynthesis (Berger and Drawert, 1984; Defilippi et al., 2005), since they are derived from a reaction catalyzed by alcohol acyltransferase (AAT; EC 2.3.1.84) involving esterification of alcohols and acyl-CoA (Sanz et al., 1997). Esters are qualitatively and quantitatively predominant in most apples, accounting for 80 % of the total volatile content in Golden Delicious apples (López et al., 1998).
A small percentage of the fruit is commercialized immediately after harvest, while most of it is stored. Apples stored for a long periods of time are usually kept under controlled atmosphere (CA) conditions (Brackmann et al., 1994). Composition of the atmosphere under CA (1.5 1.7 % O2, 2 2.2 % CO2) differs from cold storage under regular atmosphere (RA) conditions (78.08 % N2, 20.95 % O2, 0.03 % CO2) (Kader, 2002). Recent research has shown that apple storage under CA may suppress production volatile compounds that create the typical aroma (Fellman et al., 2003; Lara et al., 2007; Singh et al., 2010; Starr et al., 2010; Lumpkin et al., 2015).
Fellman et al. (2000) found that Gala apples stored for long periods under a 1 % O2 and 1 % CO2 atmosphere suppressed flavor production. Echeverría et al. (2004b) found that CA (3 % O2 and 2 % CO2) significantly suppressed volatile production after 5 months of storage, compared to apples cold-stored under RA. However, López et al. (2000) found that volatile compound emission in Golden Delicious apples increased after storage for 5 months under a low oxygen atmosphere; sampled apples kept acceptable levels of firmness, acidity, total soluble solids content, color, and high concentrations of branched-chain esters that intensified fruit flavor.
Since flavor depends on volatile biosynthesis, and Golden Delicious apples from Chihuahua, México are primarily recognized by their flavor, this study focused on volatile biosynthesis of fruit stored under CA, RA, and CA after 7 d under RA. Other variables like fatty acids quantification and measurement of activity of the enzymes lipoxygenase (LOX), alcohol acyltransferase (AAT), and alcohol dehydrogenase (ADH) were also determined.
Materials and methods
Plant material and storage conditions
Thirty-five-year old Golden Delicious apple trees from a commercial orchard located in Cuauhtémoc, Chihuahua, México (28° 23' 51.43'' N, 106° 49' 05.79'' W, at 2062 masl) were selected for this study. Apples were harvested 176 days after full bloom, when internal ethylene content (IEC) was 0.9 ppm. Ethylene production was used as a harvest index, according to Dhall (2013). Apples were selected according to color and weight to ensure uniformity in maturity and size, as well as consistent skin-pulp ratio in the analyzed samples. Apples were stored under CA (2 % O2 and 3 % CO2), RA (78.08 % N2, 20.95 % O2, 0.03 % CO2) (Kader, 2002), and CA followed by seven days at RA (CA + RA) at 1 °C. Volatile compound content, specific activity for the enzymes lipoxygenase (LOX), alcohol dehydrogenase (ADH), and alcohol acyltransferase (AAT), and fatty acid composition were evaluated at harvest and after 1, 3, 5, and 7 months of storage.
Aroma volatiles
Volatiles concentration in apples was determined by gas chromatography-mass spectrometry (GC-MS) using the solid phase microextraction (SPME) technique, as described by Maya-Meraz et al. (2014). Apple juice from eight apples per treatment was obtained with a food processor (Turmix, México). The juice (20 mL) was placed in a 20 mL PTFE (polytetrafluoroethylene) vial, frozen in liquid nitrogen, and kept at -70 ºC until analysis. An aliquot of 2 mL of thawed apple juice was placed in a 4 mL vial containing 0.7 g of sodium chloride, and stirred while a SPME fiber (65 μm, PDMS-DVB, Supelco, USA) was exposed to the headspace of the sample for 1 h at room temperature (25 °C ). The fiber was desorbed by splitless injection for 5 min at 200 °C into a GC-MS system (Varian Saturn 2100D GC/MS; California, USA) equipped with an Equity-1 column (60 m × 0.25 mm ID × 0.25μm film thickness; Supelco, USA).
Chromatographic conditions were, initial oven temperature of 33 °C held for 5 min, increased to 50 °C at 2 °C min-1, then increased to 250 ºC at 5 °C min-1, and held for 6.5 min. Helium was used as carrier gas with a flow rate of 1 mL min-1. Mass spectra were obtained by electron impact ionization at 70 eV. Transfer line and ion source temperatures were 250 and 180 °C, respectively. Spectra were recorded with a Saturn GC/MS workstation (Varian).
Volatile organic compounds (VOCs) of interest were identified by spectral match to the National Institute of Standards and Technology (1998), Mass Spectral Library (NIST 98 MS) and by comparison of retention times against high purity standards (ethanol, 2-propanol, 2-methyl 1-propanol, 1-butanol, 2-methyl-1-butanol, 1-pentanol, 3-hexen-1-ol (Z), 2-hexen-1-ol (E), 1-hexanol, 1-heptanol, 1-octanol, 2-ethyl 1-hexanol, acetaldehyde, butanal, 2-methyl butanal, pentanal, hexanal, 2-hexenal, benzaldehyde, octanal, nonanal, decanal, ethyl acetate, 1-methyl ethyl acetate, propyl acetate, 2-methyl propyl acetate, butyl acetate, 2-methyl butyl acetate, pentyl acetate, 2-buten-1-ol, 3-methyl acetate, 3-hexen-1-ol acetate, hexyl acetate, ethyl propanoate, propyl propanoate, butyl propanoate, hexyl propanoate, methyl butanoate, methyl-2methyl butanoate, ethyl butanoate, ethyl-2-methyl butanoate, butyl butanoate, butyl 2-methyl butanoate, hexyl butanoate, hexyl 2-methyl butanoate, ethyl pentanoate, ethyl hexanoate, propyl hexanoate, hexyl hexanoate, and ethyl octanoate) (Sigma-Aldrich and ChemService).
