










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista fitotecnia mexicana
versión impresa ISSN 0187-7380
Rev. fitotec. mex vol.36 no.2 Chapingo abr./jun. 2013
Artículos científicos
Parámetros genéticos de Pinus patula en un ensayo de progenies establecido en dos altitudes
Pinus patula genetic parameters in a progeny test at two altitudes
Erick Morales González1, 3, Javier López Upton1*, J. Jesús Vargas Hernández1, Carlos Ramírez Herrera1 y Abel Gil Muñoz2
1 Programa Forestal, Campus Montecillo, Colegio de Postgraduados. km 36.5 carretera México-Texcoco. 56230, Montecillo, Texcoco, Edo. de México. Tel: (52) (595) 9520200 Ext. 1463. *Autor para correspondencia (jlopezupton@gmail.com)
2 Campus Puebla, Colegio de Postgraduados. km 125.5 Carretera Federal México-Puebla. 72760, Santiago Momoxpan, San Pedro Cholula, Puebla. Tel: (52) (222)-2851442.
3 Dirección Actual: Sitio Experimental La Campana, Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias. km 33.3 Carretera Chihuahua-Ojinaga. 32910, Cd. Aldama, Chihuahua. Tel: (614) 4510601.
Recibido: 14 de Noviembre del 2011
Aceptado: 13 de Febrero del 2013
Resumen
En Aquixtla, Puebla se inició en 2001 un programa de mejoramiento con Pinus patula Schiede ex Schltdl. et Cham., con el objetivo de incrementar el volumen del fuste y la adaptabilidad. En julio de 2005 se estableció un ensayo de progenies en dos sitios ubicados a altitudes diferentes (S1 = 2660 m y S2 = 2980 m). En cada ensayo se incluyeron 84 familias de medios hermanos en un diseño de bloques incompletos al azar, con 20 repeticiones; la unidad experimental fue una planta. Se evaluó la altura (ALT), diámetro del fuste a 30 cm y a 1.3 m (D030 y DN, respectivamente), número de verticilos (NV), volumen (VOL) y supervivencia. A cuatro años de edad únicamente se midió la ALT, D030 y NV, y a cinco años de edad se evaluaron todas las características. En S1 las heredabilidades individuales (h2i) a cuatro años de ALT, D030 y NV, y a cinco años de ALT, D030, DN, Nv y VOL, fueron 0.16, 0.08, 0.18, 0.18, 0.09, 0.15, 0.16 y 0.16, respectivamente. Las medias por variable fueron: 2.86 m, 5.62 cm, 8.2, 3.85 m, 7.44 cm, 4.53 cm, 10.8 y 4.43 dm3, respectivamente. La supervivencia promedio en este ensayo fue 83 % (entre familias varió entre 57 y 100 %); 1.1 % de árboles murieron por frío (0 a 14.3 % entre familias). En S2 las h2i encontradas para las variables antes mencionadas y en el mismo orden fueron: 0.07, 0.04, 0.03, 0.13, 0.09, 0.12, 0.12, y 0.09; así mismo, las medias por variable fueron: 1.45 m, 3.05 cm, 6.3, 2.19 m, 4.97 cm, 2.13 cm, 8.4 y 0.73 dm3, respectivamente. La supervivencia promedio en S2 fue 66 % (entre familias, de 37.5 % a 100 %); 6.2 % de árboles murieron por frío (0 a 26.7 % entre familias).
Palabras clave: Pinus patula, adaptación, ensayo de progenies, mejoramiento genético forestal.
Abstract
In 2001, a Pinus patula Schiede ex Schltdl. et Cham improvement program was in Aquixtla, Puebla an improvement program with and aimed to increase bole volume and adaptation. In july 2005, a progeny test was established in two sites located at different elevation (S1 = 2660 m and S2 = 2980 m). Each trial included 84 half-sib families in an incomplete block design with 19 repetitions at random, with single tree plots. Plant height (HT), stem diameter at 30 cm (D030) and 1.3 m (DBH), whorls number (WN), volume (VOL) and survival were recorded. At 4 years, old only HT, D030 and NW were recorded, while at 5 years all traits were analized. Individual heritabilities (h2i) in S1 for HT, D030 and NW at 4 years, and HT, D030, DBH, NW and VOL at 5 years were 0.16, 0.08, 0.18, 0.18, 0.09, 0.15, 0.16 and 0.16, respectively. Mean values for those variables were: 2.86 m, 5.62 cm, 8.2, 3.85 m, 7.44 cm, 4.53 cm, 10.8 and 4.43 dm3, respectively. Mean survival value in this study was 83 % (among families, 57 to 100 %), 1.1 % of trees appeared to be killed by cold (0 to 14.3 % among families). In S2 the h2i found for the variables above mentioned and in the same order were 0.07, 0.04, 0.03, 0.13, 0.09, 0.12, 0.12, and 0.09; likewise, the means obtained for each variable were 1.45 m, 3.05 cm, 6.3, 2.19 m, 4.97 cm, 2.13 cm, 8.4 and 0.73 dm3, respectively. Average survival in S2 was 66 % (among families, 37.5 % to 100 %), 6.2 % of trees were killed by cold (0 to 26.7 %, among families).
