










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista fitotecnia mexicana
versão impressa ISSN 0187-7380
Rev. fitotec. mex vol.35 no.3 Chapingo Jul./Set. 2012
Artículos científicos
Efecto del AG4/7 y del anillado en el tronco sobre el crecimiento del brote en Pseudotsuga menziesii (Mirb.) Franco
Effect of GA4/7 and stem girdling on shoot growth in Pseudotsuga menziesii (Mirb.) Franco
Liliana Muñoz Gutiérrez1,2, J. Jésus Vargas Hernández1*, Héctor M. de los Santos Posadas1 y Javier López Upton1
1 Programa Forestal. Colegio de Postgraduados-Campus Montecillo. Km. 36.5 Carretera Federal México-Texcoco. 56230, Montecillo, Texcoco, Edo. de México. * Autor para correspondencia (vargashj@colpos.mx)
2 Campo Experimental San Luis, Centro de Investigación Regional del Noreste, Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Km. 14.5 Carretera San Luis-Matehuala. 78430, Soledad de Graciano Sánchez, San Luis Potosí, SLP, México.
Recibido: 8 de Junio del 2011
Aceptado: 6 de Junio del 2012
Resumen
Para evaluar posibles efectos colaterales negativos de los tratamientos de inducción floral en individuos juveniles de Pseudotsuga menziesii sobre el desarrollo de la copa de los árboles, aquí se analizó el efecto del AG4/7 y del anillado en el tronco, así como el impacto de la formación de estróbilos femeninos, en el crecimiento de los brotes. Los tratamientos se aplicaron en dos ensayos con árboles de 10 a 13 años de edad establecidos en terreno a cielo abierto (Ensayos 1 y 2) y uno en árboles de cuatro años crecidos en maceta y vivero (Ensayo 3). Se utilizaron cuatro dosis de AG4/7 (0, 1.27, 2.55 y 5.10 mg cm-2 de área basal) inyectadas en el tronco del árbol, en combinación con el anillado del tronco (con y sin anillado). Durante el periodo de crecimiento se midió la longitud total de los brotes y se ajustó la curva de crecimiento promedio con el modelo de Schumacher. Se encontraron diferencias (P < 0.05) entre ensayos, así como entre los niveles de anillado y las dosis de AG4/7, en la longitud del brote, pero no se encontró un efecto significativo de las interacciones entre factores. En el Ensayo 1 el brote creció 30 % más que en los otros ensayos, pero en el Ensayo 2 inició primero el crecimiento y alcanzó una mayor tasa máxima de crecimiento; los árboles de menor edad del Ensayo 3 presentaron la menor longitud del brote. El anillado redujo en 9 % el crecimiento del brote y lo retrasó en 6 d con respecto a los árboles no anillados; en cambio, el AG4/7 aumentó en 12 % la longitud total del brote y aceleró su crecimiento. La formación de estróbilos femeninos ocasionó un retraso en el crecimiento del brote, pero no redujo su longitud total. El uso de AG4/7 en dosis de 1.27 mg cm-2 para inducir floración en árboles juveniles de P. menziesii no tiene efectos negativos sobre el crecimiento de los brotes; pero el uso repetido del anillado podría afectar negativamente el desarrollo y vigor de los árboles.
Palabras clave: Pseudotsuga menziesii, ácido giberélico (AG4/7), anillado, inducción floral, longitud de brote, tasa de crecimiento.
Abstract
To evaluate potential negative collateral effects of flower induction treatments applied to juvenile Pseudotsuga menziesii trees on crown development (GA4/7 application, stem girdling on shoot elongation, and the impact of female cones (strobili) induction on shoot elongation), were evaluated. Induction treatments were applied in two field trials with 10 to 13-years-old trees (Trials 1 and 2) and one pot trial in the nursery with 4-years-old trees (Trial 3). Four doses of GA4/7 (0, 1.27, 2.55 and 5.10 mg cm-2 basal area), applied by injection into the tree trunk, combined with stem girdling (with or without girdling) were evaluated. During the growing period the total length of shoots was measured and the average growth curve was adjusted using the Schumacher model. Differences in shoot length were found across trials as well as among GA4/7 doses and girdling levels, but no significant interactions between factors were detected. In Trial 1, shoots grew 30 % more than in the others, but in Trial 2 shoot growth started first and reached highest maximum growth rate; the youngest trees in Trial 3 showed the shortest shoot length. Stem girdling reduced shoot growth by 9 % and slowed it by 6 d, whereas GA4/7 increased 12 % the final shoot length and accelerated its growth. The induction of female cones caused a slight delay in the shoot growth curve, but final shoot length was not reduced. Use of GA4/7 in doses of 1.27 mg cm-2 to induce flowering in juvenile trees of P. menziesii did not have negative effects on shoot growth; however, repeated use of stem girdling could negatively affect tree growth and vigor.
Key words: Pseudotsuga menziesii, gibberellic acid (GA4/7), stem girdling, flower induction, shoot length, growth rate.
