











 nova página do texto(beta)
nova página do texto(beta) Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
Permalink
Introducción
La familia Asteraceae o Compositae se caracteriza por presentar inflorescencias pseudánticas; es decir, que simulan una flor. Esas inflorescencias se conocen comúnmente como cabezuelas o capítulos, las cuales están conformadas por una o numerosas flores dispuestas sobre un receptáculo rodeado por un involucro de brácteas llamadas filarios, que simulan un cáliz. Cada cabezuela contiene una o numerosas flores gamopétalas epíginas con un cáliz modificado en un vilano, rara vez ausente, y estambres sinántereos, siendo este último su principal carácter diagnóstico (Funk et al., 2009). Es un grupo megadiverso: cálculos recientes estiman la existencia de 25,000 a 30,000 especies equivalentes a 10% de las Angiospermas conocidas a nivel mundial (Mandel et al., 2019). Actualmente sus miembros se agrupan en 16 subfamilias y 50 tribus (Susanna et al., 2020). Esas cifras contrastan con el número de grupos infrafamiliares reconocidos hasta finales del siglo XX, cuando se empleaba la clasificación de Bentham (1873) en la que las asteráceas estaban agrupadas en 13 tribus.
Las primeras clasificaciones de Asteraceae datan de épocas prelinneanas; desde entonces los caracteres florales y de las cabezuelas han sido fundamentales para reconocer grupos. Por ejemplo, Tournefort diferenció tres clases con base en los tipos de cabezuelas: 1) discoides y posiblemente disciformes, 2) únicamente con flores liguladas y 3) con flores radiadas en la periferia y discoides en el centro (Bonifacino et al., 2009). Mientras que Vaillant utilizó características de los filarios, el receptáculo y el vilano para identificar tres grupos: Cardueae, Cichorieae y Corymbiferae (Bonifacino et al., 2009).
Cassini (1816))propuso la primera clasificación tribal de Asteraceae. Él consideró que las características del estilo, el estigma, la corola y las cipselas eran las mejores para distinguir tribus. Además, reconoció tres grupos que serían equivalentes a algunas subfamilias actuales: Lactucées, Astérées y Carduacées; en ellos agrupó a las 19 tribus que reconoció en su propuesta de clasificación (Cassini, 1819). Bentham (1873) empleó los mismos criterios que Cassini, pero solo aceptó 13 tribus, aunque segregó a Helenieae de Heliantheae con base en la ausencia de páleas en el receptáculo. Small (1919) utilizó caracteres del estilo (forma, longitud y pubescencia) y las anteras (presencia o ausencia de apéndices apicales, cauda y collar), a los que en conjunto denominó mecanismo de presentación secundaria del polen, para distinguir a las tribus Anthemideae, Calenduleae, Cichorieae, Heliantheae, Senecioneae y Vernonieae.
Cronquist (1955), estableciendo algunas diferencias en cuanto a relaciones y circunscripción con respecto a la propuesta de Bentham (1873), planteó una clasificación en la que incluyó 12 tribus, pues consideró a Heliantheae filogenéticamente más cercana a Eupatorieae que a Vernonieae e incluyó a Helenieae en Heliantheae argumentando que la ausencia de páleas en el receptáculo no era un carácter relevante para segregarla como un grupo independiente. Carlquist (1976) coincidió con las ideas de Cronquist y las plasmó en una propuesta en la que reconoció dos subfamilias, cada una con seis tribus. En Cichorioideae agrupó aquellas tribus con cabezuelas homógamas discoides y anteras caudadas, mientras que en Asteroideae incluyó las que presentaban cabezuelas radiadas y anteras ecaudadas.
Cronquist (1977) aceptó la clasificación de Carlquist (1976) y, con base en los tipos de flores presentes en las cabezuelas, separó a las compuestas en tres subfamilias, aunque solo asignó nombre a una de ellas. En la primera agrupó a las tribus con cabezuelas discoides cuyas flores tenían corolas tubulares actinomorfas, además de Mutisieae, con corolas bilabiadas. En la segunda incluyó aquellas tribus con cabezuelas heterógamas, con flores radiadas dispuestas en la periferia y tubulares en el centro. En la tercera, que correspondía a Lactuceae, reunió géneros y especies que presentaban cabezuelas con flores liguladas exclusivamente.