Quantification was accomplished by external standard calibration curves using peak areas. All values represent the average of triplicated samples consisting of eight apples each. Acidity and soluble solids contents were measured in the same juice.
Lipoxygenase specific activity
Peel and cortical tissue from eight apple fruits per treatment were freeze-dried with a Labconco, Freezone 12 (Labconco, Corporation, USA). A 200 mg aliquot of the freeze-dried tissues mentioned above was homogenized three times for 20 s with 5 mL of an extraction solution (0.5 M sodium phosphate buffer pH 6.5, 4 mM dithiothreitol, 1 mM EDTA, 0.2 % Triton X-100, and 1 % polyvinylpolypyrrolidone) using an UltraTurrax T25 homogenizer (IKA Labortechnik, Staufen, Germany). The slurry obtained was filtered through two cheesecloth layers and centrifuged at 25,000 ×g for 15 min. The pellet was discarded, and the supernatant was used as crude extract.
LOX activity was assayed spectrophotometrically at 234 nm and 30 °C by monitoring the formation of conjugated dienes from linoleic acid, according to Wang et al. (2004). The assayed mixture (3 mL) consisted of 2.75 mL sodium phosphate buffer (100 mM, pH 6.5), 50 μL sodium linoleic acid solution (10 mM), and 0.2 mL crude extract. Each determination was done in triplicate, and one activity unit (U) was defined as the increment in one unit of absorbance per minute. Results were expressed as specific activity (U mg-1 of protein) (Wang et al., 2004).
Alcohol acyltransferase specific activity (AAT)
AAT activity was assayed according to the modified Pérez et al. (1996) method. A 10 mg sample of freezedried apple (containing peel and cortical tissue) was homogenized in 1 mL of extraction solution (0.1 M sodium phosphate buffer pH 8.0, 1 mM EDTA, 0.1 % Triton X-100, and 1 % PVPP) utilizing an UltraTurrax T25 homogenizer. The homogenate was centrifuged at 20,800 ×g for 20 min at 4 ºC. The supernatant was recovered and set on ice as crude enzyme extract. AAT activity was assayed by mixing 2.5 mL MgCl2 solution (5 mM MgCl2 in 0.1 M sodium phosphate buffer pH 8.0), 150 μL of acetyl-CoA solution (2.5 mM acetyl-CoA in 0.1 M sodium phosphate buffer pH 8.0), 50 μL butanol solution (200 mM butanol in 0.1 M sodium phosphate buffer pH 8.0), and 200 μL crude extract. The mixture was incubated at 35 ºC for 15 min. After that 100 μL of 10 mM 5,5-dithiobis (nitrobenzoic acid) (DTNB) were added, and the mixture allowed to stand at room temperature for 10 min.
AAT activity was measured spectrophotometrically by the increment in absorbance at 412 nm as a yellow thiophenol complex of DTNB and free Coenzyme A(CoA) liberated during the catalytic reaction formed. Each determination was carried out in triplicate: one activity unit (U) was defined as the increment in one absorbance unit at 412 nm per minute. Results were expressed as AAT specific activity (mUmg-1 of protein) (Echeverría et al., 2004a).
Alcohol dehydrogenase specific activity (ADH)
The method used for extraction of ADH was described by Chang et al. (1982). A 100 mg freeze-dried apple (containing peel and cortical tissue) sample was homogenized three times for 20 s with 5 mL extraction solution (10 mM sodium phosphate buffer pH 8.0, 5 mM dithiothreitol and 0.5 % polyvinylpolypyrrolidone). The homogenate was centrifuged at 15,000 ×g for 15 min at 4 ºC. The pellet was discarded, and the supernatant was used as crude extract. The reduction of acetaldehyde was followed spectrophotometrically at 25 ºC by measuring the change in absorbance at 340 nm for 2 min of a reaction mixture containing 800 mL of a 25 mM MES (2-(N-morpholino) ethanesulfonic acid) buffer at pH 7.2, 50 mL of nicotinamide adenine dinucleotide (NADH) (5 mM), 100 mL of enzyme extract, and 50 mL of acetaldehyde (80 mM). Each determination was done in triplicate; one activity unit (U) was defined as the decrease in one unit of absorbance at 340 nm per minute, and results were expressed as specific activity (U mg-1 protein).
Fatty acid analysis
Fatty acids content was determined by fatty acid methyl ester (FAME) analysis according to slight modifications to Defilippi et al. (2005). A 0.15 g sample of freeze-dried apple (containing peel and cortical tissue) was mixed with 1 mL of toluene and shaken overnight (100 rpm) at room temperature on an orbital shaker. Subsequently, 500 μL of methanolic 0.5 N sodium methoxide was added and shaken for 1 h (100 rpm) at room temperature. After the hour, the transesterification reaction was terminated by addition of 50 μL of a 10 % NaCl solution. Finally, 400 μL of heptane was added, mixed and centrifuged for 5 min at 1400 ×g. After phase separation, an aliquot of the upper phase was transferred to a vial and refrigerated for further analysis.