INTRODUCCIÓN
Las especies forestales leñosas, especialmente las coníferas de clima templado frío, requieren de un periodo prolongado para alcanzar la madurez sexual (Philipson, 1990; Aderkas et al., 2004), lo cual limita en gran medida la producción de semilla en los programas de mejoramiento genético. Una alternativa para acelerar la producción de semilla en estas especies es la inducción de estructuras reproductivas en árboles juveniles. Existen diferentes técnicas y tratamientos de cultivo utilizados con éxito para inducir floración precoz en especies de coníferas, como la aplicación de reguladores del crecimiento, combinada con tratamientos culturales de anillado en el tronco, poda, tratamientos de calor, sequía y fertilización, entre otros (Pharis et al., 1980; Ross, 1983; Webber et al., 1985; Bonnet, 1987a; Philipson, 1990).
Los tratamientos de inducción floral utilizados en las especies de coníferas estimulan el desarrollo de yemas laterales, seguido de la diferenciación y crecimiento de las estructuras reproductivas (Greenwood, 1981); sin embargo, algunos de estos tratamientos también producen diversos tipos de estrés o inducen competición por recursos entre órganos (Ross y Bower, 1991), efectos que pueden retardar o reducir el crecimiento vegetativo de las ramillas (Pharis y Kuo, 1977) y ocasionar efectos negativos sobre el desarrollo y vigor del árbol en años posteriores. Tal efecto puede ser mayor en árboles juveniles en los que el crecimiento vegetativo requiere mayor cantidad de recursos, mientras que en árboles maduros los órganos vegetativos tienen menor prioridad, por su menor tasa de crecimiento, lo que permite una mayor disponibilidad de giberelinas para los procesos reproductivos (Ross, 1983).
En un estudio con Pseudotsuga menziesii (Mirb.) Franco, la aplicación de AG4/7 (una mezcla de AG4 y AG7) estimuló la floración pero redujo la tasa relativa de crecimiento de los brotes (Cherry et al., 2007) durante el periodo de desarrollo de los conos; los tratamientos que estimularon una mayor producción de estructuras reproductivas (AG4/7 solo y AG4/7 + anillado) ocasionaron menor longitud total o menor tasa de crecimiento del brote. Además, la época de aplicación del AG4/7 puede influir en la respuesta del crecimiento del brote (Owens et al., 1985); así, las aplicaciones hechas al inicio del periodo de crecimiento de los brotes promueven una mayor longitud de éstos, mientras que las aplicaciones tardías tienen muy poco efecto o ninguno. Por ejemplo, en Tsuga heterophylla (Raf.) Sarg., aplicaciones foliares de AG4/7 a 200 mg L-1 durante el crecimiento del brote ocasionaron un aumento promedio de 80 % en su longitud final, pero la respuesta difirió entre clones lo que evidencia que también influye el genotipo (Harrison y Owens, 1994).
Ross (1983) demostró en P. menziesii que la aplicación de AG4/7 antes del inicio del crecimiento de las yemas vegetativas aumenta la longitud del brote pero no la floración, mientras que al aplicarlo durante el periodo de rápido crecimiento se incrementa la floración pero no la longitud del brote. Consistente con lo anterior, Webber et al. (1985) encontraron que aplicaciones de AG4/7 en P. menziesii estimulan preferentemente el crecimiento vegetativo y que la floración ocurre sólo cuando se aplica una dosis suficiente para satisfacer la demanda de ambos procesos.
En un trabajo previo (Muñoz-Gutiérrez et al., 2010) se describió el efecto del anillado y de la aplicación de diferentes dosis de AG4/7 sobre la inducción floral en árboles jóvenes de Pseudotsuga menziesii originados por semilla. En este trabajo se analiza el efecto de esos mismos tratamientos sobre el crecimiento de los brotes, así como la posible interacción de este proceso con la inducción de estructuras reproductivas, con el propósito de evaluar si estas técnicas de inducción reproductiva en árboles jóvenes tienen algún impacto colateral negativo sobre el desarrollo de la copa del árbol.