Los cambios más sustanciales en la clasificación de la familia ocurrieron con el inicio de estudios filogenéticos en grupos de divergencia temprana y a la par de la inclusión de microcaracteres, datos citogenéticos y moleculares (Bremer, 1987, 1996; Jansen y Palmer, 1987; Bremer y Jansen, 1992; Bremer et al., 1992; Karis et al., 1992). Las modificaciones se vieron reflejadas en la propuesta de Panero y Funk (2002), quienes circunscribieron cinco subfamilias y ocho tribus nuevas segregadas de Cichorioideae y Mutisieae. Años después reconocerían a otras tres subfamilias nuevas (Panero y Funk, 2008). En Asteroideae (la subfamilia que agrupa el mayor número de tribus y por tanto de géneros y especies de asteráceas), también se realizaron diversos trabajos enfocados a aclarar la validez de algunos grupos (Karis, 1993ª, 1993b, 1996); siendo la tribu Heliantheae s.l. una de las más estudiadas. En uno de los múltiples análisis enfocados a esclarecer sus relaciones filogenéticas, Karis (1993b) encontró que en Heliantheae s.l. se distinguían algunos grupos que podían elevarse a rango tribal: Tagetes L., Coreopsidinae y Madiinae. Posteriormente, Baldwin et al. (2002) emplearon datos moleculares y microcaracteres para validar a las tribus de la alianza Heliantheae, formalizando así la circunscripción de Helenieae, Madieae y Tageteae, además de proponer tres nuevas tribus: Bahieae, Chaenactideae y Perityleae. De las 50 tribus que se reconocen actualmente, 11 han sido segregadas de Heliantheae s.l. y 17 de Cichorieae s.l. y Mutisieae s.l. (Funk et al., 2009; Susanna et al., 2020).
Asteraceae es uno de los grupos más fáciles de identificar a nivel familia debido, principalmente, a sus inflorescencias primarias que son las cabezuelas, a veces llamadas capítulos, la presencia de vilano (cáliz modificado en las flores de Asteraceae, generalmente constituido por elementos de apariencia seca, ya sean cerdas, escamas, aristas, etc.) y a los estambres sinantéreos (aquellos que tienen filamentos libres, anteras fusionadas con dehiscencia longitudinal introrsa y están dispuestas alrededor del estilo y el estigma), carácter diagnóstico de la familia (Funk et al., 2009; Villaseñor, 2018). Sin embargo, durante revisiones rutinarias de material herborizado es posible encontrar especímenes de Ranunculaceae y Apiaceae erróneamente identificados con algún nombre correspondiente a Asteraceae. Esta situación ocurre cuando las únicas estructuras reproductivas que contienen los ejemplares son frutos, ya que en las tres familias son secos. Sin embargo, en Asteraceae son cipselas porque derivan de un ovario ínfero unilocular (Marzinek et al., 2008), y aunque Apiaceae también tiene ovario ínfero, este es bicarpelar y el fruto un esquizocarpo, mientras que en Ranunculaceae el ovario es súpero, unilocular y los frutos son aquenios, bayas, drupas o folículos (Cronquist, 1981). Cuando los especímenes presentan flores resulta más difícil confundirlos, ya que Apiaceae y Ranunculaceae tienen anteras libres (Cronquist, 1981), mientras que en Asteraceae están fusionadas. Filogeneticamente, Asteraceae y Apiaceae están relacionadas, pues ambas forman parte del clado Campanulides; caso contrario a Ranunculaceae que está ubicada en las primeras ramas de las Eudicotiledóneas (APG IV, 2016).
El reconocimiento de subfamilias, tribus, géneros y especies de Asteraceae suele ser complejo, ya que además de caracteres macromorfológicos (p. ej., filotaxia, forma, conación y número de series del involucro o tipo de cabezuelas), es necesario observar y analizar microcaracteres (Villaseñor, 1987; Funk et al., 2009). Es decir, el conjunto de características que se cuantifican o describen en una estructura analizada al microcopio, ya sea estereoscópico, óptico o electrónico. Entre ellas se encuentran: sexualidad de las flores (femeninas, masculinas o bisexuales), orientación, forma e indumento de las ramas del estilo, variaciones en los apéndices apicales y basales de las anteras, forma, volumen, número de costillas, presencia o ausencia de indumento en las cipselas, número de series, coloración y elementos del vilano, entre otros (Karis, 1993b; Baldwin et al., 2002; Panero y Funk, 2002, 2008; Roque y Funk, 2013). En algunos casos la presencia de metabolitos secundarios, números cromosómicos, hábitat o características observables únicamente in vivo, también son de utilidad taxonómica para delimitar algunos grupos (Funk et al., 2009). Por ello, resulta importante familiarizarse con las distintas estructuras que conforman las cabezuelas, así como con la terminología empleada para nombrarlas.