Fatty acid composition was determined by gas chromatography using a 7820 Agilent instrument equipped with a flame-ionization detector. The instrument was fitted with a DB-Wax capillary column (30 m, 0.25 mm I. D. 0.25 μm). Injector port and detector temperatures were 250 and 300 °C, respectively. Oven temperature was initially set at 50 ºC, increased to 200 ºC at a rate of 25 ºC min-1, increased to 230 ºC at a rate of 3 ºC min-1 and held in that condition for 4 min. The carrier gas was helium with a flow rate of 1 mL min-1. Identification of FAMEs was done by comparing the retention times to those of high purity standards analyzed under identical chromatographic conditions. Each determination was done in triplicate.
Statistical analysis
For statistical analysis, a mixed model design was used with storage conditions, storage period, and replication as fixed factors, and replications nested in treatments as random factors. Analyses were carried out using SAS Version 9 (SAS Institute, Cary, NC). Means were separated by (Least Squares) LS means test at P ≤ 0.05. Volatile compounds data collected were analyzed by PROC MIXED for the analysis of a repeated measures factorial ANOVA and the statistical comparison of means was Tukey's range test method.
Results and discussion
Volatile compounds
In this study, 35 volatile compounds (VC) were identified and quantified during storage of Golden Delicious apples. The compounds identified included 18 esters: ethyl acetate, n-propyl acetate, 2-methyl propyl acetate, butyl acetate, 2-methyl butyl acetate, ethyl pentanoate, butyl propanoate, pentyl acetate, butyl butanoate, 3-hexen-1-ol acetate, hexyl acetate, butyl 2-methyl butanoate, propyl hexanoate, hexyl propanoate, hexyl butyrate, ethyl octanoate, hexyl 2-methyl butyrate, hexyl hexanoate; seven aldehydes: butanal, butanal 2-methyl, pentanal, cis 3-hexenal, hexanal, 2-hexenal, nonanal; and ten alcohols: 1-butanol, 2 methyl 1-propanol, 2 methyl 1-butanol, 1-pentanol, 3-hexen-1-ol (Z), 2-hexen-1-ol (E), 1-hexanol, 1-heptanol, 2-ethyl 1-hexanol, 1-octanol. Storage under CA caused a decrease on VC development (Figure 1). Mattheis et al. (1995), Fellman et al. (2003) and Saquet et al. (2003) found similar results for Bisbee Delicious, Redchief Delicious and Jonagold apples, respectively.
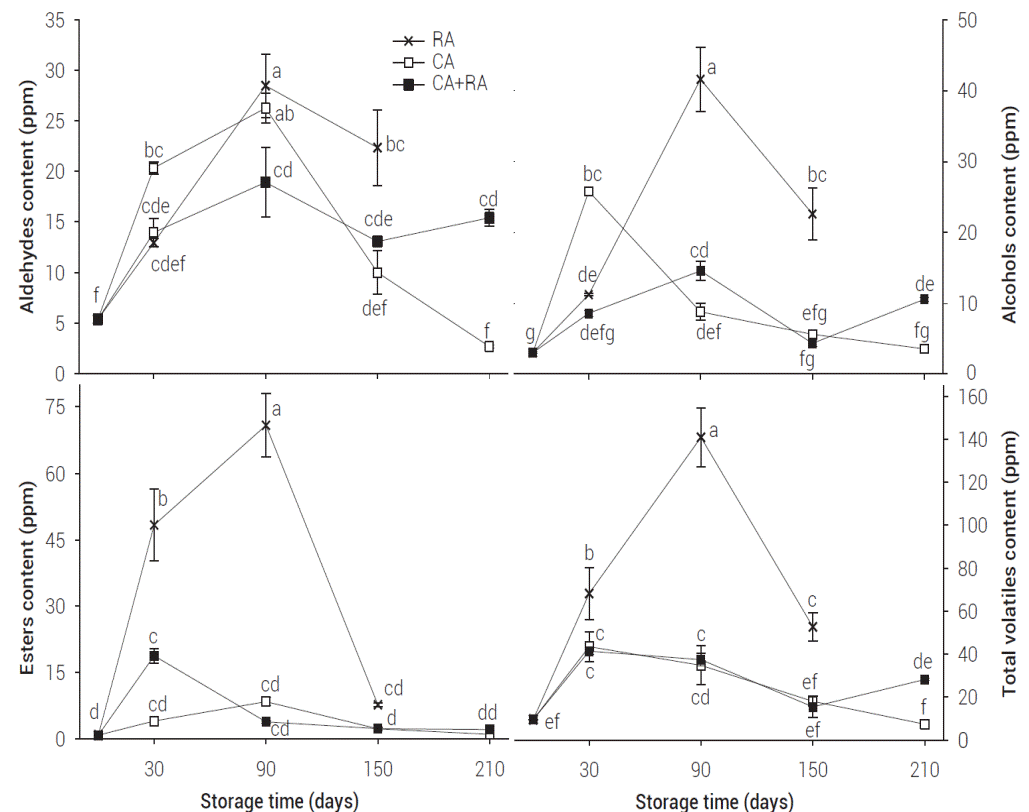
Figure 1: Aldehyde, alcohol, ester and total volatile content in Golden Delicious apple fruits under different storage conditions. Values represent mean of three repetitions. Vertical bars represent ± SE. Means showing different letters are significantly different (Tukey, 0.05).
