











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
Las selvas tropicales lluviosas han sido severamente perturbadas, transformadas y fragmentadas por diferentes actividades humanas, entre ellas la ganadería (Coates-Estrada y Estrada, 1986; Guevara et al., 1997; Von Thaden et al., 2020; Aguirre-Jaimes et al., 2021). La alteración de estos ecosistemas puede ocasionar la pérdida de hábitat y la disminución de la diversidad faunística, lo que a su vez afecta su funcionamiento (Ceccon y Martínez-Garza, 2016; Levy-Tatcher et al., 2016; Mendoza y Camargo-Sanabria, 2019). Un grupo animal muy diverso, que desempeña roles ecológicos claves para el mantenimiento de la estructura y regeneración natural de la vegetación de estos ecosistemas, es el de los mamíferos. Entre las funciones ecológicas que inciden de diferente manera en la vegetación se pueden mencionar la herbivoría, la polinización, la dispersión de semillas y también la depredación (Dirzo y Miranda, 1990; Miller et al., 2001; Ripple et al., 2014; Mendoza y Camargo-Sanabria, 2019). Sin embargo, frente a determinadas actividades antropogénicas que alteran los ecosistemas bajo ciertas condiciones, algunas especies de mamíferos son más vulnerables (p. ej., especies de talla grande, con tasas reproductivas bajas y ámbitos hogareños amplios), mientras que otras, más resistentes (p. ej., especies omnívoras) (Galetti y Dirzo, 2013; Flores et al., 2014; Rodríguez-Macedo et al., 2014; Azevedo-Ramos et al., 2006; Hernández-Rodríguez et al., 2019; Mendoza y Camargo-Sanabria, 2019).
Este escenario negativo causado por la perturbación, fragmentación y deforestación de la vegetación puede mitigarse con estrategias de restauración ecológica (p. ej., procesos para asistir la recuperación de un ecosistema que se encuentra dañado, degradado o destruido) (SER, 2004; Gann et al., 2019). En última instancia, con la restauración ecológica se busca recuperar tanto la diversidad taxonómica, como la genética, filogenética y funcional, y no únicamente de la vegetación, sino de todos los grupos biológicos que son parte del ecosistema (Ceccon y Martínez-Garza, 2016; López-Barrera et al., 2017). En el proceso de restauración de la vegetación, se espera que también se vayan recuperando aspectos funcionales como son la diversidad faunística y los roles ecológicos que los animales desempeñan. Así, la presencia de la fauna puede ser un factor tanto catalizador como indicador del avance de la restauración ecológica (Newbold et al., 2015; López-Barrera et al., 2017; Ortiz-Lozada et al., 2017; Catterall, 2018). Por ello, se recomienda que en algunas acciones de restauración ecológica se utilicen especies vegetales con una amplia variedad de frutos que resulten atractivos para la fauna (Martínez-Garza y Howe, 2003). Desafortunadamente, en la mayoría de los estudios y proyectos de restauración ecológica que se han desarrollado en México, el estudio de la fauna como parte de la restauración es uno de los componentes menos abordados (López-Barrera et al., 2017).
En el sur de Veracruz, México, en la región de Los Tuxtlas, la vegetación natural dominante es la selva tropical lluviosa. Sin embargo, en esa región ha sido frecuente que los pobladores locales talen la selva para posteriormente sembrar pastos forrajeros y usar esos espacios como pastizales inducidos para el forrajeo de ganado vacuno. En esa zona, seleccionamos en 2006 uno de estos pastizales inducidos en el que se puso en marcha un experimento de restauración ecológica de largo plazo, con la finalidad de evaluar y acelerar el proceso de recuperación de la vegetación y su funcionalidad ecológica en sitios excluidos del forrajeo del ganado, así como estudiar los cambios en el tiempo respecto a diferentes variables ecológicas (p. ej., biomasa de herbáceas, densidad de árboles, diversidad vegetal, entre otras), y recuperar los procesos de dispersión de plantas y el movimiento de animales a través de estos paisajes agropecuarios. En ese sentido, este trabajo se enfocó en determinar, a 12 años de establecido el experimento, cuáles especies de mamíferos terrestres medianos o grandes (≥1 kg de peso), y con qué frecuencia visitan parcelas de restauración experimental con diferente nivel de intervención. Se esperaba en particular, que las parcelas con intervención máxima (p. ej., con plantaciones de árboles nativos), tuvieran más visitas de las especies de mamíferos pequeños y medianos al ofrecer potencialmente más cobertura de protección o alimento (p. ej., fruta carnosa).
