











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
Salvia L. es el género más diverso de Lamiaceae, con alrededor de 1000 especies (Harley et al., 2004). Este género ha tenido una circunscripción controversial y a partir del auge de los estudios moleculares se descubrió que no era monofilético (Walker et al., 2004; Walker y Sytsma, 2007). La característica morfológica que unificaba a este grupo no natural era la presencia de una estructura espatulada conocida como palanca estaminal, la cual proviene del alargamiento y fusión del conectivo de los dos estambres (Claßen-Bockhoff et al., 2003). La palanca estaminal juega un papel importante en la polinización de Salvia, pues restringe el acceso al néctar a los polinizadores, pero estos al empujarla activan un mecanismo de balancín en el que las tecas depositan granos de polen en la región dorsal de los polinizadores (Claßen-Bockhoff et al., 2004).
En la actualidad, la propuesta más aceptada para abordar la parafilia de Salvia es la de Drew et al. (2017). En ella se plantea ampliar la circunscripción del género para incluir miembros de cinco géneros poco diversos (Dorystaechas Boiss. & Heldr. ex Benth., Meriandra Benth., Perovskia Kar., Rosmarinus L. y Zhumeria Rech.f. & Wendelbo), los cuales están intercalados entre distintos clados de Salvia y carecen de palanca estaminal. Bajo esta propuesta, Salvia se convirtió en un gran género (Frodin, 2004), que es monofilético y se caracteriza por la presencia de conectivos engrosados o alargados (Drew et al., 2017). Al interior de Salvia se reconocen 11 clados y/o subgéneros dentro de los que destaca Salvia subgénero Calosphace (Benth.) Epling, que es endémico de América y engloba a más de 580 especies (González-Gallegos et al., 2020a).
Salvia subgénero Calosphace es un grupo de plantas con una morfología muy variada, tanto vegetativa como reproductiva, que se refleja en la complejidad del grupo y en la complicación de su clasificación infragenérica, la cual ha cambiado a lo largo de su historia (Bentham, 1832, 1848; Briquet, 1897; Epling, 1939). Hasta el momento, la clasificación más aceptada y exhaustiva de Salvia subgénero Calosphace es la de Epling (1939). En ella, y en trabajos posteriores (Epling, 1940, 1941, 1947, 1951, 1960; Epling y Játiva, 1963, 1966, 1968; Epling y Mathias, 1957), se propusieron 102 secciones. No obstante, muchos de los caracteres empleados para delimitar estas agrupaciones (p. ej., presencia de estambres insertos o exsertos o de indumento formado por tricomas ramificados) han resultado ser homoplásicos a la luz de los análisis filogenéticos basados en caracteres moleculares (Jenks et al., 2013; Fragoso-Martínez et al., 2018). Por ello, de las 38 secciones propuestas por Epling en sus diferentes trabajos, y cuya monofilia se ha puesto a prueba, solamente 12 han resultado ser grupos naturales (Fragoso-Martínez et al., 2018). Aún no existe una propuesta formal de actualización de la clasificación infragénerica de Salvia subgénero Calosphace donde se reflejen las relaciones filogenéticas de sus especies. Por lo tanto, considerando que las secciones constituyen unidades morfológicas que facilitan el estudio de este clado altamente diverso, la clasificación de Epling (1939) sigue siendo empleada como una referencia para el estudio y descripción de los taxones del subgénero.
México es el centro de diversidad del subgénero Calosphace (Jenks et al., 2013), con alrededor de 310 especies, 82% de ellas endémicas (Martínez-Gordillo et al., 2017; González-Gallegos et al., 2020a). Esta diversidad continúa incrementándose, pues nuevas especies se describen constantemente; por ejemplo, en los últimos tres años se han descrito cinco especies mexicanas de Salvia (Bedolla-García et al., 2020; González-Gallegos et al., 2020b; Fragoso-Martínez et al., 2021; González-Gallegos et al., 2021a, 2021b). Se piensa que aún existen especies por describir, sobre todo en los estados considerados altamente diversos dentro de la República Mexicana; por ejemplo, Oaxaca, Chiapas, Jalisco, Veracruz y Guerrero (Villaseñor, 2016), estados donde, por su variedad orográfica y problemática social, la exploración es complicada, sobre todo en zonas remotas poco accesibles.