Key words: Pinus patula, adaptation, progeny test, genetic improvement.
INTRODUCCIÓN
Para el establecimiento de plantaciones forestales comerciales y de restauración es necesario disponer de un abasto suficiente de semilla del germoplasma con la calidad genética superior. La opción más adecuada es tener huertos semilleros con árboles mejorados genéticamente. Para establecer tales huertos es necesario emplear semilla de individuos superiores, seleccionados con base en características fenotípicas de importancia económica (White et al., 2007), cuyos fenotipos sean atribuibles mayormente a su constitución genética, por lo que es necesario determinar la calidad genética de las plantas derivadas de los árboles seleccionados.
Una forma de determinar la calidad genética es a través de la evaluación del crecimiento de la progenie, de preferencia en los terrenos donde se establecerán las plantaciones (Clausen, 1990). Un ensayo de progenies es una evaluación de los progenitores con base en la respuesta de su descendencia, y sirve para determinar el valor genético de los árboles inicialmente seleccionados, así como otros parámetros genéticos (Wright, 1964). Sin embargo, los ensayos de progenies son caros y su costo aumenta conforme avanza la edad de los individuos, por lo que se practica la selección temprana (White et al., 2007), a una edad anticipada en la que se evalúa una característica de manifestación precoz fuertemente correlacionada con un carácter de interés económico a la edad madura (Wu, 1998; Vargas-Hernández y Adams, 2002).
Las evaluaciones a edad temprana en especies de pino han sido poco estudiadas en México (Sáenz-Romero et al., 1994; Farfán et al., 2002), y la mejora de los pinos mexicanos ha sido realizada en otras regiones del mundo (Nyoka et al., 1994; Dvorak et al., 2000). Pinus patula Schiede ex Schlechtendal et Chamisso es una especie exclusiva y valiosa en México por su velocidad de crecimiento y calidad de madera (Velázquez et al., 2004). Esta especie puede crecer más de 1 m de altura por año (Vela, 1980; Monroy, 1995); es una de las especies subtropicales más plantadas en el mundo para producción de madera en rollo y material para celulosa (Gillespie, 1992; Dvorak et al., 2000); y en países de África, Asia y Sudamérica así como en Nueva Zelanda y Australia ha mostrado incrementos de volumen de 15 m3 ha-1 a 25 m3 ha-1, y en casos excepcionales hasta 36 m3 ha-1 (Wormald, 1975).
Dada la importancia comercial y ecológica de esta especie en el Estado de Puebla, productores forestales de Aquixtla, Pue. iniciaron un programa de mejoramiento genético con P. patula, con el propósito de aumentar su productividad pero sin afectar su adaptación, en terrenos que se ubican entre 2630 m a 3170 m de altitud. Por tanto, el objetivo del presente estudio fue determinar la respuesta de las familias de medios hermanos establecidas en el ensayo de progenies a dos altitudes, así como estimar parámetros genéticos útiles para cumplir con el objetivo del programa de mejoramiento genético forestal.
MATERIALES Y MÉTODOS
Material biológico
En la etapa inicial se seleccionaron fenotípicamente árboles de buen porte y volumen mayor que sus vecinos, en una propiedad de aproximadamente 250 ha ubicada en el municipio de Aquixtla, Puebla, denominada Conjunto Predial, en un intervalo altitudinal que oscila entre 2630 m y 3170 m, donde se recolectó semilla para establecer un ensayo de progenies a dos elevaciones (2660 m y 2980 m). El germoplasma se recolectó de 84 árboles (familias de medios hermanos) selectos de Pinus patula var. patula, que es la variedad "típica", natural de esta zona del centro del país.