Materiales y Métodos
Área de estudio
Este trabajo se llevó a cabo en terrenos de la colonia ganadera Adolfo Ruíz Cortines, dentro de la Reserva de la Biosfera Los Tuxtlas (RBLT), ubicada en el sur del estado de Veracruz, México (Fig. 1). Las características fisiográficas, climáticas y ambientales a detalle pueden ser consultadas en el Programa de Conservación y Manejo de la RBLT (CONANP, 2006). Esta Área Natural Protegida contiene uno de los últimos reductos de selva tropical lluviosa en el país y protege una enorme biodiversidad, pues alberga 15 tipos de vegetación diferentes, compuestos, a su vez, por más de 2600 especies de plantas (CONANP, 2006; Villaseñor et al., 2018). A nivel de fauna, en la zona se han registrado 46 especies de anfibios, 120 de reptiles, 565 de aves y hasta 139 de mamíferos, así como una enorme diversidad de insectos, incluyendo 861 especies de mariposas, 23 de abejas sin aguijón, 133 de libélulas, 118 especies de coleópteros cerambícidos y 164 especies de otros grupos de escarabajos (CONANP, 2006; González-Christen y Coates, 2019).

Figura 1: Localización de la zona de estudio. A. Reserva de la Biosfera Los Tuxtlas (RBLT; polígono en verde) en el estado de Veracruz, México; B. área de estudio dentro de la RBLT (recuadro en negro); C. las 24 parcelas de restauración experimental (indicadas por la flecha blanca), el círculo rojo muestra la ubicación de la Estación de Biología Tropical Los Tuxtlas, del Instituto de Biología, de la Universidad Nacional Autónoma de México (UNAM).
La información actualizada de la mastofauna registrada en la RBLT, incluye 117 especies (Coates et al., 2017; González-Christen y Coates, 2019). De ellas, 17 corresponden a especies de mamíferos medianos o grandes (con individuos de peso ≥1 kg; Benchimol, 2016), mayormente terrestres, que se detectan con mayor probabilidad en cámaras-trampa (Flores et al., 2014; Ríos-Solís et al., 2021). En la zona de la RBLT, la perturbación, deforestación y pérdida de hábitats se debe, sobre todo, al cambio de uso de suelo para uso agrícola y pecuario, y a las actividades silvícolas ilegales, por lo que domina un paisaje con pastizales inducidos, áreas agrícolas y fragmentos de selva tropical lluviosa de diferentes tamaños (CONANP, 2006). Este deterioro de la cobertura forestal en la RBLT ha disminuido notablemente en las primeras décadas de este siglo, en comparación con lo que pasó entre 1976 y 1986, lo que puede deberse a que cada vez queda menos cobertura forestal accesible o fuera de zonas protegidas (Von Thadden et al., 2020).
Diseño del experimento
Previo al inicio del experimento de restauración ecológica, en los terrenos de estudio, los habitantes de la colonia ganadera Adolfo Ruíz Cortines habían removido completamente la cobertura forestal para inducir pastizales y desarrollar actividades ganaderas. En 2006 se logró un acuerdo con el propietario de un predio de 12 ha para desarrollar un experimento de restauración ecológica a largo plazo. En ese pastizal, se establecieron 24 parcelas (de 30 × 30 m, separadas entre sí 35 m) que se cercaron para impedir el acceso del ganado y permitir el proceso de sucesión natural. En ocho parcelas, el tratamiento de intervención fue mínimo (solamente se cercaron, llamadas en adelante de sucesión natural). En las 16 restantes, la intervención fue máxima, con dos tratamientos de composición de plantación: 1) en ocho se plantaron plántulas (de 30-40 cm de altura) de 12 especies arbóreas nativas dispersadas por viento (llamadas de ahora en adelante plantaciones viento), y 2) en las otras ocho parcelas, plántulas (de 30-40 cm de altura) de 12 especies dispersadas por vertebrados (llamadas de ahora en adelante plantaciones animales).