Durante la elaboración del tratamiento florístico de la familia Lamiaceae del estado de Guerrero, se encontraron ejemplares que presentan una singular característica: estambres con filamentos curvados y largamente exsertos. Este trabajo se enfoca en la descripción de este taxón como una especie nueva, y su comparación con especies morfológicamente afines.
Materiales y Métodos
Durante marzo y abril de 2023 se hicieron salidas de campo a zonas poco colectadas de Guerrero, con el fin de documentar su flora. En estas expediciones se colectó un taxón perteneciente al género Salvia subgénero Calosphace, con los estambres exsertos con conectivos curvados, distinto a otras especies conocidas de la zona. Los ejemplares fueron analizados, y las flores disectadas y observadas bajo un microscopio estereoscópico Leica S6 D (Heerbrugg, Suiza), y posteriormente depositados en los herbarios FCME, MEXU y XAL (Thiers, 2023).
Para la identificación de los ejemplares se consultaron las claves disponibles, en particular la de Salvia subgénero Calosphace de Epling (1939), la cual se usó para decidir a qué sección asignar al nuevo taxón. Las comparaciones con especies morfológicamente cercanas se establecieron con base en el material de los herbarios FCME, MEXU, QCNE y XAL (Thiers, 2023), así como en el de los herbarios y bases de datos digitales JSTOR (2023), RHNM (2023) y TROPICOS (2023).
El mapa de distribución se elaboró en QGIS v. 3.30 (QGIS Development Team, 2023), el cálculo de la extensión de ocurrencia (EOO) y el área de ocupación (AOO) se realizó con GeoCAT (Bachman et al., 2011), y la evaluación del estado de conservación se hizo con base en dichos cálculos y tomando en cuenta los criterios de la IUCN (2022).
Resultados
Taxonomía
Salvia ayecarrenoi Mart. Gord., Fragoso & de Santiago, sp. nov. Figs. 1, 2.
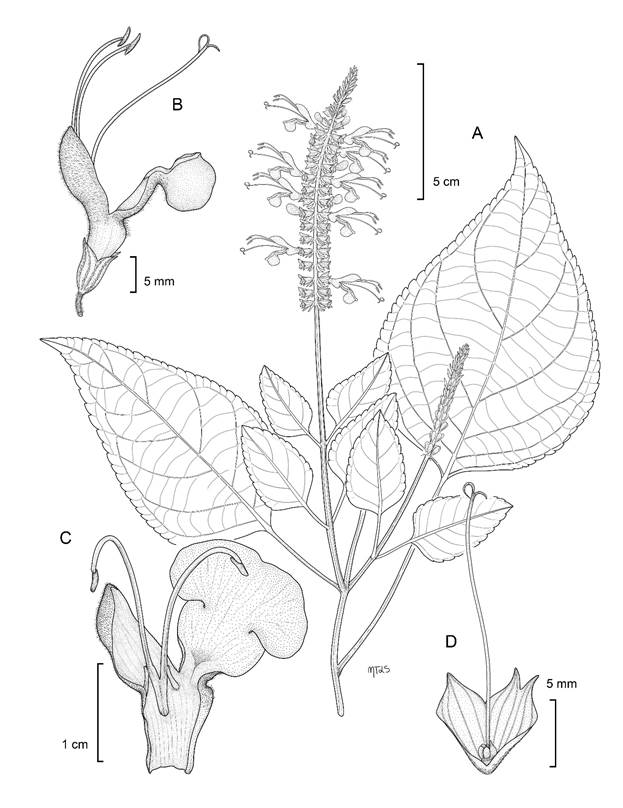
Figura 1: Morfología de Salvia ayecarrenoi Mart. Gord., Fragoso & de Santiago. A. rama con hojas e inflorescencia; B. flor; C. corola disectada con estambres; D. cáliz disectado con mericarpos en desarrollo y estilo. Ilustración de María Teresa Jiménez Segura.