MATERIALES Y MÉTODOS
Localización de los ensayos de campo
Se llevaron acabo tres ensayos independientes en el Conjunto Predial Forestal, en el municipio de Aquixtla, Puebla (19° 43' 13'' LN y 97° 59' 20'' LO), dos de ellos con árboles jóvenes de P. menziesii establecidos en el bosque y el tercero con plantas en maceta. El Ensayo 1 se efectuó en el paraje El Manantial a 2900 m de altitud, en una plantación de árboles de navidad de 10 años de edad (plantados en este paraje en el 2000, a la edad de 2 años; la semilla para producir estos árboles se recolectó en Mineral del Chico, Estado de Hidalgo, 20° 10' 18'' LN, 98° 44' 00'' LO, 3130 m de altitud), y las plantas crecieron con luz solar plena y podas recurrentes de la copa. El Ensayo 2 se estableció en el Área semillera a 2800 msnm, en una plantación con árboles de 11 a 13 años de edad (plantados en 1996 y 1997, a la edad de un año, originados con semilla recolectada en Terrenates, Estado de Tlaxcala, sitio ubicado a 19° 31' 11'' LN, 97° 53' 52'' LO, 2750 msnm; y en Huayacocotla, Estado de Veracruz, a 20° 24' 16'' LN, 98° 28' 06'' LO, 2510 msnm); estas plantas crecieron bajo el dosel de árboles adultos de Pinus patula, sin poda de ramas durante los cuatro años previos al estudio (2004-2007). El Ensayo 3 se estableció en el vivero del paraje El Manantial, a 2900 msnm, en condiciones de luz solar plena, con plantas de cuatro años de edad en maceta (originados con semilla recolectada en Mineral del Chico, Estado de Hidalgo).
En cada paraje se instaló una micro estación meteorológica automatizada para registrar temperatura y precipitación. En cada ensayo se seleccionaron los 32 árboles más homogéneos en cuanto a altura (Ensayo 1: 1.77 ± 0.10 m; Ensayo 2: 2.80 ± 0.18 m; Ensayo 3: 0.80 ± 0.13 m), diámetro (Ensayo 1: 5.4 ± 0.3 cm; Ensayo 2: 6.1 ± 0.5 cm; Ensayo 3: 2.8 ± 0.3 cm), y de condiciones de la copa. En cada árbol se midió el diámetro del tronco a 50 cm (Ensayos 1 y 2) o 10 cm (Ensayo 3) del suelo, para calcular el área transversal del mismo.
Definición y aplicación de los tratamientos
En cada ensayo se evaluó el anillado (con y sin) en combinación con cuatro dosis de AG4/7 (0, 1.27, 2.55 y 5.10 mg cm-2 del área transversal del tronco, equivalentes a 0, 100, 200 y 400 mg de AG4/7 en un árbol de 10 cm de diámetro del tronco), en un arreglo factorial y diseño experimental completamente al azar con cuatro repeticiones. Estas concentraciones de AG son similares a las que se han utilizado en otras especies de coníferas para estimular floración, pero ajustadas por las diferencias en tamaño (área transversal del tronco) de los árboles. El anillado consistió en hacer dos cortes transversales al tronco con un serrucho, en forma de media luna, de manera que cada corte abarcara aproximadamente 60 % de la circunferencia del tronco; el primer corte se hizo a 50 cm (Ensayos 1 y 2) o a 10 cm (Ensayo 3) del suelo, y el segundo corte a una distancia del primero igual a 1.5 veces el diámetro de tronco (Muñoz-Gutiérrez et al., 2010). Los tratamientos se aplicaron al inicio del crecimiento del brote terminal y en cada árbol se determinó el estado fenológico de la copa (porcentaje de brotación de las yemas vegetativas en la copa del árbol).
El AG4/7 se aplicó en una solución de alcohol etílico a 95 % (v/v), con una concentración base de 30 mg mL-1, en un solo evento en los tres ensayos. En el tronco se hicieron orificios inclinados a 10 cm por arriba del anillado superior y a una altura similar en los árboles no anillados. La cantidad requerida (mL) de solución, en función de la dosis y área transversal del tronco, se aplicó con micro pipeta; en el testigo se aplicó un volumen equivalente de alcohol etílico a 95 %; los orificios se taparon con cera para evitar pérdida de la solución (Muñoz-Gutiérrez et al., 2010).
Variables medidas y análisis de los datos
En los 32 árboles incluidos en cada ensayo se seleccionaron y marcaron siete (Ensayos 1 y 2) o cinco (Ensayo 3) ramillas terminales por árbol distribuidas a lo largo de la copa y contadas a partir del ápice; cada ramilla se identificó con un listón de color específico en función de su posición en la copa. Al momento de aplicar los tratamientos se determinó el estado fenológico de la yema terminal de cada ramilla y la longitud inicial del brote (en caso de que la yema ya hubiera iniciado el alargamiento del brote). Posteriormente, cada dos semanas se midió la longitud total del brote en cada ramilla, a partir de la base de la yema terminal, con una regla graduada en mm; la toma de datos continuó durante un periodo de siete meses (del 21 de marzo hasta el 4 de octubre), para cubrir todo el periodo de crecimiento de los brotes. Cuando alguna ramilla marcada murió o se dañó, se eliminó de la base de datos. Con los datos de crecimiento obtenidos se generó una curva de crecimiento para cada brote marcado.