Los objetivos de este trabajo son presentar una clave de identificación para las tribus de Asteraceae nativas e introducidas presentes en el país e ilustrar los caracteres diagnósticos de cada una.
Materiales y Métodos
Para recopilar las características distintivas de cada una de las 26 tribus de asteráceas presentes en México, incluyendo a las dos introducidas (Villaseñor, 2018), se realizó una revisión bibliográfica de trabajos relacionados con la clasificación de Asteraceae, cambios en la circunscripción tribal y tratamientos taxonómicos enfocados en grupos presentes en el país publicados principalmente en las dos últimas décadas (Panero y Funk, 2002, 2008; Baldwin et al., 2002; Funk et al., 2009; Roque y Funk, 2013; Redonda-Martínez, 2018; Panero, 2019; Redonda-Martínez, 2020; Susanna et al., 2020; Lichter-Marck y Baldwin, 2022) y la policlave para tribus del mundo (Bonifacino y Funk, 2018). También se realizó un análisis morfológico de material fresco, conservado en alcohol etílico al 70% (en espíritu) y herborizado, complementado con observaciones hechas durante expediciones de campo en el periodo 2010-2019. Con la finalidad de hacer más accesible la clave generada, se definieron de manera sencilla los caracteres empleados en el reconocimiento tribal, tomando como referencia la terminología del glosario ilustrado de Compositae (Roque et al., 2009), optando por incluir fotografías de aquellas estructuras o características que podrían resultar confusas para el lector no especializado en la familia.
Las fotografías de las cabezuelas ≥3 cm fueron obtenidas con una cámara digital (Olympus VR370, Tokio, Japón) o con un teléfono inteligente (Motorola G6 Play, Libertyville, EUA). Las imágenes de las cabezuelas ≤1 cm, las flores, las anteras, los estilos, el vilano y las cipselas se tomaron con un microscopio estereoscópico (Leica EZ4HD, Heerbrugg, Suiza) con cámara integrada. Cada foto se guardó en formato .tiff con una resolución de 600 dpi. En algunas fotografías se ajustaron los valores de brillo y color con el analizador de imágenes Leica Application Suite EZ v. 3.4.0 (Leica Microsystems, 2021). Las figuras que ilustran este trabajo se diseñaron con Adobe Photoshop CS5 Extended v. 12.0 (Knoll et al., 2010).
Finalmente, se actualizó la diversidad conocida de Asteraceae en México. Para ello, se consideraron como punto de partida las cifras y nombres registrados por Villaseñor (2016; 2018). A estos se sumaron los registros de especies recién incorporadas a la flora mexicana, ya sea por ser nuevas para la ciencia, no reportadas en los trabajos referidos anteriormente, o conocidas de otras regiones y documentadas en el país en los últimos cuatro años. En algunas tribus hubo reducción de números debido a cambios nomenclaturales (principalmente sinonimias a nivel de géneros y especies) o recircunscripción de algunos géneros, posteriores a la publicación de los trabajos antes mencionados; es decir, de junio de 2018 a septiembre de 2022.
Resultados y Discusión
Diversidad de Asteraceae en México
Considerando la propuesta más reciente de clasificación de Asteraceae (Susanna et al., 2020), en México se encuentran seis subfamilias que agrupan 26 tribus; dos de ellas, Calenduleae y Arctotideae, son introducidas. El mayor número de tribus (17) se encuentra en la subfamilia Asteroideae, seguida de Mutisioideae y Vernonioideae con tres, cada una; mientras que Carduoideae, Cichorioideae y Gochnatioideae están representadas por una sola tribu cada una (Cuadro 1). En cuanto al número de géneros y especies, Villaseñor (2018) determinó que en el país había 417 géneros y 3113 especies, de las cuales 3050 eran nativas y 1988 endémicas. Después de actualizar las cifras, en nuestro país existen 428 géneros, de los cuales 324 son nativos, 67 endémicos y 37 introducidos; el número de especies aumentó a 3127, de ellas 3065 son nativas, 1996 endémicas y 64 introducidas (Cuadro 1).