Figura 2: Salvia ayecarrenoi Mart. Gord., Fragoso & de Santiago. A. inflorescencia; B. flor con estambres exsertos y curvos; C. hoja. Fotografías de R. de Santiago.
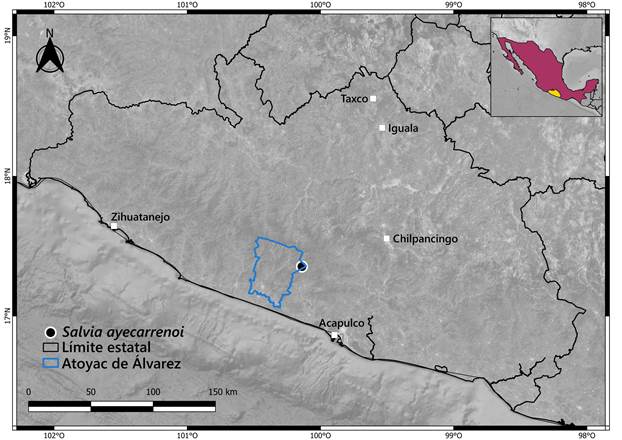
TIPO: MÉXICO. Guerrero, municipio Atoyac de Álvarez, aproximadamente 2.5 km al S de El Edén, 1365 m s.n.m., 17°21'17''N, 100°8'22.5''W, 13.IV.2023, R. de Santiago 4034 (holotipo: FCME!, isotipos: MEXU!, XAL!).
Similar to Salvia sigchosica, from which it can be distinguished by being a suffrutex of 1.5-2.5 m tall, with leaves 3.7-20 cm long; inflorescences of 14-46 cm long; bracts ovate or ovate-elliptic, long, acuminate; calyx with the upper lobe triangular, slightly acuminate; corolla lacking nectar guides, lower lip of the corolla cucullate, and upper stigmatic branch longer than the inferior.
Sufrútices erectos, 1.5-2.5 m de alto; tallos poco ramificados, glabrescentes, con tricomas simples en los ángulos; hojas membranáceas; pecíolos 2-9 cm de largo, glabrescentes, con tricomas simples, cortos, láminas ovadas a ovado-lanceoladas, 3.7-20 × 1.6-11.5 cm, ápice acuminado, base obtusa, menos frecuentemente redondeada, a veces oblicua, margen crenado-serrulado, haz glabrescente, con tricomas simples y cortos sobre las nervaduras, envés glabrescente; inflorescencias laxas, terminales, racemosas, 14-46 cm de largo, 6-8 flores por verticilastro, raquis hirsuto, con tricomas simples, cortos; brácteas ovadas u ovado-elípticas, 4.6-9.1 mm de largo, ápice largamente acuminado, margen entero, glabras en la cara adaxial, hirsútulas en la cara abaxial, con tricomas cortos, por lo general caducas en la antesis; pedicelos 2-7.7 mm de largo, hirsútulos; cáliz verde, 0.8-1.2 cm de largo, tubo ligeramente infundibuliforme, 6.2-7.6 mm de largo, labio superior 3-nervado, triangular, ligeramente acuminado y curvado hacia arriba, 2.6-5 mm de largo, labio inferior dividido en dos lóbulos, 3.4-5.2 mm de largo; corola de color violeta, 0.9-2.1 cm de largo, guías de néctar ausentes, tubo de la corola sin papilas, ligeramente curvado, ventricoso, 0.8-1.2 cm de largo, ligeramente blanquecino en la región proximal, labio superior galeado, 0.9-1.2 cm de largo, recto, densamente pubescente en la superficie externa, tricomas simples, labio inferior 1.1-1.4 cm de largo, trilobado, el lóbulo medio cuculado; botón floral cilíndrico; estambres largamente exsertos, filamentos 3.4-9 mm de largo, glabros, conectivo 1.8-2.2 cm de largo, estaminodios filiformes, 0.8-0.9 mm de largo; estilo 2.6-3.5 cm, glabro, largamente exserto, bilobado, rama superior más larga que la inferior, curvada hacia atrás; disco nectarífero glabro, prolongación del disco ligeramente más larga que los mericarpos, 1.8-2.3 mm de largo, triquetros, marrón-obscuro, glabros, lisos.