Los ensayos de progenies se establecieron en julio del 2005 con plántulas de seis meses de edad, en parajes de Chichicaxtla [19° 44' 10" LN y 97° 58' 57" LO, a una altitud de 2660 m, con exposición N, temperatura media anual de 12.6 °C (Crookston, 2008)], y en Sierra Mojada [19° 42' 46" LN y 97° 59' 21" LO, a 2980 m, con exposición prácticamente zenital, temperatura media anual de 11.1 °C (Crookston, 2008)]. En los dos sitios la plantación se hizo a un espaciamiento de 2.5 m x 2.5 m, en un diseño de bloques incompletos al azar con 19 repeticiones, con un árbol por familia como unidad experimental (parcela). Debido a diferencias en el número de plantas germinadas y supervivientes de cada familia, los últimos bloques en cada ensayo no incluyeron a todas las familias seleccionadas, por lo que para el análisis se estimó la media armónica del número de plantas por familia en cada ensayo. Simultáneamente se plantaron dos hileras de árboles a iguales espaciamientos como una franja de protección, para propiciar competencia completa de las plantas en los ensayos.
Variables evaluadas y análisis estadístico
En los dos ensayos las variables se midieron durante los inviernos de 2009 y 2010, en la temporada del año en que los árboles detienen su crecimiento (Greenwood, 1977), cuando los árboles tenían cuatro y cinco años de edad, respectivamente, a partir de su siembra en vivero. Se midió la altura total del árbol (ALT) en cm, el diámetro del tronco a 30 cm (D030) del suelo y a 1.30 m (diámetro normal, DN) con aproximación a mm. El punto de medición en el tronco en 2009 se marcó para asegurar que la medición del año siguiente fuera en el mismo punto. Se registró el número de verticilos (NV) en cada árbol, y se evaluó la supervivencia como una característica binaria, 0 (muerto) y 1 (vivo), para cuantificar plantas muertas ocasionadas por ambientes adversos, como heladas, sequías y plagas, en los años de medición (evaluaciones anuales desde su plantación). Se calculó el volumen del fuste con corteza mediante la ecuación para árboles jóvenes de P. patula (Carrillo et al., 2004):
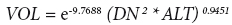
donde: ALT = altura total del árbol (m); y DN = diámetro normal a 1.30 m (cm).
En el ensayo establecido a mayor elevación (Sierra Mojada), debido al menor crecimiento de los árboles, durante el año 2009 no se pudo medir DN, y consecuentemente tampoco se calculó el volumen, pero sí en el año 2010. Todas las variables se analizaron por sitio, con el modelo lineal siguiente: Yij = μ + Bi +Fj +eij, donde: Yij = valor observado de la j-ésima familia en el ij-ésimo bloque (repetición); μ = media poblacional; Bi = efecto fijo del i-ésimo bloque; Fj = efecto aleatorio de la j-ésima familia ~ NID (0, σ2f); eij= error experimental ~ NID (0, σ2); i = 1, 2,..., 19; j = 1, 2,..., 84.
Los análisis estadísticos se hicieron con el paquete Statistical Analysis System (SAS Institute, 1988). Para determinar las diferencias de altura, diámetro y volumen entre familias, y obtener los componentes de varianza σ2f y σ2e, se hicieron análisis de varianza con el procedimiento MIXED de SAS y el algoritmo de máxima verosimilitud restringida o REML (Littell et al., 1996).
Generalmente se supone que las familias de polinización libre representan familias de medios hermanos y que el componente de varianza de familias (σ2f) representa ¼ de la varianza genética aditiva (σ2A) (Falconer y Mackay, 1996). Esto sólo se cumple cuando los padres no están emparentados y la endogamia es cero; pero en la mayoría de los casos la correlación intra-clase de hermanos obtenidos por polinización libre es mayor de ¼ (Squillace, 1974). Un coeficiente de ¼ en esta situación ocasiona una sobrestimación de la σ2A, y por tanto de la heredabilidad y de las ganancias genéticas esperadas (Namkoong, 1966). Por lo anterior, para el cálculo de la varianza genética aditiva en este trabajo se empleó un coeficiente de determinación genética de 3, bajo el supuesto de que la correlación genética entre hermanos obtenidos por polinización libre es de ⅓ (Campbell, 1986; Sorensen y White, 1988).
Con los componentes de varianza se estimaron las heredabilidades en sentido estricto, tanto individual (h2i) como de medias de familia (h2f), para todas las variables de crecimiento, mediante las ecuaciones descritas por Falconer y Mackay (1996):
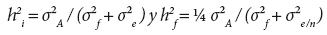
donde: σ2A = varianza aditiva (estimada como 3σ2f); σ2f= varianza de familias; σ2A= varianza del error; n = media armónica del número de plantas por familia en el ensayo. En Chichicaxtla (S1), la media armónica estimada en 2009 (4 años) para las variables ALT, D030 y NV fue de 9.44 árboles; en el 2010 (5 años) fue de 9.42 para ALT y 9.35 para DN y VOL. En Sierra Mojada (S2), la media armónica estimada para las variables antes descritas y en el mismo orden fue: 8.29, 7.85 y 6.69 árboles, respectivamente.