Cuadro 1: Riqueza de especies de Asteraceae con distribución en México. Los números en negritas corresponden a cifras actualizadas con base en cambios nomenclaturales, de circunscripción, especies y registros nuevos disponibles en el apéndice; los demás fueron retomados de Villaseñor (2018). GN=géneros nativos (sin considerar endémicos), GE=géneros endémicos, GI=géneros introducidos, TG=total de géneros; EN=especies nativas con distribución dentro y fuera de México, EE=especies endémicas, EI=especies introducidas, TE=total de especies.
| Tribus | Subfamilias | GN | GE | GI | TG | EN | EE | EI | TE |
|---|---|---|---|---|---|---|---|---|---|
| Anthemideae | Asteroideae | 3 | 0 | 8 | 11 | 11 | 3 | 16 | 30 |
| Arctotideae | Vernonioideae | 0 | 0 | 2 | 2 | 0 | 0 | 2 | 2 |
| Astereae | Asteroideae | 49 | 2 | 2 | 53 | 178 | 201 | 3 | 382 |
| Bahieae | Asteroideae | 13 | 1 | 0 | 14 | 25 | 23 | 0 | 48 |
| Calenduleae | Asteroideae | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 |
| Cardueae | Carduoideae | 2 | 0 | 6 | 8 | 18 | 28 | 11 | 57 |
| Chaenactideae | Asteroideae | 1 | 0 | 0 | 1 | 7 | 1 | 0 | 8 |
| Cichorieae | Cichorioideae | 18 | 1 | 11 | 30 | 49 | 19 | 17 | 85 |
| Coreopsideae | Asteroideae | 12 | 2 | 0 | 14 | 57 | 144 | 0 | 201 |
| Eupatorieae | Asteroideae | 44 | 8 | 0 | 52 | 186 | 437 | 0 | 623 |
| Gnaphalieae | Asteroideae | 13 | 1 | 1 | 15 | 46 | 23 | 4 | 73 |
| Gochnatieae | Gochnatioideae | 1 | 1 | 0 | 2 | 1 | 6 | 0 | 7 |
| Helenieae | Asteroideae | 10 | 1 | 0 | 11 | 29 | 14 | 0 | 43 |
| Heliantheae | Asteroideae | 51 | 23 | 1 | 75 | 185 | 460 | 1 | 644 |
| Inuleae | Asteroideae | 4 | 0 | 0 | 4 | 9 | 2 | 0 | 11 |
| Liabeae | Vernonioideae | 2 | 1 | 0 | 3 | 15 | 10 | 0 | 25 |
| Madieae | Asteroideae | 12 | 2 | 0 | 14 | 28 | 7 | 0 | 35 |
| Millerieae | Asteroideae | 22 | 5 | 0 | 27 | 45 | 137 | 0 | 175 |
| Mutisieae | Mutisioideae | 4 | 0 | 0 | 4 | 7 | 9 | 0 | 16 |
| Nassauvieae | Mutisioideae | 3 | 0 | 0 | 3 | 14 | 83 | 0 | 97 |
| Neurolaeneae | Asteroideae | 4 | 1 | 0 | 5 | 9 | 19 | 0 | 28 |
| Onoserideae | Mutisioideae | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 1 |
| Perityleae | Asteroideae | 7 | 1 | 0 | 8 | 9 | 44 | 0 | 53 |
| Senecioneae | Asteroideae | 16 | 4 | 3 | 23 | 54 | 167 | 6 | 227 |
| Tageteae | Asteroideae | 17 | 10 | 0 | 27 | 60 | 112 | 1 | 173 |
| Vernonieae | Vernonioideae | 15 | 3 | 2 | 20 | 26 | 47 | 2 | 75 |
| Total 26 | 6 | 324 | 67 | 37 | 428 | 1069 | 1996 | 64 | 3127 |
En términos generales hubo un incremento en la riqueza de Asteraceae con distribución en México con respecto a los datos previamente publicados (Villaseñor, 2016; 2018). Esto debido principalmente a la descripción de géneros nuevos en las tribus Gochnatieae (Panero, 2019) y Senecioneae (García-Mendoza et al., 2020), así como la incorporación de nuevos registros en Astereae (Hinojosa-Espinosa et al., 2021), Cichorieae (Pruski, 2018) y Vernonieae. En esta última tribu destaca Trichospira verticillata (L.) S.F. Blake, especie documentada a partir de un ejemplar colectado en Tabasco (F. Ventura A. 20566 (IEB)) hace casi cuarenta años, cuya presencia pasó desapercibida a la vista de los especialistas, a pesar de que el tipo corresponde a un espécimen proveniente de Veracruz según consta en Hortus Cliffortianus (399 (1737)) (Blake, 1915) (Apéndice). La resurrección de algunos géneros en Eupatorieae (Pruski, 2018) y Perityleae (Lichter-Marck y Baldwin, 2022), además del reconocimiento válido de taxones que habían sido sinonimizados en Mutisieae (Redonda-Martínez, 2018).