Entre las especies que se plantaron, ocho se consideran pioneras (especies sucesionales tempranas de rápido crecimiento; cuatro de ellas dispersadas por viento y cuatro por animales) y 16 no-pioneras (especies sucesionales tardías de lento crecimiento; la mitad de ellas dispersadas por viento y la otra mitad por animales; Cuadro 1; Martínez-Garza et al., 2009, 2013; Rivas-Alonso et al., 2021). Las parcelas están dispuestas en una gradilla de tres columnas, ocho filas, de modo que los tres tratamientos están presentes en cada fila de parcelas (Fig. 2). Desde el inicio del experimento, en el 2006, dentro de cada parcela se establecieron cuatro subparcelas (cada una de 169 m2), separadas por dos transectos de 2 m de ancho, a manera de pasillos centrales, en forma de cruz (Fig. 2). Esto con la finalidad de permitir el paso por el interior de la parcela, para facilitar los muestreos y la medición de diferentes variables ecológicas, así como para el establecimiento de las redes para el monitoreo de aves y murciélagos. Algunas características de las parcelas cercadas, como su tamaño, ubicación y separación, se deben, en parte, a limitantes logísticas como el tamaño y la porción del pastizal donde el propietario del predio permitió que se realizara el proyecto.
Cuadro 1: Familia, historia de vida y síndrome de dispersión de 24 especies de árboles nativos plantados en 24 parcelas de restauración ecológica en junio de 2006 en la Reserva de la Biosfera Los Tuxtlas, Veracruz, México (Martínez-Garza et al., 2013).
| Especie | Familia | Historia de vida |
| Dispersión por viento | ||
| Albiza purpusii Britton & Rose | Fabaceae | No-pionera |
| Aspidosperma megalocarpon Müll. Arg. | Apocynaceae | No-pionera |
| Bernoullia flammea Oliv. | Malvaceae | No-pionera |
| Cedrela odorata L. | Malvaceae | Pionera |
| Ceiba pentandra (L.) Gaertn. | Malvaceae | Pionera |
| Cordia megalantha S.F. Blake | Boraginaceae | No-pionera |
| Heliocarpus appendiculatus Turcz. | Malvaceae | Pionera |
| Lonchocarpus guatemalensis Benth. | Fabaceae | No-pionera |
| Ochroma pyramidale (Cav. ex Lam.) Urb. | Malvaceae | Pionera |
| Platymiscium pinnatum (Jacq.) Dugand | Fabaceae | No-pionera |
| Tabebuia guayacan (Seem.) Hemsl. | Bignoniaceae | No-pionera |
| Vochysia guatemalensis J. D. Smith | Vochysiaceae | No-pionera |
| Dispersión por animales | ||
| Amphitecna tuxtlensis A.H.Gentry. | Bignoniaceae | No-pionera |
| Brosimum alicastrum Sw. | Moraceae | No-pionera |
| Cecropia obtusifolia Bertol. | Cecropiaceae | Pionera |
| Cojoba arborea (L.) Britton & Rose | Mimosaceae | No-pionera |
| Dussia mexicana (Standl.) Harms | Fabaceae | No-pionera |
| Ficus yoponensis Desv. | Moraceae | Pionera |
| Guarea grandifolia DC. | Meliaceae | No-pionera |
| Inga sinacae M. Sousa & Ibarra- Manríquez | Fabaceae | No-pionera |
| Poulsenia armata (Miq.) Standl. | Moraceae | No-pionera |
| Pouteria sapota (Jacq.) H.E. Moore & Stearn | Sapotaceae | No-pionera |
| Rollinia jimenezii Saff. | Annonaceae | Pionera |
| Tabernaemontana donnell-smithii Rose | Annonaceae | Pionera |
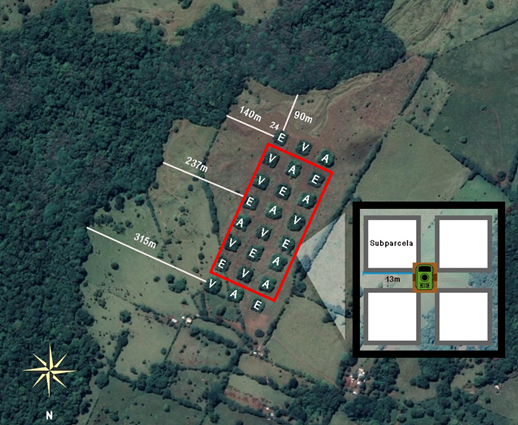
Figura 2: Parcelas de muestreo y sus tratamientos: E: sucesión natural, V: parcelas con plantaciones de especies dispersadas por viento, A: parcelas con plantaciones de especies dispersadas por animales. Se excluyeron las dos filas de tres parcelas, fuera del recuadro rojo, para evitar sesgos, pues una es la fila más cercana a las casas (la más al norte) y la otra la más cercana al fragmento de selva conservado más cercano (la más al sur). Las líneas blancas muestran la distancia al fragmento de selva tropical lluviosa continua más cercano. En el recuadro negro se muestran los pasillos al interior de cada parcela y la ubicación de cada cámara de fototrampeo en ella.