The main VC found in Golden Delicious apples at harvest time were (in decreasing order) 2-hexenal, 2-methyl 1-butanol, hexanal, butyl acetate, 2-methyl 1-propanol, 2-methyl butyl acetate, cis 3-hexenal, and hexyl acetate (Table 1). Significant interaction between different atmospheric conditions and storage time was detected. CA and CA + RA apples showed their highest total VC values after one month of storage, with no significant differences among CA, CA + RA, and RA (P < 0.05) (Figure 1). However, although total VC values were similar among treatments after one month of storage, their specific composition was different for each treatment (Figure 1, Table 1).
Table 1: Production of volatiles compounds (ppm) by Golden Delicious apples under different storage conditions.
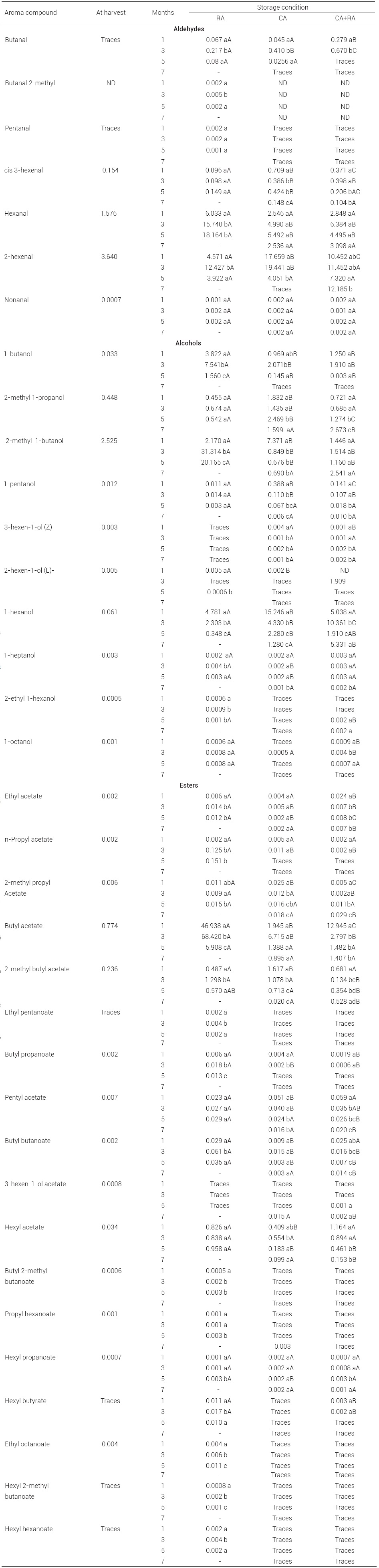
Values are mean of three repetitions. Means within the same storage period followed by different capital letters are significantly different at P ≤ 0.05 (LS means test). Means within the same storage conditions followed by different small letters are significantly different at P ≤ 0.05 (LS means test). Traces are values below 0.0005 ppm. ND, not detected.
Treatment RA presented considerably higher concentration of esters (mainly butyl acetate) when compared to CA + RA and CA apples, after one month of storage (P < 0.05). CA + RA treatment induced 78 % higher ester values than CA-treated apples, essentially butyl acetate, after one month of storage. On the other hand, CA-treated apples had 27.3 % higher aldehyde concentration and 58 % more alcohol levels than RA and CA + RA apples (mainly hexanol and 2-metyl-1-butanol) after one month of storage (Figure 1, Table 1).
Butyl acetate (66.7 %, 46.9 ppm), hexanal (8.6 %, 6.0 ppm), and 2-hexenal (6.5 %, 4.5 ppm) make up about 82 % of the total VC produced by apples, after one month of RA storage. Compounds 2-hexenal (34.7 %, 17.7 ppm), 1-hexanol (30 %, 15.3 ppm) and 2-methyl 1-butanol (14.5 %, 7.4 ppm) account for 80 % of the total VC by CA stored apples after one-month storage. Butyl acetate (34.5 %, 13 ppm), 2-hexenal (27.9 %, 10.5 ppm), and 1-hexanol (13.5 %, 5 ppm) amount to about 76 % of total VC by CA + RA stored apples, after month one of storage.
CA conditions inhibited the production of butyl acetate and hexyl acetate and increased hexanol concentration, after one month of storage. At this moment, branched ester, 2-methyl butyl acetate, increased on CA apples. According to López et al. (1998) branch chain esters are not affected by CA storage, since these come from the amino acid pathway. Fellman et al. (1993) found greater concentration of 2-methyl butyl acetate on apples stored under CA conditions, when compared to RA apples.