El error estándar de la heredabilidad [EE(h2)] se calculó con la fórmula de Dickerson (1969):
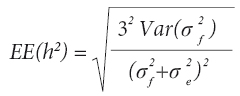
Se calcularon las correlaciones genéticas y fenotípicas entre todas las características, tanto a la misma edad como a diferentes edades (entre 4 y 5 años de edad). Las correlaciones fenotípicas se estimaron como coeficientes de correlación de Pearson, con el procedimiento CORR de SAS (SAS Institute, 1988). Para calcular las correlaciones genéticas entre pares de variables se utilizó la fórmula indicada por Falconer y Mackay (1996):
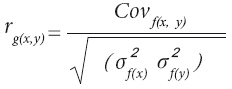
donde: Cov f (x , y) es la covarianza entre la variable x y la variable y, evaluadas en un misma familia; σ2f (x) y σ2f (y) son las varianzas de familias para la variables x e y. La covarianza entre ambas variables se calculó con la fórmula descrita por White Hodge (1989).
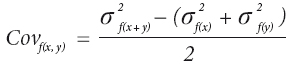
donde σ2f (x + y) es la varianza de familias para la variable (x + y), obtenida de la suma de las dos variables involucradas. Asimismo, se calculó el error estándar de las correlaciones genéticas con la ecuación de Falconer y Mackay (1996).
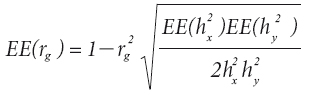
El análisis de correlaciones genéticas requiere que ambas variables tengan el mismo número de observaciones; por ello, y puesto que entre las evaluaciones del 2009 y 2010 se murió 1.5 % de los árboles, para este análisis se empleó una base de datos depurada (esto es, con información solamente para los árboles que tenían datos para ambos años). No obstante, y con fines de comparación, las heredabilidades se calcularon con la base depurada.
RESULTADOS Y DISCUSIÓN
En Chichicaxtla (2660 m de altitud, 12.6 °C media anual) hubo diferencias entre familias a los 4 años de edad en altura y número de verticilos con P = 0.0019, mientras que para el diámetro a 30 cm fue significativo con P = 0.0605 (Cuadro 1). A 5 años de edad las diferencias entre familias se incrementaron, en altura (P = 0.0002), diámetro a 30 cm (P = 0.0139), diámetro normal (P = 0.0011), número de verticilos (P = 0.0001) y volumen (P = 0.0004). También hubo diferencias significativas en la supervivencia promedio entre repeticiones (P = 0.0001) pero no entre familias (P = 0.2875). Con relación a los árboles muertos por frío no hubo diferencias entre repeticiones (P = 0.1254) ni entre familias (P = 0.1256). La supervivencia promedio de este ensayo a los 5 años de edad fue 83.3 % (entre familias, de 57.1 % a 100 %) y se registró 1.12 % de árboles muertos por frío (0 % a 14.3 % entre familias). La temperatura mínima media anual de este sitio es 5.1 °C, y la mínima histórica del mes de enero es 1.8 °C (Crookston, 2008), aunque no se tuvieron registros de esos años en el sitio.
En Sierra Mojada (2980 m) se tuvo una respuesta distinta, pues las menores temperaturas provocaron un crecimiento reducido de las plantas (la mínima promedio anual es 4.4 °C, y la mínima promedio del mes de enero es 0.9 °C, aproximadamente 1 °C menos que en Chichicaxtla, según datos de Crookston, 2008) (Cuadro 1), lo que ocasionó que las diferencias entre familias fueran reducidas y no significativas. A los 4 años de edad no hubo diferencias significativas entre familias para altura, diámetro a 30 cm y número de verticilos, cuyos valores de P fueron 0.2533, 0.3846 y 0.4848, respectivamente. Tal respuesta se mantuvo a los 5 años para altura, diámetro a 30 cm, diámetro normal, número de verticilos y volumen, con valores de P de 0.0510, 0.0998, 0.1225, 0.0627 y 0.1933, respectivamente. En cuanto a la supervivencia promedio hubo diferencias significativas entre repeticiones (P = 0.0001) y entre familias (P = 0.0115). No se detectaron diferencias significativas en la cantidad de árboles muertos por frío entre familias (P = 0.1074). La supervivencia promedio de este ensayo a los 5 años de edad fue 66.1 %, y osciló entre 37.5 % y 100 % entre familias; el porcentaje de árboles muertos por frío fue 6.2 %, y varió desde cero hasta 26.7 % entre familias.