Diseño de muestreo
Para la obtención de datos sobre las especies de mamíferos medianos que visitaron las parcelas, se usaron cámaras-trampa, cuyos detalles se especifican más adelante. Estas se colocaron en 18 de las 24 parcelas (seis por tratamiento), descartando la primera y última fila de la gradilla de cuadros cercados, para evitar sesgos, pues una es la fila más cercana a las casas y la otra la más cercana al fragmento de selva tropical lluviosa conservado más cercano (Fig. 2). Se colocó una cámara-trampa en el tronco de un árbol, a una altura de 60 cm del suelo, al centro de cada una de las 18 parcelas estudiadas, orientándolas hacia la intersección de los pasillos, con el fin de aumentar el campo de captura (Fig. 2). Esta colocación de las cámaras buscaba maximizar el registro de mamíferos medianos terrestres, lo que seguramente implicó un subregistro de mamíferos arborícolas o de pequeños mamíferos (p. ej., roedores).
Se utilizaron tres modelos de cámaras-trampa: Stealth Cam PX14 (Stealth Cam®, Grand Prairie, Texas, EUA), y Cuddeback Attack modelos IR 1156 y Black Flash 1231 (Cuddeback®, Non Typical, Inc., Park Falls, Wisconsin, EUA). Estas fueron programadas para tomar tres fotografías y un vídeo con duración de 30 s por cada evento de detección, con un intervalo mínimo de 30 s entre cada uno y operando continuamente las 24 h del día durante todo el tiempo de muestreo. Todas fueron colocadas el mismo día y se dejaron operando por 10 días consecutivos por periodo de muestreo, uno durante la temporada de lluvias, en septiembre de 2018 y otro en la temporada seca, en mayo de 2019. Al finalizar cada periodo de muestreo, se retiraron las cámaras para revisar sus tarjetas de memoria y obtener las capturas fotográficas de cada parcela.
Preparación de base de datos
Cada tarjeta de memoria se revisó, identificando la especie de los individuos capturados en fotografía y video con ayuda de guías de referencia (Coates-Estrada y Estrada, 1986; Guevara et al., 1997; Aranda, 2012). Esos datos se usaron para construir una base de datos de las fotocapturas en ambos muestreos, usando el programa Wild ID v. 0.9.28 (Fegraus et al., 2011; TEAM, 2017; Thomson et al., 2018). En esa base de datos, cada fotografía y video tiene datos de fecha y hora de captura, especie identificada, así como clave de la parcela en la que estaba la cámara. Con esos datos se determinó el número de especies que visitó cada parcela y la tasa de registro fotográfico, que se calcula al dividir el número de foto registros independientes (p. ej., con al menos 24 h de intervalo entre fotos de la misma especie en la misma cámara) / esfuerzo de muestreo (número de cámaras automáticas operando por el total de días de muestreo) multiplicado por 100 para estandarizar el valor a ese esfuerzo de muestreo (Lira-Torres y Briones-Salas, 2012).
Resultados
El esfuerzo de muestreo total esperado fue de 360 días-trampa; por fallos en el funcionamiento de las cámaras, solo 16 operaron correctamente en el periodo lluvioso y 14 en el periodo seco. Por tanto, se logró un esfuerzo de muestreo total de 290 días-trampa.
Para el periodo de lluvias se obtuvieron 13 fotografías y cuatro vídeos de tres especies de mamíferos silvestres. Para la época seca se lograron 11 fotografías y cinco vídeos con registro de una especie de mamífero silvestre. En total, en 34 fotografías se registraron tres especies de mamíferos silvestres de talla media pertenecientes a tres órdenes y tres familias distintas (en 24 de las 35 fotos): armadillo común (Dasypus novemcinctus Linnaeus, 1758), tlacuache (Didelphis spp.) y guaqueque mexicano (Dasyprocta mexicana Saussure, 1905) (Fig. 3). Además, se obtuvieron 11 registros fotográficos de tres especies domésticas (que no eran el objetivo de nuestro estudio): perro (Canis lupus familiaris Linnaeus, 1758), cabra (Capra aegagrus hircus Linnaeus, 1758) y borrego (Ovis orientalis aries Linnaeus, 1758).