Las tribus Helenieae, Heliantheae y Liabeae registraron una ligera disminución de géneros y especies debido a errores de circunscripción, aceptación de sinónimos como nombres válidos o viceversa (Villaseñor, 2016; 2018), los cuales fueron corregidos en trabajos recientes (Pruski, 2018; Lichter-Marck y Baldwin, 2022). En Bahieae también hubo actualizaciones nomenclaturales (Baldwin y Wood, 2016); sin embargo, las cifras permanecieron igual (Cuadro 1, Apéndice).
En Eupatorieae, Gnaphaliae e Inuleae también se registraron ajustes nomenclaturales que derivaron en la reducción a sinonimia o correcta circunscripción de algunas especies (Redonda-Martínez y Martínez Salas, 2019; Redonda-Martínez, 2020; Freire et al., 2022). Sin embargo, debido a la descripción de nuevos taxones durante los últimos cuatro años hubo un incremento a nivel familia. Con respecto a las tribus, la riqueza de especies aumentó en Eupatorieae (8 spp.), seguida de Coreopsideae, Perityleae y Senecioneae con dos taxones cada una, además de Astereae, Heliantheae y Millerieae, cada tribu con una especie. En el caso de Mutisieae, el número de especies se incrementó como resultado de estudios taxonómicos que permitieron reconocer la validez de taxones reducidos erróneamente a sinónimos (Redonda-Martínez, 2018) o que solían confundirse con otros (Redonda-Martínez, 2018; 2022) (Cuadro 1, Apéndice).
Caracteres de importancia taxonómica
Para reconocer y diferenciar cada una de las 26 tribus de Asteraceae presentes en México (Fig. 1), es necesario combinar macro y microcaracteres. Los más importantes son el tipo de cabezuelas, flores que las conforman, sexualidad, simetría y arreglo de los lóbulos de las corolas, conación y número de series del involucro, presencia o ausencia de calículo, forma del ápice y base de las anteras, presencia o ausencia y contorno del collar, orientación y forma de las ramas del estilo, así como disposición del indumento, presencia o ausencia de fitomelanina en las cipselas, presencia o ausencia de vilano, simetría, número de series y elementos que lo conforman.

Figura 1: Especies representativas de cada una de las 26 tribus de Asteraceae presentes en México; los asteriscos indican las tribus introducidas. A. Anthemideae, Achillea millefolium L.; B. Arctotideae*, Gazania Gaertn.; C. Astereae, Heterotheca inuloides Cass.; D. Bahieae, Schkuhria pinnata (Lam.) Kuntze ex Thell.; E. Calenduleae*, Calendula officinalis L.; F. Cardueae, Cirsium rhaphilepis (Hemsl.) Petr.; G. Chaenactideae, Chaenactis lacera Greene; H. Cichorieae, Pinaropappus roseus (Less.) Less.; I. Coreopsideae, Dahlia imperialis Roezl ex Ortgies; J. Eupatorieae, Stevia microchaeta Sch. Bip.; K. Gnaphalieae, Pseudognaphalium chartaceum (Greenm.) Anderb.; L. Gochnatieae, Nahuatlea obtusata (S.F. Blake) V.A. Funk; M. Helenieae, Helenium mexicanum Kunth; N. Heliantheae, Zinnia peruviana (L.) L.; O. Inuleae, Pluchea salicifolia (Mill.) S.F. Blake var. salicifolia; P. Liabeae, Sinclairia andrieuxii (DC.) H. Rob. & Brettell; Q. Madieae, Madia exigua (Sm.) A. Gray; R. Millerieae, Galinsoga parviflora Cav.; S. Mutisieae, Chaptalia pringlei Greene; T. Nassauvieae, Trixis grandibracteata C.E. Anderson; U. Neurolaeneae, Calea ternifolia Kunth; V. Onoserideae, Onoseris onoseroides (Kunth) B.L. Rob.; W. Perityleae, Perityle emoryi Torr.; X. Senecioneae, Pittocaulon praecox (Cav.) H. Rob. & Brettell; Y. Tageteae, Dyssodia decipiens (Bartl.) M.C. Johnst.; Z. Vernonieae, Lepidaploa canescens (Kunth) H. Rob. Escalas D=1 mm, Q y R=2 mm. Fotos: Rosario Redonda-Martínez, excepto T, Fernando Araujo Mondragón.