Seven days of RA after CA caused a regeneration of volatiles to get a composition resembling RA apples at month one of storage: ester biosynthesis was present, mainly butyl acetate, and concentration of alcohols and aldehydes (mainly 1-hexanol and 2-hexenal) decreased (Figure 1, Table 1). These results have important sensory implications: studies on commercial apple odor have correlated 'unwanted essences' with high levels of alcohols like hexanol which give an earthy unpleasant flavor, and 'desirable essences' with high levels of hexanal, 2-hexenal, and butyl acetate (Dürr and Schobinger, 1981; Petró-Turza et al., 1986). Altisent et al. (2011) found that the emission of 26 volatile compounds increased on Golden Reinders apples, after a regeneration period (air storage) of 2 and 4 weeks after ultralow-oxygen storage. According to Dixon and Hewett (2000), after hypoxia apples increase ester concentration. Young et al. (2004) indicated that low molecular weight esters increase more rapidly than their counterparts.
RA apples showed a four-fold value on total VC when compared to CA and CA + RA apples after three months of storage (Figure 1) (P < 0.05); however, by the third month of RA storage, apples had the highest ester content (70.8 ppm, mainly butyl acetate), but CA and CA + RA apples showed 88 % lower ester values, with no significant differences among them (Figure 1) (P < 0.05).
An increase on aldehydes content was observed in all treatments, without statistical differences between CA and RA, for the three-month storage. However, aldehyde composition among CA and RA apples varied: hexanal dominated RA-treated apples, while 2-hexenal prevailed in CA apples (Table 1). A 79 % increase in alcohol content was found in RA apples compared to CA apples, mainly 2-metyl-1-butanol, 1-butanol and 1-hexanol, after the same storage period. For the same period, a higher concentration of alcohols was observed on CA + RA apples when compared to CA apples, mainly due to hexanol production (Figure 1, Table 1).
At the third month of storage, RA apples showed the highest content of total VC's, mostly composed by by butyl acetate (68 ppm, 48.5 %), 2-methyl-1-butanol (33.1 ppm, 22.2 %), hexanal (15.7 ppm, 11.2 %) and 2-hexenal (12.4 ppm, 8 %). In contrast, CA-stored apples had a VC profile made up of 2-hexenal (19.4 ppm, 45.8 %), butyl acetate (15.8 %, 6.7 ppm), hexanal (11.8 %, 5.0 ppm) and 1-hexanol (4.3 ppm, 10 %). CA + RA apples showed a VC profile with 2-hexenal (11.4 ppm, 29.2 %), 1-hexanol (10.4 ppm, 26.4 %) and hexanal (6.4 ppm, 16.2 %). Butyl acetate ester accounted for almost 70 % of total VC in RA apples, and 2-hexenal, an aldehyde, accounted for almost 35 % of total VC content in CA apples.
This behavior demonstrates the delaying ripening effect in CA-stored apples, since aldehydes are precursors of alcohols, and in turn, alcohols are ester precursors. Butyl acetate ester was the main compound produced under RA (66.7 %) and under CA + RA apples (34.5 %), which agrees with Drawert (1973), who found this compound to be the prevailing ester in Golden Delicious apples under RA-stored conditions. The higher ester concentration on RA apples (mainly butyl acetate) modifies sharply aroma, compared to CA apples, since butyl acetate is an impact compound in Golden Delicious apple (Kakiuchi et al., 1986), while hexanal and 2-hexenal are aldehydes related to 'unripe' flavors in Golden Delicious apples (Flath et al., 1967; Rizzolo et al., 1989).
After five months of storage, total VC content decreased sharply on RA apples, being 74 % lower than after three months of storage, although still considerably higher than in CA and CA + RA apples. A significant decrease in the concentration of aldehydes on CA apples was observed (55 % lower values than on RA conditions), mainly due to a major decrease in 2-hexenal. CA + RA apples showed higher aldehyde concentration when compared to CA after five months of storage (Figure 1) (P < 0.05). A considerable decrease in ester concentration was found in RA apples on the fifth month of storage, an 89 % lower ester content than at three months of storage. No significant difference on esters profiles was found among RA, CA, and CA + RA apples (Figure 1).
At the fifth month of storage, the main VC's on Golden Delicious apples under RA were 2-methyl-1-butanol (38 %, 20.2 ppm), hexanal (34 %, 18.2 ppm) and butyl acetate (11 %, 6.0 ppm), while the main VC occurring on CA apples were hexanal (31 %, 5.5 ppm), 2-hexenal (23 %, 4.0 ppm), and 2-methyl 1-propanol (14 %, 2.5 ppm). On CA + RA apples the compounds 2-hexenal (39 %, 7.3 ppm), hexanal (24 %, 4.5 ppm), and 1-hexanol (10 %, 1.9 ppm) were the major VC produced after five months of storage.
Finally, after seven months of storage, CA + RA apples showed a volatile ́s recovery, presenting 82.5 % more aldehydes and 66 % more alcohol concentration than CA apples (Figure 1). The main VC produced on CA+RA apples were 2-hexenal (43 %, 12 ppm), 1-hexanol (19 %, 5.3 ppm) and 2-methyl-1-propanol (10 %, 2.7 ppm), while in CA apples the major VC were hexanal (35 %, 2.5 ppm), 2-methyl 1-propanol (22 %, 1.6 ppm) and hexanol (17 %, 1.3 ppm). RA apples were not evaluated at seven months of storage, since fruit did not maintain the required quality.
LOX, AAT, and ADH activity
Lipoxygenase (LOX) may play a key role in determining the composition of volatile compounds in apple (Fellman et al., 2000). In this study an increase in LOX specific activity was observed from harvest to the first month of storage (Figure 2). No significant difference in LOX activity was found among RA and CA apples at one month of storage.