Figura 3: Especies de mamíferos silvestres registradas. A. Dasypus novemcinctus Linnaeus, 1758; B. Dasyprocta mexicana Saussure, 1905; C. Didelphis spp.
Dada la poca cantidad de registros de mamíferos silvestres medianos y ninguno grande en las parcelas de muestreo, aunado a que, en ambas temporadas de muestreo, en la mayoría de las parcelas no se obtuvieron registros de mamíferos, no fue posible realizar un análisis estadístico. Los pocos registros obtenidos no permiten inferir diferencias entre tratamientos y temporadas de muestreo. Por tanto, solo se presentan datos descriptivos, pese a ello, es información valiosa que documenta parte del proceso de recuperación del movimiento de mamíferos pequeños y medianos en este tipo de paisajes agropecuarios, información que no es frecuentemente documentada en proyectos de restauración ecológica.
En el muestreo del periodo lluvioso se registraron las tres especies de mamíferos silvestres antes mencionadas, y en el muestreo del periodo seco solo al armadillo. Este último, en cuatro parcelas de las plantaciones viento, durante el muestreo del periodo lluvioso y en el muestreo del periodo seco, en dos parcelas, una de sucesión natural y otra de las plantaciones viento. El mayor valor promedio de frecuencia relativa de captura fotográfica fue para el armadillo, en sucesión natural en el periodo seco, lo cual se debe a que en la única parcela en la que se fotografió, obtuvimos ocho registros independientes (p. ej., con al menos 24 h de intervalo entre fotos de la misma especie en la misma cámara). Del guaqueque mexicano y el tlacuache solo obtuvimos fotografías en el muestreo del periodo lluvioso. El primero en una parcela de las plantaciones animales, y el segundo en una parcela de las plantaciones animales y en una de las plantaciones viento. En lluvias no se obtuvieron registros de mamíferos silvestres en las parcelas de sucesión natural, mientras que se registraron dos especies de mamíferos en las plantaciones animales y dos especies de mamíferos en las plantaciones viento. En el muestreo del periodo seco, se registró una especie en las parcelas bajo sucesión natural y otra en las plantaciones viento.
Los valores promedio de frecuencia relativa de captura fotográfica por parcela, para cada especie de mamíferos registrada, presentan una gran variabilidad entre tratamientos y temporadas de muestreo (Cuadro 2), sin una tendencia clara. Lo más evidente es que en el periodo lluvioso se registraron dos especies más de mamíferos que en el periodo seco y que el armadillo tiene valores de frecuencia relativa de captura mayores que los de las otras dos especies. En conjunto, como se esperaba, se obtuvieron más registros de mamíferos silvestres en las plantaciones de restauración (intervención máxima), que en la sucesión natural (intervención mínima), y esto fue más evidente en el periodo lluvioso del año. Durante el periodo seco, esta diferencia entre plantaciones y sucesión natural no es tan evidente, e incluso parece a la inversa, pero sobre todo porque en la única parcela bajo sucesión natural donde se registró el armadillo, se obtuvo un valor elevado de frecuencia relativa de captura fotográfica.
Cuadro 2: Frecuencia relativa de captura promedio (± Desviación estándar) por parcela de muestreo para cada especie de mamífero mediano registrada en cada tratamiento de restauración en Los Tuxtlas, Veracruz, México (SN=sucesión natural; A=plantaciones animales; V=plantaciones viento) y temporada de muestreo. Armadillo común (Dasypus novemcinctus Linnaeus, 1758), guaqueque mexicano (Dasyprocta mexicana Saussure, 1905), tlacuache (Didelphis spp.).