Cabezuelas y flores
La inflorescencia básica de las asteráceas son las cabezuelas. Agrupan una, decenas o cientos de flores. Cuando todas son iguales, la cabezuela se denomina homógama (Fig. 2A, C, E, G). En cambio, si presenta dos o tres tipos de flores diferentes es heterógama (Fig. 2I, K, M, O), ya que se distinguen fácilmente las flores periféricas de las centrales (Cuadros 2, 3).

Figura 2: Cabezuelas y flores de Asteraceae. A-B. Vernonia alamanii DC., A. cabezuela homógama discoide; B. flor bisexual, corola infundibuliforme; C-D. Baccharis heterophylla Kunth; C. cabezuela homógama discoide con flores femeninas; D. flor femenina, corola cilíndrico filiforme; E-F. Baccharis salicifolia (Ruiz & Pav.) Pers.; E. cabezuela homógama discoide con flores masculinas; F. flor masculina, corola tubular angosta; G. Sonchus oleraceus L., cabezuela homógama ligulada; H. Taraxacum officinale F.H. Wigg., flor bisexual, corola ligulada, 5 dentada en el ápice; I-J. Helianthus annuus L.; I. cabezuela heterógama radiada; J. flor periférica neutra, corona radiada (fprn) y flores centrales bisexuales, corolas cilíndrico tubulares (fc), la primera mostrando las ramas del estilo y la segunda las anteras; K. Chaptalia pringlei Greene, cabezuela heterógama radiada, flores periféricas con corolas radiadas y flores centrales con corolas bilabiadas; L. Chaptalia hidalgoensis L. Cabrera & G.L. Nesom, flor periférica externa femenina, corola radiada (fper), flor periférica interna femenina, corola filiforme (fpif), flor central bisexual, corola bilabiada (fcb) (por motivos de espacio no se muestran androceo ni gineceo); M-N. Gerbera jamesonii Adlam; M. cabezuela heterógama radiado-bilabiada; N. flor periférica externa femenina, corola radiado-bilabiada (fperb), flor periférica interna femenina, corola radiado-bilabiada (fpirb), flor central bisexual, corola bilabiada (fcb); O-P. Conyza bonariensis (L.) Cronquist; O. cabezuela heterógama disciforme; P. flor periférica femenina, corola filiforme (fpf) y flor central bisexual, corola tubular (fct). Fotos: Rosario Redonda-Martínez.
Cuadro 2: Características de las cabezuelas y las flores periféricas en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas, NA=No Aplica.