Con los promedios de longitud de brote por árbol se hizo un análisis de varianza para determinar el efecto de los factores ensayo, anillado y dosis de AG4/7, con el procedimiento GLM (SAS Institute, 1998); en los factores que mostraron efectos significativos se hizo una comparación de medias mediante la prueba de Tukey y la prueba de contrastes (P ≤ 0.05). Para evaluar el efecto de los tratamientos sobre el patrón de crecimiento del brote, con el procedimiento MODEL (SAS Institute, 1998) se estimaron los parámetros del siguiente modelo de regresión no lineal: Longitud = f (dosis, anillado). Posteriormente, con los parámetros estimados se ajustó el modelo de tipo Schumacher, por medio del método de mínimos cuadrados, con el paquete estadístico S-Plus® (S-Plus, 2005). El modelo de Schumacher utilizado fue el siguiente:

donde Y = longitud del brote (cm); d = número de días; e = base del logaritmo natural; β0 y β1 = parámetros de regresión del modelo.
Este modelo permite separar los parámetros específicos β0 y β1 que representan la asíntota o valor máximo que puede alcanzar la curva de crecimiento, y la fecha en que ocurre la tasa máxima de crecimiento acumulado, respectivamente. Para comparar el efecto de los factores ensayo, dosis de AG4/7 y anillado del tronco sobre la curva de crecimiento y longitud total del brote, se tomó como base la Ec. 1, de la que se derivó la siguiente ecuación:

donde: Di = variable indicadora del i-ésimo nivel de dosis; Aj = variable indicadora del j-ésimo nivel de anillado; y Ek = variable indicadora del k-ésimo ensayo; las demás variables fueron definidas anteriormente.
Para evaluar la relación entre la inducción de estructuras reproductivas y el crecimiento del brote, se hizo un análisis de varianza de la longitud total del brote en los árboles del Ensayo 2, en el que se obtuvo la mayor proporción de árboles con estructuras reproductivas; los árboles de este ensayo que recibieron dosis de 1.27 y 5.10 mg cm-2 de AG4/7 se separaron en dos grupos (árboles con y sin estróbilos), y se comparó la longitud promedio del brote entre ellos. Además, se comparó la curva de crecimiento del brote de los dos grupos de árboles mediante el procedimiento de ajuste del modelo de Schumacher (Ec. 1) descrito anteriormente:

donde F = variable indicadora de la formación de estróbilos femeninos; las demás variables fueron definidas anteriormente.
RESULTADOS
Crecimiento promedio del brote en los tres ensayos
Los análisis de varianza mostraron diferencias significativas entre ensayos (P < 0.001), entre niveles de anillado y entre dosis de AG4/7 (P < 0.05), en la longitud del brote al final del periodo de crecimiento, pero no se encontró efecto significativo de las interacciones entre los factores evaluados (Cuadro 1).
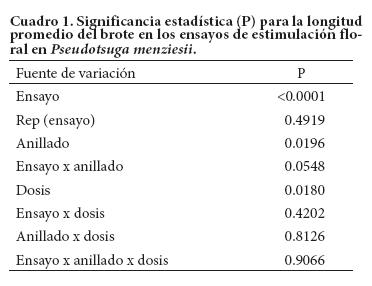
El crecimiento total del brote en el Ensayo 1 fue 30 % mayor que en el Ensayo 2 (28.2 vs. 21.6 cm), en árboles de edad similar; en los árboles más jóvenes del Ensayo 3 el crecimiento del brote fue menor (Figura 1a). Por otro lado, el anillado en el tronco redujo en 9 % la longitud promedio del brote (21.3 vs. 23.3 cm; Figura 1b) mientras que la aplicación de AG4/7 aumentó 12 % en promedio la longitud del brote con respecto a los árboles que no recibieron este regulador del crecimiento (Figura 1c); la prueba de contrastes indicó que el testigo (sin AG4/7) fue menor (P ≤ 0.01) al promedio de los árboles que recibieron AG4/7.
Parámetros estimados del modelo de crecimiento del brote
La mayoría de los factores evaluados tuvieron un efecto significativo en el patrón de crecimiento del brote (Cuadro 2). El modelo básico (β00 y β10) representa la curva de crecimiento del brote en los árboles testigo del Ensayo 3 (sin anillado y sin AG4/7 ); los otros parámetros muestran el efecto aditivo de la dosis de AG4/7, del anillado y del ensayo, que influyeron tanto en el crecimiento total del brote (β0) como en el momento en que ocurrió la tasa máxima de crecimiento acumulado (β1); los valores positivos o negativos de los parámetros indican si el efecto específico de cada factor fue aumentar o reducir los valores respectivos (β00 y β10) en el modelo básico. Por ejemplo, en la Figura 2 se observa que los árboles del Ensayo 1 alcanzaron una mayor longitud del brote (β0E1 =13.13) que en los otros ensayos, pero los árboles del Ensayo 2 iniciaron primero el alargamiento del brote y alcanzaron la tasa máxima de crecimiento acumulado casi un mes antes (β1E2 = -37.05) que en los otros dos ensayos. En el Ensayo 3, con árboles en un estado fenológico menos avanzado que en los otros dos ensayos, el alargamiento del brote se retrasó y tuvo menor tasa de crecimiento y menor crecimiento potencial (Figura 2b).