| TEMPORADA | TRATAMIENTO | Armadillo común | Guaqueque mexicano | Tlacuache |
| SN | 0 (±0.0) | 0 (±0.0) | 0 (±0.0) | |
| Luvias 2018 | A | 0 (±0.0) | 2 (±4.5) | 2 (±4.5) |
| V | 6.7 (±5.2) | 0 (±0.0) | 1.7 (±4.1) | |
| SN | 22.5 (±45) | 0 (±0.0) | 0 (±0.0) | |
| Secas 2019 | A | 0 (±0.0) | 0 (±0.0) | 0 (±0.0) |
| V | 2 (±4.5) | 0 (±0.0) | 0 (±0.0) |
Discusión
De un total de 17 especies de mamíferos terrestres medianos y grandes que potencialmente se podrían detectar con cámaras-trampa en la zona de la RBLT (Flores et al., 2014; Ríos-Solís et al., 2021), se logró el registro de tres especies, aunque se obtuvieron pocas capturas fotográficas de ellas. Es importante aclarar que en este experimento de restauración no se buscaba per se la recuperación de la diversidad de especies animales, ya que las parcelas son muy pequeñas y difícilmente se les podría equiparar a un fragmento grande de selva tropical lluviosa. Sin embargo, lo que sí se esperaba, de manera particular, era que al progresar la recuperación de la vegetación en las parcelas y por la producción de frutos y semillas, estas pudieran incentivar el movimiento de fauna silvestre entre los fragmentos de selva conservada, al usar las parcelas como peldaños o “stepping stones” (p. ej., sitios cercanos, pero sin conexión entre ellos que facilitan el tránsito de especies entre espacios discontinuos; Baum et al., 2004) para cruzar el paisaje agropecuario y ello se debería reflejar en un mayor registro de especies. Por otra parte, también es importante destacar que, en general, no está bien documentado el cambio en el tiempo en el registro de visitas de las especies de fauna nativa silvestre y la recuperación de sus funciones ecológicas (entre ellas los mamíferos medianos o grandes) en sitios bajo proceso de restauración ecológica (p. ej., sitios de selva tropical lluviosa transformados a pastizales y después libres del pastoreo para buscar el restablecimiento de la vegetación; Howe y Davlantes, 2017). El presente trabajo fue una primera aproximación a esto, enfocada en los mamíferos terrestres medianos presentes en la RBLT.
Un resultado notable de este estudio es que, a 12 años de iniciado el experimento, el registro de la actividad de mamíferos terrestres medianos o grandes en las parcelas experimentales fue muy bajo, en comparación con muestreos con foto-trampeo hechos en fragmentos de selva conservada de la zona. En ellos, Ríos-Solís et al. (2021) utilizando sitios de muestreo de 9 km2, lograron un esfuerzo de muestreo total de 1100 días-trampa y 308 registros (independientes) de 13 especies de mamíferos medianos y grandes. En el presente estudio con 290 días-trampa, se lograron solamente 24 registros independientes de tres especies de mamíferos silvestres en las parcelas experimentales, mismas que se registraron pocas veces y no en la mayoría de las parcelas, independientemente del tratamiento.
A pesar de las diferencias en diseño de muestreo (p. ej., en el esfuerzo de muestreo y en la escala espacial) entre los estudios, el trabajo de Ríos-Solís et al. (2021) es un referente de las especies que se pueden registrar con foto-trampeo en la selva conservada cercana a nuestras parcelas, y por ello se considera que, a pesar del tiempo transcurrido, aún hay especies de mamíferos terrestres medianos presentes en la selva, pero que no están visitando las parcelas o usándolas para cruzar el paisaje agropecuario. Esto puede deberse a las diferencias entre especies en su propensión a cruzar matrices que son estructuralmente diferentes a su hábitat natural, a la distancia entre los parches de hábitats o al tamaño de los mismos (Martines y Toppa, 2018; Beltrán-Lacouture y Howe, 2020). El rango perceptual de un animal (p. ej., la distancia máxima desde la que un animal puede percibir un elemento del paisaje que le facilite orientarse) varía entre especies y se relaciona con su disponibilidad para cruzar un ambiente alterado (Rocha et al., 2021).
Las respuestas de cada especie a la fragmentación se relacionan con diferencias entre ellas, en sus capacidades para percibir la estructura y la escala de la fragmentación a nivel de paisaje (Gehring y Swihart, 2003). Los mamíferos más grandes y con mayor capacidad de movimiento, pueden percibir los ambientes alterados, como el pastizal inducido donde se ubican las parcelas experimentales estudiadas, como más homogéneos y pueden además cruzarlos más fácilmente (Gehring y Swihart, 2003), por lo que podrían no usar o visitar con frecuencia los pequeños elementos de hábitat que sirven como conectores, como las parcelas experimentales. Por otra parte, los animales más grandes pueden tener un rango perceptual mayor, probablemente por tener una posición de los receptores sensoriales (de olfato, visión y audición) más elevada con respecto al suelo y podrían orientarse más fácilmente en ambientes alterados, pero también pueden percibir más fácilmente riesgos de depredación y encontrar poca cobertura vegetal de protección en los pastizales, que los animales más pequeños (Prevedello et al., 2011; Rocha et al., 2021).