La supervivencia promedio en Chichicaxtla fue 17 % mayor que en Sierra Mojada; ello puede atribuirse a las diferencias en temperaturas mínimas prevalecientes en cada sitio, de 5.5 °C vs. 4.4 °C en el promedio de las temperaturas mínimas, y de 12.6 °C vs. 11.1 °C de la temperatura media anual, ya que Chichicaxtla se encuentra a una altitud donde P. patula es una especie dominante (Vela, 1980). El ensayo de Sierra Mojada se estableció en un área donde domina P. montezumae Lamb., árbol que habita lugares más fríos que P. patula (Vela, 1980). Las bajas temperaturas al final del invierno pudieron haber debilitado los árboles, haciéndolos susceptibles a la falta de humedad de la primavera, provocando que algunos murieran por déficit de humedad (26.6 %) o se tornaran más susceptibles al ataque de un insecto descortezador (Dendroctonus mexicanus Hopkins) que eliminó a 1 % de los árboles.
Para todas las variables evaluadas, tanto a los 4 como a los 5 años de edad, los valores promedio más altos se registraron en Chichicaxtla (Cuadro 1), lo cual es explicable porque este sitio se ubica a una menor altitud (2660 m) que es más propia para esta especie, ya que Vela (1980) menciona que P. patula prospera mejor a altitudes que se encuentran entre 1900 m y 2400 m. A pesar del menor crecimiento en Sierra Mojada, el incremento proporcional en crecimiento entre estos dos años es mayor en este sitio de mayor elevación; por ejemplo, en altura entre los 4 y 5 años hay una diferencia de 51 % (1.45 m vs. 2.19 m), mientras que en Chichicaxtla fue de 35 % (2.8 m vs. 3.85 m). Así para D030 las diferencias entre años es de 63 % en Sierra Mojada vs. 32 % en Chichicaxtla. Para el número de verticilos la diferencia es similar, 33 % y 32 %.
Heredabilidades individual (h2i) y de familias (h2f) por sitio
En los dos años de evaluación, en Chichicaxtla las heredabilidades fueron más altas tanto a nivel individual como a nivel de familias (Cuadro 2), lo que se relaciona con las diferencias detectadas entre familias en el sitio de menor elevación. Lo anterior puede atribuirse a la menor mortandad y al mayor crecimiento promedio de las familias en localidad (que ocasionó una mayor diferenciación entre familias), factores que según White et al. (2007) redundan en un aumento de las heredabilidades. En Chichicaxtla las h2i a las edades 4 y 5 años fluctuaron entre 0.08 para diámetro a 30 cm y 0.18 para el número de verticilos, mientras que las de familias (h2f) oscilaron entre 0.15 y 0.28, respectivamente; valores que se clasificadan como heredabilidades bajas (Molina, 1992).
Una situación similar ocurrió en Sierra Mojada, lugar en donde las heredabilidades fueron menores que las reportadas por Farfán et al. (2002), quienes en Pinus ayacahuite Ehren. var. ayacahuite obtuvieron valores de h2i para altura y diámetro que oscilaron entre 0.31 y 0.52, y 0.32 y 0.55, respectivamente; y para h2f de 0.71 a 0.81 para la altura y de 0.71 a 0.83 para diámetro normal.
Estos valores de heredabilidad se asemejan más a los estimados por Valencia-Manzo y Vargas (1986) en un ensayo de progenies de P. patula de siete años, con valores de h2i de 0.14, 0.10 y 0.15 para altura, diámetro normal y volumen del fuste, y a los de Mora y Zamudio (2006) en pruebas de progenies de P radiata D. Don de siete años de edad, con valores de h2i de 0.08 a 0.13 para altura, de 0.0 a 0.13 para diámetro y de 0.0 a 0.09 para volumen. Las heredabilidades de cero aquí se atribuyeron a que la varianza de familias no tuvo incidencia significativa en la varianza total. No obstante, los valores de heredabilidades encontrados en el presente trabajo difieren de los estudios citados porque se refieren a especies y edades diferentes, y a que las condiciones de manejo o cultivo que se le dieron los ensayos quizás no fueron las mismas. Además, la heredabilidad disminuye en condiciones ambientales o de manejo variables y aumenta cuando éstas son más uniformes (Falconer y Mackay, 1996).