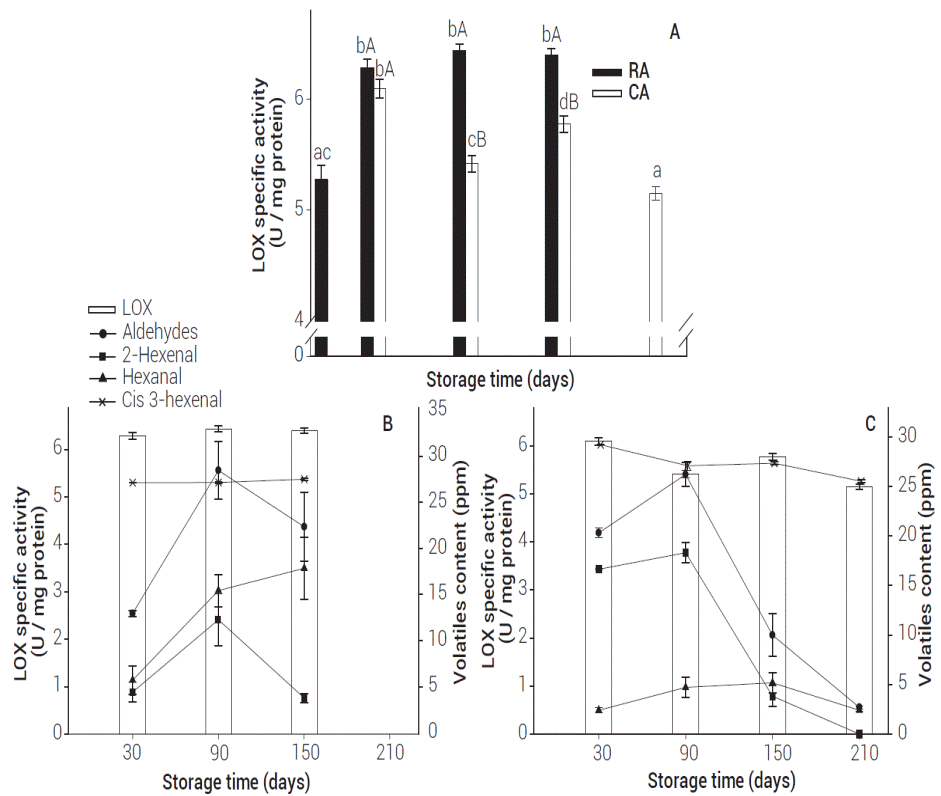
Figure 2: LOX specific activity in Golden Delicious apples at different storage conditions (A), LOX specific activity at RA (B) and at CA (C) compared to important aldehydes. Values represent mean of three repetitions. Vertical bars represent ± SE. Means within the same storage period followed by different capital letters are significantly different at P ≤ 0.05 (LS means test). Means within the same storage conditions followed by different small letters are significantly different at P ≤ 0.05 (LS means test).
After the first month, storing fruit under CA caused a decrease in the specific activity of the enzyme (Figure 2). Lara et al. (2007) found similar results attributing this effect to LOX-O2 requirements. Figure 2 shows the relationship between LOX specific activity and total aldehydes production during apple storage under RA and CA conditions. Determination coefficient (r2) between aldehydes and LOX activity was 0.25 for RA apples and 0.85 for CA apples. These details, along with decreased LOX specific activity observed on CA apples, could indicate that LOX activity plays an important role in controlling aldehyde production when oxygen concentration is limited.
Ester-like volatile compounds are generated by the esterification of alcohols and acyl-CoA catalyzed by the enzyme alcohol acyltransferase (AAT) (Sanz et al., 1997). The effect of storage conditions on the AAT specific activity is shown in Figure 3. The highest AAT activity was observed at harvest (208.26 mU mg-1 protein). After the first month of storage, higher enzyme specific activity was found in RA stored fruit when compared to CA conditions (P < 0.05). Fellman et al. (2000) noted this same effect in Law Rome and 262 Rome apples. On RA apples, as the storage period advanced AAT specific activity decreased, suggesting enzymatic activity was affected by storage conditions.