Las parcelas experimentales pueden aumentar el rango perceptual de los animales, en particular cuando la vegetación en ellas y su contraste con el entorno alterado va aumentando. Se sabe que, en las parcelas de este experimento en particular, la recuperación de la vegetación se aceleró con el establecimiento y acumulación de árboles y arbustos de sucesión tardía dispersadas por animales (de la Peña-Domene et al., 2013), lo que contrasta con la matriz de pastizal para forrajeo por ganado, donde no hay establecimiento de vegetación típica de selva tropical lluviosa. Este aumento de vegetación natural en las parcelas, incluyendo especies que producen fruta carnosa, ha favorecido el registro en las parcelas, de animales frugívoros voladores como aves y murciélagos (Howe, 2017; Rivas-Alonso et al., 2021).
Para el caso de los roedores pequeños en las mismas parcelas experimentales, durante los primeros seis años del experimento fue notorio un aumento progresivo de la cantidad promedio de capturas por parcela (aunque en más de 90% de solo una especie de roedor), seguido de un decremento progresivo de la especie dominante y con un incremento incipiente de otras especies de roedores en los siguientes cuatro años (Howe y Davlantes, 2017). Ese patrón coincide con un aumento paulatino en la cobertura leñosa por parcela, que se aceleró a partir del sexto año del experimento, sobre todo en las parcelas con plantaciones (Howe y Davlantes, 2017). En nuestro muestreo no registramos roedores pequeños, muy probablemente debido a que el foto-trampeo no es una técnica adecuada para este grupo.
Un experimento a largo plazo de restauración ecológica como este, iniciado en 2006 en la RBLT, ofrece una oportunidad única para observar cómo avanza la sucesión ecológica y el efecto de esta restauración sobre la fauna que puede utilizar las parcelas establecidas para moverse entre fragmentos de selva tropical lluviosa continua a través de un paisaje agropecuario permanente. Se sabe que conforme se da el crecimiento de la vegetación, aumenta la disponibilidad de perchas y alimento para la fauna silvestre (Martínez-Garza et al., 2009). Estas “islas” de vegetación ubicadas entre los remanentes de selva (Figs. 1, 2) pueden ofrecer sitios de paso y refugio para diferentes especies de fauna, por ejemplo, aves y murciélagos, los que a su vez aportan conectividad reproductiva a las especies vegetales que se encuentran en los parches (de la Peña-Domene et al., 2014). Por lo tanto, pueden ayudar a mantener una porción significativa de la diversidad local (Ortiz-Lozada et al., 2017). Sin embargo, la habilidad de un individuo de moverse entre parches de vegetación depende de factores abióticos y bióticos como la distancia a recorrer y la exposición de las áreas entre parches, la capacidad de dispersión de las especies vegetales, la disponibilidad de recursos en cada parche y la posibilidad de interacciones competitivas o el riesgo de depredación (Estrada et al., 1993; Howe, 2014; Beltrán-Lacouture y Howe, 2020).
La expectativa es que las parcelas faciliten el movimiento de la fauna silvestre a través del paisaje perturbado y no necesariamente que permitan su establecimiento en ellas. Sin embargo, también hay evidencia de que estas parcelas pueden ya servir como hábitat y zonas de refugio para algunas especies, pues algunos individuos de armadillo ya tienen madrigueras, aunque ello no ha pasado desapercibido por los habitantes locales que ahora buscan aprovechar esto, llevando perros que obligan a salir a los armadillos para poder cazarlos (Beltrán-Lacouture, 2021). También se registró la presencia de fauna doméstica en las parcelas, pero esto puede tener algunos efectos negativos para los mamíferos nativos medianos, como podría ser la depredación, pues se sabe que este es un efecto causado por los perros ferales o de libre movimiento (Gompper, 2013), o incluso, un escenario en que los pobladores locales explotan para cazar fácilmente, como se mencionó anteriormente.