Es importante mencionar que en los dos sitios las heredabilidades aumentaron de un año a otro, lo que coincide con lo encontrado en otras especies de coníferas como Pinus ayacahuite var. ayacahuite (Farfán et al., 2002), P. caribaea Morelet (Otegbeye, 1988), P. caribaea var. hondurensis Barr. et Golf. (Dean et al., 1986) y Pseudotsuga mensiezii (Mirb.) Franco (Johnson et al., 1997). Este incremento en los valores de heredabilidad es atribuible al aumento de la competencia intergenotípica conforme aumenta el tamaño de los árboles, lo que probablemente acentuó las diferencias entre familias. También Zobel y Talbert (1988) mencionaron que la heredabilidad cambia con la edad y cuando el control genético de la característica cambia a medida que los árboles maduran.
Correlaciones genéticas y fenotípicas por sitio
En Chichicaxtla todas las correlaciones genéticas fueron altas y positivas entre las variables altura, diámetros y volumen, evaluadas a los 4 y 5 años de edad, así como entre edades (Cuadro 3), con magnitudes que variaron de 0.82 (i. e., diámetro a 30 cm a los 4 años y la altura total a los 5 años) hasta valores de 1.00 (i. e., las mismas variables medidas a los 4 años). Un aspecto relevante fue que las correlaciones genéticas para una misma característica (i. e., D030 a los 4 y a los 5 años) fueron altas y positivas, lo cual es conveniente porque indica que se pueden tomar decisiones a edades más tempranas, lo que reduce gastos en la evaluación de los genotipos.
Todas las correlaciones genéticas de los verticilos (Cuadro 3) fueron de magnitud baja a moderada (de 0.42 a 0.78), lo que indica, por ejemplo, que si se selecciona para mayor altura total o diámetro del fuste, aumentará también en algún grado el número de ramas. Sin embargo, tal aumento no será tan importante como lo sería el incremento en diámetro derivado de la correlación entre diámetro a 30 cm (D030 4 años) y diámetro normal (DN 4 años), que tienen una correlación genética muy elevada y positiva, lo que indica que si se selecciona con base en el diámetro normal también se obtendrá una respuesta correlacionada elevada en el diámetro a 30 cm. El uso del diámetro normal como criterio de selección sería más apropiado que el D030, pues el diámetro normal (a 1.30 m) es una variable más fácil y precisa de medir, especialmente cuando los árboles tienen muchas ramas en la parte inferior, como por lo regular ocurre en los primeros años de crecimiento.
Las correlaciones fenotípicas entre cada par de variables (Cuadro 3) resultaron altas y positivas, pero con coeficientes de correlación menores a las obtenidas en las correlaciones genéticas. Las características correlacionadas contra el número de verticilos presentaron los valores más bajos. Una de las más altas fue a los cuatro años entre diámetro normal y volumen, lo cual era de esperarse porque los árboles con diámetros mayores también presentan mayores volúmenes.
Las correlaciones genéticas y fenotípicas del ensayo de Sierra Mojada (Cuadro 4) difirieron de las de Chichicaxtla. En las correlaciones genéticas hubo valores muy bajos, incluso de cero; en las restantes, el signo fue positivo y sus valores oscilaron de 0.23 a 1.0. Los valores positivos y altos, al igual que el ensayo de Chichicaxtla, permiten elegir mejor la característica que más convenga para obtener una respuesta positiva al hacer la selección. Las correlaciones fenotípicas también fueron positivas y en algunos casos fueron más altas que las de Chichicaxtla. Hubo correlaciones importantes entre variables medidas tanto el mismo año como entre las registradas en diferentes años (Cuadro 4), lo cual es útil pues permite practicar la selección en etapas tempranas mediante el empleo de características fácilmente medibles.
En ambos ensayos hubo correlaciones genéticas y fenotípicas altas y positivas entre altura y diámetro normal, particularmente a los 5 años (Cuadros 3 y 4), lo que coincide con lo encontrado por Arregui et al. (1999), Farfán et al. (2002) y Sánchez et al. (2003). Por ejemplo, Valencia-Manzo y Vargas (2001) determinaron valores de rg ≥ 0.89 y rp ≥ 0.78 para diámetro, altura y volumen en plantaciones de P. patula con 6 años de edad. Sin embargo, las correlaciones genéticas que fueron altas y positivas en un ensayo, no necesariamente lo fueron en el otro. Por ejemplo, en árboles de 5 años en Chichicaxtla hubo una correlación genética alta y positiva entre diámetro a 30 cm y diámetro normal, mientras que en Sierra Mojada para las mismas características y a la misma edad la correlación genética fue positiva pero inferior a la de Chichicaxtla. Una situación similar se presentó con las características correlacionadas entre el número de verticilos y la altura, que difirieron de un sitio a otro. Estas diferencias han sido atribuidas a causas genéticas y ambientales que afectan los caracteres de los árboles a través de mecanismos fisiológicos diferentes (Falconer y Mackay, 1996). Algo importante es que las correlaciones genéticas dependen de las frecuencias génicas, de forma que podrían variar en diferentes poblaciones (Bohren et al., 1966).