Distribución y hábitat: se conoce solamente de una población, que se encuentra en un bosque mesófilo de montaña a 1365 m de altitud en la sierra Madre del Sur, en el estado de Guerrero (Fig. 3). La especie se recolectó en vegetación secundaria, junto con Alchornea latifolia Sw., Dendropanax arboreus (L.) Decne. & Planch., Fuchsia arborescens Sims, Pinus chiapensis (Martínez) Andresen y Ruellia eumorphantha Lindau, entre otras.
Fenología: Salvia ayecarrenoi ha sido colectada en abril con flores y frutos; probablemente su periodo de floración se extienda al menos un par de meses antes y después de abril.
Etimología: la nueva especie está dedicada a Ayesha Carreño Martínez, por su infinita comprensión al trabajo botánico y su apoyo incondicional.
Estado de conservación: Salvia ayecarrenoi está representada por dos colectas realizadas en una zona abierta de un bosque mesófilo de montaña de la Sierra Madre del Sur donde la exploración es peligrosa y, por tanto, solamente se conoce una población formada por un solo manchón con cerca de 20 individuos. En el análisis en GeoCAT (Bachman et al., 2011), la extensión de presencia de la nueva especie (EOO) es de 0 km2; mientras que el tamaño del área de ocupación (AOO) es de 4 km2. Tomando en cuenta estos resultados, junto con los criterios de la IUCN (2022) y los escasos datos del hábitat y población de la especie, sugerimos colocar a S. ayecarrenoi en la categoría Peligro Crítico (CR B1a). La amenaza más inmediata que podría enfrentar Salvia ayecarrenoi es el cambio de uso de suelo.
Otros especímenes analizados: MÉXICO. Guerrero, municipio Atoyac de Álvarez, aproximadamente 2.5 km al S de El Edén, 1365 m s.n.m., 17°21'17''N, 100°8'22.5''O, 13.IV.2023, L. Lucena 1 (FCME, MEXU, XAL).
Discusión
Salvia ayecarrenoi es una especie con características singulares. Se trata de un sufrútice poco ramificado, flores de color violeta, corola con garganta muy abierta, tubo ventricoso y estambres largamente exsertos y curvados. Esta última es una característica inusual en las especies mexicanas de Salvia subgénero Calosphace, ya que la mayoría de los taxones mexicanos con estambres exsertos presentan conectivos rectos (p. ej., Salvia cinnabarina M. Martens & Galeotti, Salvia coccinea Buc'hoz ex Etl., Salvia elegans Vahl, Salvia iodantha Fernald, Salvia iodophylla Epling, Salvia lasiantha Benth. y Salvia longistyla Benth.). La presencia de estambres exsertos en Salvia ayecarrenoi sugiere una posible polinización por aves, pues las especies ornitófilas de Salvia subgénero Calosphace suelen tener palancas estaminales no funcionales y estambres exsertos, mientras que la mayoría de las especies melitófilas presentan palancas funcionales y estambres insertos (Wester y Claßen-Bockhoff, 2007; 2011).