Estos experimentos de largo plazo pueden permitir explorar cómo se da el recambio de especies faunísticas a lo largo del tiempo (aunque solo sea en cuanto a la visita de las parcelas), y si diferentes tratamientos de restauración afectan y de qué forma, este recambio. Es importante considerar que la restauración ecológica se necesita para disminuir los efectos negativos de la perdida, degradación y fragmentación de los hábitats naturales sobre la fauna, pero la restauración no siempre cumple este objetivo para todos los grupos faunísticos (Hale et al., 2020). Por otra parte, el monitoreo de experimentos de restauración ecológica por lo común se concentra en evaluar el cambio en la cobertura, estructura y diversidad vegetal, asumiendo que las poblaciones de especies animales y las interacciones entre plantas y animales se recuperarán con el tiempo (Hale et al., 2019). Por ello es tan importante ir más allá de supervisar la recuperación de la vegetación al dar seguimiento a la respuesta de diferentes grupos faunísticos ante las acciones de restauración.
Implicaciones para la restauración
Los registros obtenidos sugieren una mayor presencia de especies de mamíferos terrestres medianos en las parcelas con plantaciones que en las parcelas sin plantación (sucesión natural), esto fue más evidente en el periodo lluvioso del año. Las parcelas con plantaciones tienen más cobertura de herbáceas y de plantas leñosas, en particular de especies de estados sucesionales tardíos que las que no tienen plantaciones (Howe y Davlantes, 2017; Rivas-Alonso et al., 2021).
A pesar de que los resultados obtenidos no son concluyentes, se considera que, al continuar la recuperación de la vegetación en el tiempo, se incrementará la diversidad y la frecuencia de visita de mamíferos medianos a estas parcelas, ya sea porqué encuentren más recursos o refugio en ellas, o porqué, al ser más contratantes con el entorno perturbado, faciliten más su uso como sitios de paso para moverse entre fragmentos continuos de selva tropical lluviosa. Es claro que diversas especies de mamíferos medianos o grandes tienen diferentes necesidades ambientales que se cubren a diversas escalas espaciales y temporales. Se sabe que, en ambientes transformados en proceso de sucesión natural, la riqueza faunística en general no llega a ser semejante a la de una selva tropical lluviosa madura sino hasta 20-40 años después de que inició la recuperación de la vegetación y que, al aumentar el tiempo de recuperación de la vegetación, es más claro un cambio en la biomasa total de mamíferos y en el peso promedio por especie, que en la riqueza de especies (Dunn, 2004; Derhé et al., 2018).
El monitoreo de zonas bajo restauración ecológica necesita evaluar si diferentes grupos de fauna utilizan los hábitats en proceso de restauración y si eso puede afectar positivamente su sobrevivencia y el crecimiento de sus poblaciones. Identificar cuándo puede ocurrir este efecto positivo tras el inicio de las acciones de restauración, permite obtener información acerca de los patrones temporales y el tipo de respuestas que diferentes grupos faunísticos pueden presentar (Hale et al., 2020).
Las parcelas experimentales estudiadas, poco a poco se van transformando en pequeños parches de selva tropical, incrementando la heterogeneidad ambiental del pastizal en el que se ubican y ya funcionan como sitios de paso, refugio temporal y/o sitios de alimentación para algunos grupos faunísticos (Howe y Martínez-Garza, 2014) y con ello podrían facilitar el mantenimiento local de metapoblaciones de plantas y animales (Vandermeer y Carvajal, 2001).
Que a 12 años de iniciado el experimento, con cambios importantes en la vegetación dentro de las parcelas, el registro de mamíferos medianos en su interior sea escaso y de pocas especies, revela información valiosa sobre cómo la recuperación de la funcionalidad asociada con la fauna de mamíferos silvestres pequeños y medianos puede avanzar más lentamente que la recuperación de características estructurales o funcionales de la vegetación. El monitoreo de la fauna silvestre y sus respuestas ante el avance de la restauración ecológica también permite identificar situaciones que ameritan acciones de manejo específicas. Tal es el caso de los hábitats en proceso de restauración que causan un aumento no deseado de especies invasoras, debido a que pueden tener características que les hacen más eficientes que las especies nativas, para usar o colonizar hábitats nuevos (Hale et al., 2020). Finalmente es importante enfatizar que el monitoreo de la presencia de mamíferos medianos en estas parcelas debe continuar e intensificarse.