CONCLUSIONES
Se encontraron diferencias significativas en el crecimiento de las 84 familias evaluadas en cada ensayo. La mejor respuesta se detectó en Chichicaxtla (el sitio ubicado a menor altitud). Una situación similar se observó para las heredabilidades, correlaciones genéticas y fenotípicas, probablemente debido a que las condiciones ambientales de esa localidad fueron más favorables para el desarrollo de la especie.
Las heredabilidades, tanto individuales como de familias, aumentaron con la edad en la mayoría de las características evaluadas, lo cual se atribuye a la diferenciación entre familias que ocurre de un año a otro.
En ambos sitios, la mayoría de las correlaciones genéticas y fenotípicas fueron positivas y altas, particularmente entre las variables altura, diámetros a 0.30 y a 1.30 m, y volumen, lo que resulta útil para fines de selección temprana.
AGRADECIMIENTOS
A los Ings. León Jorge Castaños Martínez y Salvador Castro Zavala, por el establecimiento de la prueba de progenies. Al Fondo CONAFOR-CONACYT 2003-C03-10714, por el apoyo al proyecto "Establecimiento de huertos semilleros con material genéticamente superior de Pinus patula y P. greggii", y a la Línea Prioritaria de Investigación Núm. 1 del Colegio de Postgraduados.
BIBLIOGRAFÍA
Arregui A, S Espinel, A Aragonés, R Sierra (1999) Estimación de parámetros genéticos en un ensayo de progenie de Pinus radiata D. Don en el país vasco. Inv. Agr. Sist. Rec. For. 8:119-128. [ Links ]
Bohren B B, W G Hill, A Robertson (1966) Some observations on asymmetrical correlated responses to selection. Genet. Res. 7:44-57. [ Links ]
Campbell R K (1986) Mapped genetic variation of Douglas-fir to guide seed transfer in southwest Oregon. Silvae Genet. 35:85-96. [ Links ]
Carrillo A F, M Acosta M, G Tenorio G, F Becerra L (2004) Tabla de volumen para Pinus patula Schl. et Cham., en el Estado de Hidalgo. Folleto Técnico No. 2. INIFAP-SAGARPA. 16 p. [ Links ]
Clausen K (1990) Diseños genéticos y pruebas de progenie. In: Memoria Mejoramiento Genético y Plantaciones Forestales. Centro de Genética Forestal, A. C. Chapingo, México. pp:67-77. [ Links ]
Crookston N L (2008) Custom data requests. In: Current and Future Climate Estimates, Plant-Climate Relationships, and Potential Impacts of Global Warming Scenarios in Western North America and Mexico. http://forest.moscowfsl.wsu.edu/climate/customData/. USDA Forest Service, Moscow Forestry Sciences Laboratory. [ Links ]
Dean C A, P P Cotterill, R Eiseman (1986) Genetic parameters and gains expected from selection in Pinus caribaea var. hondurensis in Northern Queensland, Australia. Silvae Genet. 35:229-236. [ Links ]
Dickerson G E (1969) Techniques for research in quantitative animal genetics. In: Techniques and Procedures in Animal Science Research. Am. Soc. Animal Res. Albany, N. Y. pp:36-79. [ Links ]
Dvorak W S, G R Hodge, J E Kietzka, F Malán, L F Osorio, T K Stanger (2000) Pinus patula. In: Conservation & Testing of Tropical & Subtropical Forest Tree Species by the CAMCORE Cooperative. College of Natural Resources, NCSU. Raleigh, NC. pp:149-173. [ Links ]
Falconer D S, T F C Mackay (1996) Introduction to Quantitative Genetics. 4th ed. Longman. Exxex, UK. 464 p. [ Links ]
Farfán V E G, J Jasso M, J López U, J J Vargas H, C Ramírez H (2002) Parámetros genéticos y eficiencia a la selección temprana en Pinus ayacahuite Ehren. var. ayacahuite. Rev. Fitotec. Mex. 25:239-246. [ Links ]
Gillespie A J R (1992) Pinus patula Schiede and Deppe. Patula pine. Southern Forest Experiment Station, Forest Service, USDA. SO-ITF-SM-54. New Orleans, LA. 5 p. [ Links ]