Efecto del anillado y de la dosis de AG4/7 sobre el crecimiento del brote
El anillado redujo y retrasó el crecimiento del brote en los tres ensayos (Figura 3); en promedio, el crecimiento potencial estimado en los árboles no anillados fue 18 % mayor (38.6 vs. 35.4 cm en el Ensayo 1), y la tasa máxima de crecimiento se alcanzó casi 6 d antes que en los árboles anillados (β1A1 = 5.86). De la misma manera, la tasa máxima de crecimiento acumulado fue 22 % mayor en los arboles no anillados.
Consistentemente, la dosis de 2.55 mg cm-2 de AG4/7 ocasionó el mayor alargamiento del brote en los tres ensayos, 12 % mayor que en los árboles testigo (Figura 4); sin embargo, también ocasionó un retraso de 11 d para alcanzar la tasa máxima de crecimiento acumulado (β1D2 = 11.80). En contraste, la dosis de 1.27 mg cm-2 tuvo un efecto menor sobre el crecimiento total del brote, pero aceleró en casi 6 d la fecha (β1D1 = -5.66) de la tasa máxima de crecimiento (Cuadro 2) con respecto al testigo. Las mayores tasas de crecimiento en los tres ensayos se alcanzaron con la dosis baja (1.27 mg cm-2) de AG4/7.
Relación entre inducción floral y crecimiento del brote
Los análisis de varianza no indicaron diferencias significativas en el crecimiento total del brote en los árboles del Ensayo 2 por efecto de la formación de estróbilos femeninos o por la dosis de AG4/7 (1.27 vs. 5.10 mg cm-2). Sin embargo, el análisis de regresión de las curvas de crecimiento del brote mostró que la floración influyó sobre la fecha de la tasa máxima (β1F) de crecimiento, y que la dosis de AG4/7 afectó su valor máximo (β0D3) o crecimiento potencial (Cuadro 3). Aunque la formación de estróbilos femeninos no redujo el crecimiento potencial del brote, causó un retraso de 10 d (β1F = 9.62) en la fecha en que ocurrió la tasa máxima de crecimiento, así como una reducción en ésta (Figura 5).
DISCUSIÓN
Diferencias entre ensayos
Las diferencias observadas entre ensayos en la longitud total del brote se pueden deber a diversos factores, que incluyen la edad y las condiciones de crecimiento y manejo de los árboles. Las plantas del Ensayo 3 eran más jóvenes que las de los otros dos ensayos y en las especies de coníferas el crecimiento anual del brote aumenta gradualmente con la edad en los primeros años de vida (Borchert, 1976). Los árboles del Ensayo 1 fueron podados el año previo y se encontraban creciendo a plena luz solar, a diferencia de los árboles del Ensayo 2 que crecieron bajo sombra y no fueron podados en los últimos años. También pueden deberse al estado fenológico del brote al momento de la aplicación de los tratamientos, ya que los árboles del Ensayo 2 se encontraban más avanzados fenológicamente que los del Ensayo 1 (Muñoz-Gutiérrez et al., 2010); por tanto, el efecto de los tratamientos en el Ensayo 2 pudo haber sido menor.
Efecto de la aplicación de AG4/7
Los tratamientos con AG4/7 para inducir floración influyeron en el crecimiento de los brotes, consistente con el estudio de Pharis y Kuo (1977), quienes reportaron que en especies de coníferas la inducción de estructuras reproductivas por medio de AG4/7 a menudo está acompañada por un incremento en el crecimiento del brote, mientras que otras técnicas de inducción pueden retrasar y reducir el crecimiento (Bonnet et al., 1982).
En el presente trabajo se encontró que en los árboles que recibieron AG4/7 los brotes crecieron en promedio 12 % más que en los árboles testigo. Sin embargo, en otros estudios se ha encontrado que el AG4/7 no tiene efecto sobre la longitud final del brote, como en Tsuga heterophylla (Harrison y Owens, 1994), Picea mariana (P. Mill.) B. S. P. (Smith y Greenwood, 1997) y P. menziesii (Ross y Pharis, 1976), pero sí en la tasa máxima de crecimiento. Ho y Eng (1995), por ejemplo, determinaron que el AG4/7 acelera el crecimiento de los brotes en Pinus strobus L. aunque no haya tenido efecto sobre la longitud final. En el presente estudio, las mayores tasas de crecimiento acumulado se alcanzaron cuando se utilizó una dosis baja de AG4/7.
Según Ross (1983), el AG4/7 no afecta el patrón de crecimiento del brote, sino sólo la tasa de crecimiento y la longitud final. En cambio, la tasa de crecimiento de los brotes depende de la fenología del árbol; por su parte, Owens et al. (1985) señalaron que al momento de reactivación de las yemas en P. menziesii la división celular es frecuente, pero no el alargamiento de las células; después de la brotación la división celular disminuye y el alargamiento de las células incrementa rápidamente y continúa hasta que el brote alcanza 90 % de su longitud final. La acción de las giberelinas depende de varios factores, como la posición del brote en la copa, la disponibilidad de luz, y el momento y método de aplicación (Chalupka, 1980), entre otros.