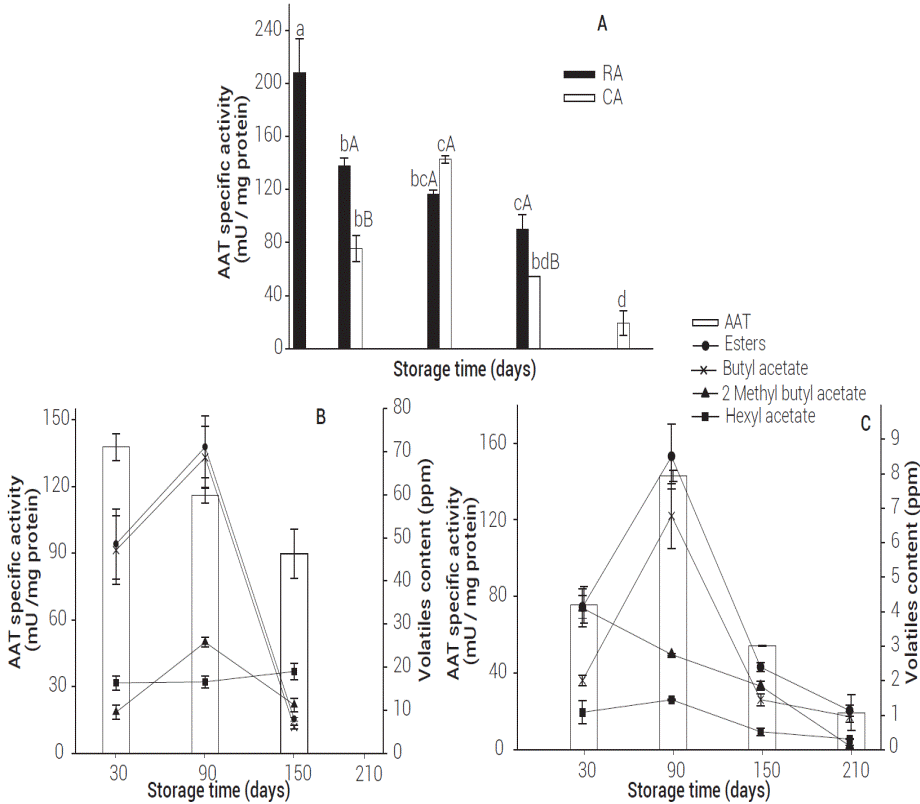
Figure 3: AAT specific activity in Golden Delicious apples at different storage conditions (A), AAT specific activity at RA (B), and at CA (C) compared to important alcohols and esters. Values represent mean of three repetitions. Vertical bars represent ± SE. Means within the same storage period followed by different capital letters are significantly different at P ≤ 0.05 (LS means test). Means within the same storage conditions followed by different small letters are significantly different at P ≤ 0.05 (LS means test).
After three months of storage there was an increase in AAT specific activity of CA apples that was not significantly different from RA apples (P < 0.05). Lara et al. (2007) found higher AAT specific activity in fruit stored under CA than in RA stored fruit. The highest AAT specific activity for CA apples occurred during the third month of storage, which coincides with their highest ester production. A decrease in AAT specific activity was observed in both treatments (RA and CA) after five months of storage; RA showed significantly higher AAT activity than CA (P < 0.05).
Figure 3 shows the relationship between AAT activity and esters (total esters, butyl acetate, 2 methyl butyl acetate, and hexyl acetate) on RA and CA stored apples. RAstored apples did not show a clear connection between AAT specific activity and the ester synthesis. However, CA apples showed high correlation between AAT specific activity and total esters (r2 = 0.92). These findings could indicate that under CA conditions, ester production is correlated with AAT enzyme activity. In the case of RA apples, ester production did not show to be dependent on AAT enzyme activity. Echeverría et al. (2004a) and Villatoro et al. (2008) noted that modifications in AAT specific activity could not explain the observed behavior in the production of esters.
Figure 4 shows ADH specific activity in apples stored under CA and RA (P < 0.05). CA apples showed higher ADH activity during storage, when compared to RA apples (P < 0.05). The highest ADH activity in CA stored apples was observed after three months of storage, having values four-fold higher than RA apples. Higher ADH activity in the CA-stored apples coincided with a lower VC production; it is suggested that this enzyme activity may be related to the onset of fermentative processes after extended storage under hypoxia. Furthermore, AAT activity after extended storage under CA conditions decreased (Figure 3), thus preventing esterification of alcohol precursors (Lara et al., 2007).
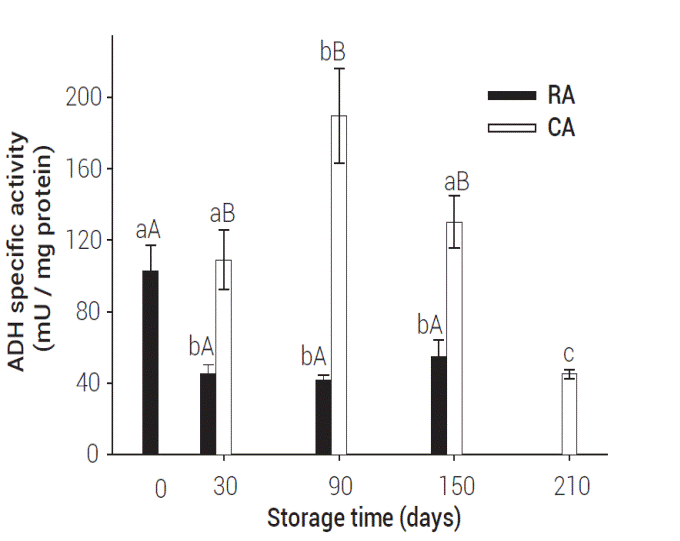
Figure 4: ADH specific activity in Golden Delicious apples at different storage conditions. Values represent mean of three repetitions. Vertical bars represent ± SE. Means within the same storage period followed by different capital letters are significantly different at P ≤ 0.05 (LS means test). Means within the same storage conditions followed by different small letters are significantly different at P ≤ 0.05 (LS means test).
Fatty acids
Fatty acids are considered the main precursors of volatile compounds; they are important structural compounds and metabolic constituents of fruit cells (Marangoni et al., 1996). They are detached and metabolized by lipase, β-oxidation enzymes and/or lipoxygenase, and they produce volatile aroma substances (Fellman et al., 2000). Figure 5 shows fatty acids obtained from RA and CA apples. Linoleic acid was found in higher concentration (85 - 125 ppm), followed by palmitic acid (27 - 45 ppm), linolenic acid (5 - 17 ppm), oleic acid (3 - 12 ppm) and stearic acid (5 - 8 ppm).