Greenwood M S (1977) The role of dormancy in the development of male and female strobili of loblolly pine. For. Sci. 23:373-375. [ Links ]
Johnson G R, R A Sniezko, N L Mandel (1997) Age trends in Douglas-fir genetic parameters and implication for optimum selection age. Silvae Genet. 46:349-358. [ Links ]
Littell R C, G A Milliken, W W Stroup, R D Wolfinger (1996) SAS System for Mixed Models. SAS. Cary, NC. 633 p. [ Links ]
Molina G J D (1992) Introducción a la Genética de Poblaciones y Cuantitativa (Algunas Implicaciones en Genotecnia). A. G. T. México, D. F. 349 p. [ Links ]
Monroy R C (1995) Pinuspatula Schl. et Cham., en México. Folleto Técnico No. 29. SAGAR/CIRGOC-INIFAP. Veracruz, Ver. 145 p. [ Links ]
Mora F, F Zamudio (2006) Variabilidad genética del crecimiento en progenies selectas de Pinus radiata. Ciên. Flor. 16:399-405. [ Links ]
Namkoong G (1966) Inbreeding effects on estimation of genetic additive variance. For. Sci. 12:8-13. [ Links ]
Nyoka B I, J S Birks, C M Gumbie (1994) Pinus patula progeny test: heritability estimates and genetic correlations between fifth- and eighth-year traits. South Afr. For. J. 168:23-26. [ Links ]
Otegbeye G O (1988) Genetic variation in growth and form characteristics of Pinus caribaea. Silvae Genet. 37:232-236. [ Links ]
Sáenz-Romero C, H Nienstaedt, J J Vargas-Hernández (1994) Performance of Pinus patula genotypes selected in South Africa and growing in their native Mexican environment. Silvae Genet. 43:73-81. [ Links ]
Sánchez M V, J G Salazar G, J J Vargas H, J López U, J Jasso M (2003) Parámetros genéticos y respuesta a la selección en características del crecimiento de Cedrela odorata L. Rev. Fitotec. Mex. 26:19-27. [ Links ]
SAS Institute (1988) SAS/STAT Guide for Personal Computers. SAS. Cary, NC. 378 p. [ Links ]
Sorensen F C, T L White (1988) Effect of natural inbreeding on variance structure in tests of wind-pollination Douglas-fir progenies. For. Sci. 34:102-118. [ Links ]
Squillace A E (1974) Average genetic correlations among offspring from open-pollinated forest trees. Silvae Genet. 23:149-156. [ Links ]
Valencia-Manzo S, J J Vargas-Hernández (1986) Control genético de la velocidad de crecimiento y características de la madera en Pinus patula. Agrociencia 30:265-273. [ Links ]
Valencia-Manzo S, J J Vargas-Hernández (2001) Correlaciones genéticas y selección simultánea del crecimiento y densidad de la madera en Pinus patula. Agrociencia 35:109-120. [ Links ]
Vargas-Hernández J J, W T Adams (1992) Age-age correlation and early selection for wood density in young coastal Douglas-fir. For. Sci. 38:467-478. [ Links ]
Vela G L (1980) Contribución a la ecología de Pinus patula Schl. et Cham. Publicación Especial No. 19. INIF/SARH. México, D. F. 109 p. [ Links ]
Velázquez M A, G Ángeles P, T Llanderal O, A Román J, J V Reyes H (2004) Monografía de Pinus patula. CONAFOR/Colegio de Postgraduados. México, D. F. 124 p. [ Links ]
White T L, G R Hodge (1989) Predicting Breeding Values with Applications in Forest Tree Improvement. Kluwer Acad. Pub. The Netherlands. 368 p. [ Links ]
White T L, T W Adams, D B Neale (2007) Forest Genetics. CAB Int., Oxford. 682 p. [ Links ]
Wormald T J (1975) Pinus patula. Tropical Forestry Paper No. 7. Tropical Forestry Institute. Commonwealth Forestry Institute. Oxford, England. 172 p. [ Links ]
Wright J W (1964) Mejoramiento Genético de los Árboles Forestales. FAO: Estudio de Silvicultura y Productos Forestales No. 16. Roma, Italia. 436 p. [ Links ]
Wu H X (1998) Study of early selection in tree breeding. Silvae Genet. 47:146-155. [ Links ]
Zobel B J, J T Talbert (1988) Técnicas de Mejoramiento Genético de Árboles Forestales. Ed. Limusa. México, D. F. 545 p. [ Links ]