Al seguir la clave de Epling (1939) para las secciones del subgénero Calosphace, Salvia ayecarrenoi se ubicaría en la sección Siphonantha Epling, un grupo de cinco especies sudamericanas, particularmente de Ecuador (Fernández-Alonso, 2006; Fig. 4). Las especies de la sección Siphonantha son arbustos o subarbustos, con inflorescencias racemosas, brácteas caducas, cáliz con el labio superior 3-7-nervado, corola azul obscuro, con el tubo cilíndrico, ventricoso o arqueado, ascendente, sin papilas en el interior, con estambres exsertos y estilo comúnmente glabro (Epling, 1939). De acuerdo con Fernández-Alonso (2006), además de las características antes mencionadas, la presencia de estilos con ramas estigmáticas superiores más cortas que las inferiores también es una característica diagnóstica de Salvia sección Siphonantha. Sin embargo, este carácter no forma parte de la circunscripción original de Epling (1939), puesto que una de las especies que incluyó en la sección, Salvia lobbii Epling, presenta la rama estigmática superior más larga que la inferior (Fig. 4A).

Figura 4: Algunas especies de Salvia sección Siphonantha Epling. A. Salvia lobbii Epling (Ecuador, I. Fragoso-Martínez 799 (QCNE)); B. Salvia pichinchensis Benth. (Ecuador, I. Fragoso-Martínez 634 (QCNE)); C. Salvia sigchosica Fern. Alonso (Ecuador, I. Fragoso-Martínez 732 (QCNE)). Fotografías de G. A. Salazar.
A pesar de que la distribución de Salvia ayecarrenoi no coincide con la de la sección Siphonantha, comparte con sus integrantes varias de las características que la definen, por lo que tentativamente se decidió asignarla a dicha sección, hasta que se amplíe el muestreo taxonómico en la filogenia de Salvia subgénero Calosphace y se revise su clasificación infragenérica. No obstante, es importante señalar que incluir a la nueva especie en Siphonantha podría resultar en una agrupación no natural. Lo anterior podría resultar semejante a aquellas secciones con distribución disyunta entre México y Sudamérica, que se han recuperado como grupos polifiléticos (p. ej., Angulatae Epling, Flocculosae (Epling) Epling y Tomentellae (Epling) Epling), cuyas especies geográficamente cercanas se encuentran emparentadas de una manera más estrecha (Fragoso-Martínez et al., 2018).
Al emplear la clave de identificación más actualizada para la sección Siphonantha (Fernández-Alonso, 2006), la especie con mayor parecido morfológico a Salvia ayecarrenoi es Salvia sigchosica Fern. Alonso (Fig. 4C), un taxón endémico de Ecuador. Ambas especies tienen hojas ovadas, acuminadas, glabrescentes, el labio superior del cáliz 3-nervado, el tubo de la corola sin papilas, la galea vilosa en la superficie externa, estambres exsertos y estilos glabros. Sin embargo, la nueva especie presenta varias características que la diferencian de Salvia sigchosica (Cuadro 1). Entre ellas destacan: las corolas con guías de néctar ausentes, tubo ventricoso y lóbulo medio del labio inferior de la corola cuculado en Salvia ayecarrenoi (vs. corolas con guías de néctar, tubo recto y lóbulo medio del labio inferior de la corola plano en S. sigchosica; Figs. 1B, 4C), los estilos más largos que los estambres (vs. estilos y estambres de tamaño similar; Figs. 1B, 4C), y la rama estigmática superior más larga que la inferior, curvada hacia atrás (vs. rama estigmática superior más corta que la inferior, recta; Figs. 1B, 4C).