Owens et al. (1985) demostraron la importancia de la época de aplicación del AG4/7 debido a su efecto diferencial sobre la división y el alargamiento de las células. Ross (1983) obtuvo el máximo crecimiento del brote cuando el AG4/7 se aplicó antes de la reactivación de las yemas, pero sólo hubo un leve incremento cuando se aplicó 4.5 semanas después de la reactivación; esto sugiere que el AG4/7 afecta principalmente el alargamiento de las células. Además, el nivel y distribución de carbohidratos varía durante el crecimiento del brote, por lo que la competición entre órganos por estos recursos también influye sobre la longitud total del brote en árboles con diferentes condiciones fisiológicas y ambientales (Owens et al., 1985; Ross y Bower, 1991).
Efecto del anillado en el tronco
En otras especies se han encontrado resultados similares en el efecto colateral del anillado sobre el crecimiento del brote; en árboles injertados de Picea sitchensis el anillado redujo en 15 % la longitud anual del brote con respecto a los árboles no anillados (Longman et al., 1986); sin embargo, en P. menziesii se observó un aumento en la longitud promedio del brote (0.98 vs. 1.27 m año-1) por efecto del anillado, y en Pinus contorta Dougl. y P. rigida (Mill.) la longitud de brote no se afectó (Wilson y Gartner, 2002). Las diferencias entre estudios pueden deberse a diferencias en el momento de realizar el anillado, ya que en las especies de Pinus se hizo antes de que iniciara el crecimiento del brote, mientras que en P. menziesii fue durante la reactivación de las yemas.
Cuando se anilla el tronco se afecta parcialmente el floema, lo que ocasiona un bloqueo en el transporte de carbohidratos y de las hormonas de crecimiento; además, el anillado reduce la conductividad hidráulica del xilema, y ocasiona un estrés que afecta el crecimiento del árbol (Zwieniecki et al., 2004). El anillado en el tronco también puede estimular el flujo de carbohidratos (Ebell, 1971), o reducir la disponibilidad de carbohidratos a la raíz y provocar un estrés. Los tratamientos culturales que provocan estrés pueden alterar el metabolismo de las giberelinas, ya sea para incrementar o reducir su disponibilidad (Philipson, 1985).
En el efecto del anillado influyen muchos factores, como especie, edad del árbol, tipo de herida, severidad de la herida, regulación de la herida y la interacción con otros tratamientos o condiciones como la fertilización, estrés por humedad, factores climáticos (Puritch, 1972; Bonnet 1987a; Ross y Bower, 1991), entre otros. Al respecto, Ross et al. (1984) señalaron que los factores que inducen estrés, combinados con aplicaciones de AG, tienen un efecto sinérgico sobre la floración. Esto apoya la hipótesis de que las giberelinas causan una redistribución de nutrientes dentro de los brotes, lo que puede retrasar su desarrollo. No obstante, en este trabajo no se encontró una interacción significativa de los factores evaluados (Cuadro 1).
Efecto de la inducción floral sobre el crecimiento del brote
Contrario a lo esperado, no se detectó un efecto negativo de la formación de estróbilos femeninos sobre el crecimiento total del brote en los árboles que recibieron AG4/7. Aunque se observó un retraso en la velocidad de crecimiento del brote y una reducción en la tasa máxima de crecimiento en los árboles con estróbilos femeninos (Figura 5), la longitud promedio del brote fue similar en los dos grupos de árboles. Estos resultados muestran una posible competición temporal por recursos entre el crecimiento reproductivo y el vegetativo en los árboles que fueron estimulados con AG4/7, pero tal efecto no fue suficiente para afectar negativamente el crecimiento total del brote.
En un estudio con árboles de 9 a 10 años de edad de P. menziesii, en familias con un historial de floración conocido, se encontró que las giberelinas aumentaron en mayor grado la longitud del brote en las familias con menor producción de conos (Webber et al., 1985). En otro estudio, los árboles con estróbilos mostraron menor crecimiento del brote que los que no formaron estróbilos (Bonnet, 1987b). Los estróbilos femeninos pueden competir con el crecimiento del brote por recursos, especialmente en el segundo año cuando están en pleno desarrollo, aunque no siempre limitan su crecimiento (Dick et al., 1990).
Otros autores han indicado que la producción de estróbilos de ambos sexos aumenta con la edad del árbol, mientras que el crecimiento de los brotes se reduce (Greenwood, 1981). En P. menziesii, la respuesta de los brotes al AG4/7 disminuye con la edad, mientras que la floración (en particular masculina) aumenta (Ross, 1983). Sin embargo, determinar el grado de competición entre los estróbilos femeninos y el crecimiento de los brotes es muy complejo, ya que es difícil comparar adecuadamente los estados reproductivos y vegetativos de árboles con diferencias genéticas entre ellos, por el estado fisiológico y por otros factores asociados con la producción de estróbilos (Dick et al., 1990; Ross y Bower, 1991).