| Tribus | Cabezuelas | Flores periféricas | |||
|---|---|---|---|---|---|
| Tipo | Número de series | Sexualidad | Forma de las corolas | Coloración predominante | |
| Anthemideae | heterógamas radiadas | uniseriadas | femeninas | radiadas, 1 lobuladas, a veces 3-dentadas en el ápice | blancas o amarillas |
| Arctotideae* | heterógamas radiadas | uniseriadas | femeninas | radiadas, 1 o 2-dentadas en el ápice | tricoloras, rara vez blanquecinas |
| Astereae | heterógamas radiadas, a veces disciformes | uni o multiseriadas | femeninas | radiadas, 1 lobuladas en el ápice; a veces filiformes | amarillas o blancas, a veces rosadas, moradas, azules o verdosas |
| Bahieae | heterógamas radiadas, a veces homógamas discoides | uniseriadas | femeninas | radiadas, 1-3-lobuladas en el ápice | amarillas, a veces blancas |
| Calendueleae* | heterógamas radiadas | multiseriadas | femeninas | radiadas, 3-dentados en el ápice | anaranjadas, a veces amarillas |
| Cardueae | homógamas discoides | NA | NA | NA | NA |
| Chaenactideae | homógamas discoides | NA | NA | NA | NA |
| Cichorieae | homogámas liguladas | NA | NA | NA | NA |
| Coreopsideae | heterógamas radiadas | uniseriadas | femeninas o neutras | radiadas, 1 o a veces 3-4 lobuladas o dentadas en el ápice | amarillas, blancas, rojizas, rosadas o moradas |
| Eupatorieae | homógamas discoides | NA | NA | NA | NA |
| Gnaphalieae | heterógamas disciformes | multiseriadas | femeninas | filiformes o tubular angostas | verdosas, blanquecinas o amarillentas |
| Gochnatieae | homógamas discoides | NA | NA | NA | NA |
| Helenieae | heterógamas radiadas | uniseriadas | femeninas | radiadas, profundamente 3 lobuladas en el ápice | amarillas, a veces anaranjadas o rojizas en plantas cultivadas |
| Heliantheae | heterógamas radiadas, rara vez disciformes u homógamas discoides | uniseriadas | neutras, a veces femeninas | radiadas, 1-3 lobuladas en el ápice; a veces filiformes, reducidas o ausentes | amarillas, a veces blancas, anaranjadas, rojizas, rosadas o moradas |
| Inuleae | heterógamas disciformes | multiseriadas | femeninas | filiformes | rosadas o moradas, rara vez blanquecinas |
| Liabeae | heterógamas radiadas, a veces homógamas discoides | uniseriadas | femeninas | radiadas, 3-dentadas en el ápice; a veces reducidas o filiformes | amarillas o anaranjadas |
| Madieae | heterógamas radiadas | uniseriadas | femeninas | radiadas, 1-3 lobuladas en el ápice | amarillas |
| Millerieae | heterógamas radiadas | uniseriadas, a veces biseriadas | femeninas, rara vez neutras | radiadas, 2 dentadas o 3 lobuladas en el ápice | amarillas, a veces blancas |
| Mutisieae | heterógamas radiadas | biseriadas, multiseriadas en plantas cultivadas (Gerbera L.) | femeninas | radiadas, 1 lobuladas en el ápice, o radiado-bilabiadas (3+2), a veces reducidas o filiformes | blanquecinas o rosadas, colores llamativos solo en plantas cultivadas (Gerbera) |
| Nassauvieae | homógamas discoides, a veces heterógamas radiadas | uniseriadas cuando presentes | femeninas cuando presentes | radiado-bilabiadas (3+2) cuando presentes | rosadas, blanquecinas o amarillas cuando presentes |
| Neurolaeneae | heterógamas radiadas, a veces homógamas discoides | uniseriadas | femeninas | radiadas, 1 o 3 lobuladas en el ápice; a veces tubulares, reducidas o ausentes | amarillas o blancas |
| Onoserideae | homógamas discoides | NA | NA | NA | NA |
| Perityleae | heterógamas radiadas | uniseriadas | femeninas | radiadas, 3 lobuladas en el ápice | blancas o amarillas |
| Senecioneae | heterógamas radiadas, a veces disciformes; en ocasiones homógamas discoides | uniseriadas | femeninas | radiadas, 1 lobuladas o 3 dentadas en el ápice; a veces filiformes, reducidas o ausentes | amarillas, a veces rosadas, blanquecinas, rojizas o anaranjadas |
| Tageteae | heterógamas radiadas, a veces homógamas discoides | uniseriadas | femeninas | radiadas, 1 lobuladas en el ápice; a veces reducidas o ausentes, con glándulas oleíferas o cavidades secretoras | amarillas, a veces blancas, rojizas o anaranjadas |
| Vernonieae | homógamas discoides | NA | NA | NA | NA |
Cuadro 3: Características de las flores centrales en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas.