Cuadro 1: Comparación morfológica entre Salvia ayecarrenoi Mart. Gord., Fragoso & de Santiago y Salvia sigchosica Fern. Alonso.
| Caracteres | Salvia ayecarrenoi Mart. Gord., Fragoso & de Santiago | Salvia sigchosica Fern. Alonso |
|---|---|---|
| Altura (m) | 1.5-2.5 | 0.4-0.8 |
| Hojas | ||
| Forma | ovadas a ovado-lanceoladas | ovado-lanceoladas a lanceoladas |
| Tamaño (cm) | 3.7-20 × 1.6-11.5 | 3-6 × 2-3.5 |
| Base | obtusa a redondeada, en ocasiones oblicua | cuneada a redondeada |
| Margen | crenado-serrulado | aserrado |
| Longitud del pecíolo (cm) | 2-9 | 1-1.5 |
| Inflorescencia | ||
| Longitud (cm) | 14-46 | 4.5-8.5 |
| Número de flores por verticilastro | 6-8 | 2-5 |
| Brácteas | ||
| Forma | ovadas a ovado-elípticas | ovado-lanceoladas o lanceoladas |
| Longitud (mm) | 4.6-9.1 | 9-11 |
| Pubescencia | hirsútulas | glabrescentes |
| Cáliz | ||
| Longitud (mm) | 6.2-7.6 | 6-9.5 |
| Forma del labio superior | triangular | redondeado |
| Ápice del labio superior | acuminado | mucronado |
| Corola | ||
| Guías de néctar | ausentes | presentes |
| Forma del tubo de la corola | ventricoso | recto |
| Longitud del labio inferior (mm) | 11-14 | 8-10 |
| Forma del botón floral | cilíndrico | piriforme |
| Estambres | ||
| Longitud del conectivo (cm) | 1.8-2.2 | 2.8-3 |
| Forma del conectivo | curvado | ligeramente arqueado |
| Estilo | ||
| Proporción de las ramas estigmáticas | rama superior más larga que la inferior, curvada hacia atrás | rama superior más corta que la inferior, recta |
A continuación, se presenta una clave de identificación para las especies de Salvia sect. Siphonantha, en la que se incluye la nueva especie.
Clave de identificación para las especies de Salvia sect. Siphonantha
1a. Cáliz con el labio superior con 7 nervios …………………………………….…. Salvia lobbii Epling
1b. Cáliz con el labio superior con 3-5 nervios …………………………………………………...…….… 2
2a. Inflorescencias con 12-24 flores por verticilastro; pedicelos 1.5-4 cm de largo; corola color vino …………………………………………………………………………….... S. medusa Epling & Játiva
2b. Inflorescencias con 8 flores o menos por verticilastro; pedicelos 1 cm de largo o menos; corola azul o violeta ………………………………………………………………………………….………..………… 3
3a. Rama superior del estilo más larga que la inferior ……………………………………………………………………………. Salvia ayecarrenoi Mart. Gord, Fragoso & de Santiago
3b. Rama superior de estilo más corta que la inferior ……………………………..…..………………… 4
4a. Hierbas, menores a 1 m de alto; tubo de la corola 1.1-1.4 mm de largo …………………………………………………………….….. Salvia sigchosica Fern. Alonso
4b. Arbustos, 1-3 m de alto; tubo de la corola 2-3.5 cm de largo ……………………………………….. 5
5a. Cáliz 1.4-1.6 cm de largo; tubo de la corola 3-3.5 cm de largo …………. Salvia cyanocephala Epling
5b. Cáliz 1.2-1.3 cm de largo; tubo de la corola 2-2.5 cm de largo ………. Salvia pichinchensis Benth.
La descripción de Salvia ayecarrenoi, un taxón con características inusuales para las especies mexicanas conocidas de Salvia subgénero Calosphace, es un recordatorio de que el estudio de las Lamiaceae del país aún está en proceso, y que la exploración de áreas poco colectadas es vital para avanzar en el conocimiento de la flora mexicana. Estas exploraciones son también imprescindibles para expandir la escasa información sobre la distribución y poblaciones de las nuevas especies como Salvia ayecarrenoi que se encuentra en Peligro Crítico de acuerdo con la IUCN.