Implicaciones para el manejo
El uso de técnicas para estimular la diferenciación de estructuras reproductoras en individuos juveniles de Pseudotsuga menziesii y de otras especies de coníferas que requieren de muchos años para alcanzar la madurez sexual, es cada vez más importante en la producción de semilla de calidad requerida para abastecer los programas de plantaciones forestales en México o para asegurar la conservación de las especies forestales amenazadas.
Los resultados del estudio mostraron que algunos tratamientos tienen efectos colaterales sobre el crecimiento de las ramillas y el desarrollo de la copa. Sin embargo, a diferencia de otros estudios en donde se han encontrado reducciones importantes en el desarrollo de la copa y vigor de los árboles por el uso de AG4/7 o del anillado para estimular floración (Ross y Bower, 1991; Cherry et al., 2007), el principal efecto negativo en este caso fue una ligera reducción en la longitud total del brote y un retraso en su curva decrecimiento, ocasionados por el anillado del tronco y por la competición con los estróbilos. La magnitud del efecto colateral podría aumentar con el uso recurrente de tratamientos de inducción floral, especialmente del anillado en los árboles de menor edad o tamaño.
Para reducir los riesgos de una pérdida de vigor en los árboles por el uso recurrente del anillado es importante considerar otras prácticas de manejo complementarias, como el riego y la fertilización, que reduzcan el nivel y duración del estrés fisiológico o permitan un mayor periodo de recuperación antes de anillar nuevamente el tronco. En el caso del AG4/7 no se encontraron efectos negativos sobre el crecimiento del brote, por lo que se podrían hacer aplicaciones cada año; en este caso se sugiere utilizar la dosis baja (1.27 mg cm-2), ya que a mediano plazo esta dosis ocasionaría menor daño por heridas en el tronco y menor riesgo de efectos tóxicos en los árboles, además del menor costo de aplicación.
CONCLUSIONES
Los tratamientos de AG4/7 y anillado en el tronco que promovieron la formación de estructuras reproductivas también afectaron el crecimiento de los brotes; el anillado redujo la longitud final del brote y la tasa máxima de crecimiento, y retrasó la velocidad de crecimiento. La aplicación de giberelinas, en cambio, aumentó la longitud total y la tasa máxima de crecimiento del brote en los tres ensayos, y aceleró la velocidad de crecimiento, especialmente con dosis bajas de AG4/7. El efecto de los tratamientos fue consistente en los tres ensayos realizados con árboles de diferente edad, a pesar de las diferencias en el crecimiento del brote asociadas con la edad y con las condiciones ambientales de cada ensayo.
En los árboles que recibieron AG4/7 no se encontró un efecto negativo de la formación de estróbilos sobre el crecimiento total del brote, pero sí un retraso en la curva de crecimiento, evidencia de una posible competición temporal por recursos entre ambos procesos durante el primer año de los tratamientos. Debido a lo anterior, el uso repetido del anillado para inducir floración en árboles juveniles de Pseudotsuga menziesii podría afectar negativamente el desarrollo de la copa, pero la aplicación de AG4/7 en dosis bajas no tiene estos efectos colaterales negativos.
AGRADECIMIENTOS
Al Ing. León Jorge Castaños Martínez y al Ing. Salvador Castro Zavala, propietarios del Conjunto Predial Forestal, por todas las facilidades y apoyo logístico proporcionado durante la realización del estudio.