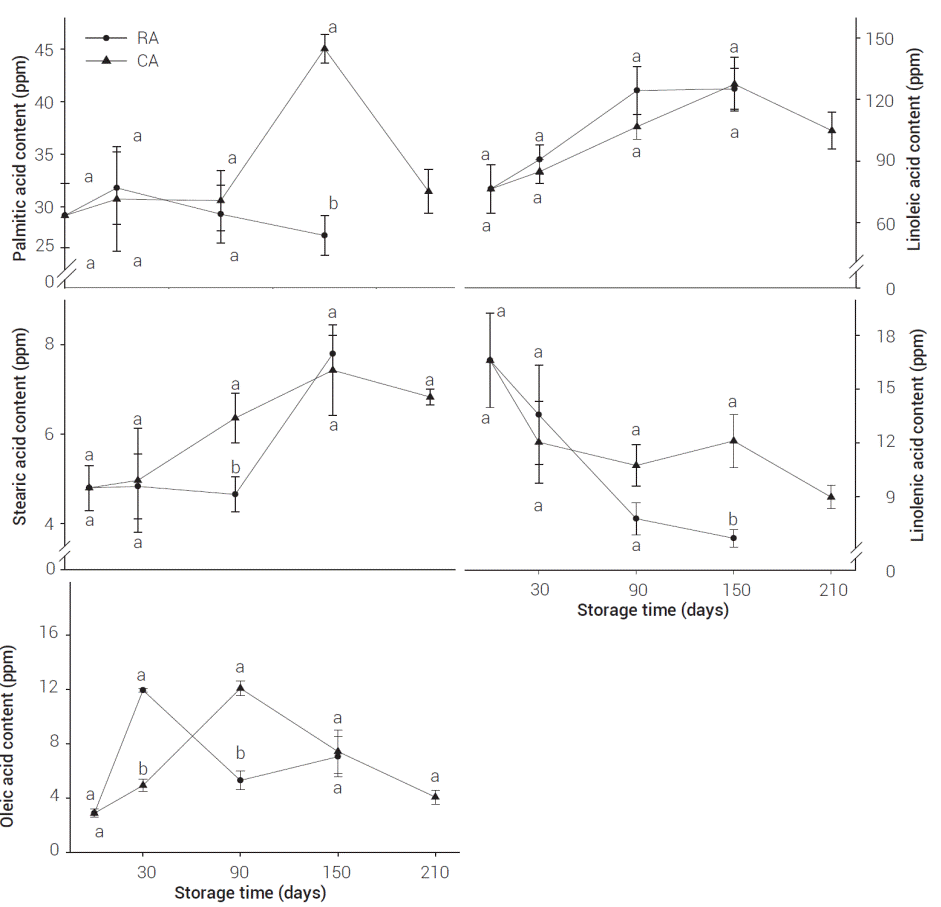
Figure 5: Fatty acids in Golden Delicious apples at different storage conditions. Values represent mean of three repetitions. Verticals bars represent ± SE. Means showing different letters are significantly different at P ≤ 0.05 (LS means test).
Linoleic acid is considered one of the main precursors of volatile compounds in apples (Yahia, 1994). Linoleic acid concentration increased during storage in both treatments, but no significant differences were found between RA and CA (P < 0.05). Linoleic acid concentration, as well as the VC of the fruit stored under RA-conditions, increased continuously until the fifth month of storage, which is associated with fruit climacteric ripening (Song and Bangerth, 1996; Song and Bangerth, 2003; Defilippi et al., 2005).
Palmitic acid concentration increased on CA apples after five months of storage. Palmitic acid concentration remained steady on RA apples through storage (P < 0.05). Similar results were shown by Song et al. (2003) who found higher concentration of palmitic acid on CA apples after 6 months of storage. Linolenic acid concentration decreased thorough storage, more markedly in RA apples; this was the only fatty acid that decreased in concentration steadily during storage. Previous research found that linolenic acid in apples decreases with ripening, as a result of breakdown of fatty acids in chloroplasts to produce straight C-chain esters (Meigh and Hulme, 1965; Galliard, 1968). Oleic acid concentration increased after one month of storage for RA and after three months of storage in CA apples. Stearic acid content steadily increased during storage, and it reached its highest value after five months of storage. CA apples reached higher values of stearic acid after three months of storage, when compared to RA apples. Stearic acid in Jonagold apples showed similar behavior under CA conditions (Saquet et al., 2003).
No significant decrease of the fatty acid content in CAstored apples with respect to that observed in RA-stored apples was observed in this study, except for oleic acid during the first month of storage. These results differed with those found by Saquet et al. (2003), who reported that fatty acid concentrations in Jonagold apple pericarp tissue are lowered under CA storage. This suppression effect on fatty acid concentration in CA apples may be dependent upon atmospheric composition and apple variety under study, among other conditions.
Conclusions
Storage conditions affected composition and concentration of volatile compounds in apples. Volatile compounds synthesis decreased under CA storage. This storage conditions inhibited the production of esters butyl acetate and hexyl acetate, two key apple aroma compounds. We propose that the main cause of this reduction is the decrease in the activity of LOX and AAT enzymes at some stage during CA storage.