BIBLIOGRAFÍA
Aderkas P, L Kong, S Abrams, I Zaharia, S Owens, B Porter (2004) Flower Induction Methods for Lodgepole Pine and Douglas-fir. Centre for Forest Biology, University of Victoria. 59 p. [ Links ]
Bonnet M M (1987a) Floral induction in conifers: a review of available techniques. For. Ecol. Manage. 19:135-146. [ Links ]
Bonnet M M (1987b) Preliminary results on gibberellins induction of flowering of seedlings and cuttings of Norway spruce indicate some carry-over effects. For. Ecol. Manage. 19:163-171. [ Links ]
Bonnet M M, P Delanzy, G Chanteloup, J Coupaye (1982) Influence de I'état d'activité des raciness sur la floraison induite par des gibberellines 4 et 7 chez Pseudotsuga mensiesii (Mirb.) Franco. Silvae Genet. 31:178-183. [ Links ]
Borchert R (1976) Differences in shoot growth patterns between juvenile and adult trees and their interpretation based on systems analysis of trees. Acta Hort. 56:123-130. [ Links ]
Chalupka W (1980) Regulation of flowering in Scots pine (Pinus sylvestris L.) grafts by gibberellins. Silvae Genet. 29:118-121. [ Links ]
Cherry M L, T S Anekonda, M J Albrecht, G T Howe (2007) Flower stimulation in young miniaturized seed orchards of Douglas-fir (Pseudotsuga menziesii). Can. J. For. Res. 37:1-10. [ Links ]
Dick J McP, R R B Leakey, P G Jarvis (1990) Influence of female cones on the vegetative growth of Pinus contorta trees. Tree Physiol. 6:151-163. [ Links ]
Ebell L F (1971) Girdling: its effect on carbohydrate status and on reproductive bud and cone development of Douglas-fir. Can. J. Bot. 45:453-466. [ Links ]
Greenwood M S (1981) Reproductive development in loblolly pine. II. The effect of age, gibberellins plus water stress and out-of-phase dormancy on long shoot growth behavior. Amer. J. Bot. 68:1184-1190. [ Links ]
Harrison D L S, J N Owens (1994) Gibberellins A4/7 enhanced flowering in Tsuga heterophylla. II. Apical development and shoot formation. Int. J. Plant Sci. 155:302-312. [ Links ]
Ho R H, K Eng (1995) Promotion of cone production on field-grown eastern white pine grafts by gibberellin A4/7 application. For. Ecol. Manage. 75:11-16. [ Links ]
Longman K A, J McP Dick, M Mugglestone, R I Smith (1986) Effects of gibberellins A4+7 and bark-ringing on cone initiation in mature Picea sitchensis grafts. Tree Physiol. 1:101-113. [ Links ]
Muñoz-Gutiérrez L, J J Vargas-Hernández, J López-Upton, N Gutiérrez-Rangel (2010) Inducción de estructuras reproductivas en Pseudotsuga menziesii. Agrociencia 44:835-847. [ Links ]
Owens J N, J E Webber, S D Ross, R P Pharis (1985) Interaction between gibberellins A4/7 and root-pruning on the reproductive and vegetative processes in Douglas-fir. III. Effects on anatomy of shoot elongation and terminal bud development. Can. J. For. Res. 15:354-364. [ Links ]
Pharis R P, C G Kuo (1977) Physiology of gibberellins in conifers. Can. J. For. Res. 7:299-325. [ Links ]
Pharis R P, S D Ross, E E McMullan (1980) Promotion of flowering in the Pinaceae by gibberellins. III. Seedlings of Douglas-fir. Physiol. Plant. 50:119-126. [ Links ]
Philipson J J (1985) The promotion of flowering in large field-grown Sitka spruce by girdling and stem injections of gibberellins A4/7. Can. J. For. Res. 15:166-170. [ Links ]
Philipson J J (1990) Prospects for enhancing flowering of conifers and broadleaves of potential silvicultural importance in Britain. Forestry 63:223-240. [ Links ]
Puritch G S (1972) Cone Production in Conifers. Pacific Forest Research Centre, Canadian Forest Service Inf. Rep. BC-X-65, Victoria BC. 94 p. [ Links ]
Ross S D (1983) Enhancement of shoot elongation in Douglas-fir by gibberellins A4/7 and its relation to the hormonal promotion of flowering. Can. J. For. Res. 13:986-994. [ Links ]
Ross S D, R C Bower (1991) Promotion of seed production in Douglas-fir grafts by girdling + gibberellin A4/7 stem injection, and effect of retreatment. New For. 5:23-34. [ Links ]
Ross S D, R P Pharis (1976) Promotion of flowering in the Pinaceae by gibberellins. I. Sexually mature, non-flowering grafts of Douglas-fir. Physiol. Plant. 36:182-186. [ Links ]
Ross S D, M P Bollmann, R P Pharis, G B Sweet (1984) Gibberellin A4/7 and the promotion of flowering in Pinus radiata. Effects on partitioning of photoassimilate within the bud during primordial differentiation. Plant Physiol. 76:326-330. [ Links ]
SAS Institute (1998) SAS User's Guide: Statistics. Release 6.03 Ed. SAS Institute, Inc. Cary, NC. 1028 p. [ Links ]
Smith R F, M S Greenwood (1997) Effects of cone-induction treatments on black spruce (Picea mariana) current-year needle development and gas exchange properties. Tree Physiol. 17:407-414. [ Links ]
S-Plus (2005) S-Plus 7.0 Guide to Statistics Vol. 1. Insightful Corporation. Seattle, Washigton. [ Links ]
Webber J E, S D Ross, R P Pharis, J N Owens (1985) Interaction between gibberellin A4/7 and root-pruning on the reproductive and vegetative process in Douglas-fir. II. Effects on shoot elongation and its relationship to flowering. Can. J. For. Res. 15:348-353. [ Links ]
Wilson B F, B L Gartner (2002) Effects of phloem girdling in conifers on apical control of branches, growth allocation and air in wood. Tree Physiol. 22:347-353. [ Links ]
Zwieniecki M A, P J Melcher, T S Field, N M Holbrook (2004) A potential role for xylem-phloem interactions in the hydraulic architecture of trees: effects of phloem girdling on xylem hydraulic conductance. Tree Physiol. 24:911-917. [ Links ]