| Tribus | Sexualidad | Forma de las corolas | Lóbulos | Coloración predominante |
|---|---|---|---|---|
| Anthemideae | bisexuales | tubulares o campanuladas, pentámeras | libres, divididos ≤¼ de la longitud de la corola, erectos, a veces apicalmente reflexos | amarillas, a veces blanquecinas o verdosas |
| Arctotideae* | bisexuales | tubular infundibuliformes, pentámeras | libres, divididos ≤¼ de la longitud de la corola, erectos | amarillas |
| Astereae | bisexuales, a veces masculinas o femeninas | tubulares angostas, a veces filiformes o tubular infundibuliformes, pentámeras | libres, divididos ≤1/8 de la longitud de la corola, erectos; ausentes en flores filiformes | amarillas, blanquecinas o verdosas |
| Bahieae | bisexuales | tubulares o campanuladas, pentámeras | libres, divididos ≤1/3 de la longitud de la corola, erectos; generalmente con glándulas | amarillas |
| Calenduleae* | funcionalmente masculinas | tubular infundibuliformes, pentámeras | libres, divididos ≤1/4 de la longitud de la corola, erectos | anaranjadas o amarillas |
| Cardueae | bisexuales | infundibuliformes, pentámeras; a veces filiformes | libres, profundamente divididos ≥1/2 de la longitud de la corola, erectos | moradas o rosadas, a veces azules, anaranjadas o blanquecinas |
| Chaenactideae | bisexuales | tubular infundibuliformes, pentámeras | libres, divididos ≤1/4 de la longitud de la corola, erectos, glandulares | blancas, a veces rosadas o amarillas |
| Cichorieae | bisexuales | liguladas, 5-dentadas en el ápice | fusionados (5+0) | amarillas, a veces rosadas o azules |
| Coreopsideae | bisexuales | tubular infundibuliformes o campanuladas, pentámeras | libres, divididos ≤1/4 de la longitud de la corola, erectos, a veces glandulares | amarillas |
| Eupatorieae | bisexuales | tubiformes, pentámeras | libres, divididos ≤1/8 - 1/10 de la longitud de la corola, erectos, a veces recurvados, glandulares o pilosos | blancas, rosadas, moradas o azules, a veces rojizas o verdosas |
| Gnaphalieae | bisexuales, a veces funcionalmente masculinas | tubulares, pentámeras | libres, divididos ≤1/4 de la longitud de la corola, erectos, a veces glandulares, pilosos o glabros | verdosas, blanquecinas o amarillentas, a veces rojizas en el ápice |
| Gochnatieae | bisexuales | infundibuliformes, pentámeras | libres, profundamente divididos ≥1/2 de la longitud de la corola, reflexos y enrollados en la antesis | blanquecinas |
| Helenieae | bisexuales | campanuladas o tubulares, pentámeras, glandulares | libres, ligeramente divididos ≤1/10 de la longitud de la corola, erectos, glandulares, pilosos o papilosos | amarillas, ennegrecidas en el ápice |
| Heliantheae | bisexuales | campanuladas o tubulares, pentámeras | libres, inconspicuamente divididos ≥1/5 - ≤1/10 de la longitud de la corola, erectos, glabros, papilosos, pilosos o glandulares | amarillas, a veces blanquecinas, anaranjadas o rosadas |
| Inuleae | bisexuales o funcionalmente masculinas | tubular infundibuliformes, pentámeras | libres, divididos ≥1/4 de la longitud de la corola, erectos, a veces glandulares | rosadas o moradas, rara vez blanquecinas |
| Liabeae | bisexuales | infundibuliformes, pentámeras | libres, divididos ≥1/3 de la longitud de la corola, reflexos, a veces enrollados en la antesis | amarillas o anaranjadas |
| Madieae | bisexuales | tubulares, pentámeras | libres, divididos ≥1/4 de la longitud de la corola, erectos | amarillas |
| Millerieae | funcionalmente masculinas, a veces bisexuales | tubulares, campanuladas, a veces tubular infundibuliformes, pentámeras | libres, divididos ≤1/4 de la longitud de la corola; erectos o recurvados, papilosos, a veces glandulares | amarillas |
| Mutisieae | bisexuales o funcionalmente masculinas | bilabiadas, a veces tubular infundibuliformes, pentámeras | fusionados, 3+2 en corolas bilabiadas, recurvados; ligeramente divididos ≤¼ de la longitud en corolas actinomorfas; erectos o recurvados, glabros | blanquecinas o rosadas, colores llamativos solo en plantas cultivadas (Gerbera L.) |
| Nassauvieae | bisexuales | bilabiadas, a veces tubular infundibuliformes, pentámeras | fusionados, 3+2 en corolas bilabiadas, recurvados; ligeramente divididos ≤¼ de la longitud en corolas actinomorfas; erectos o recurvados, glabros | rosadas, blanquecinas o amarillentas |
| Neurolaeneae | bisexuales | infundibuliformes o campanuladas, pentámeras | libres, divididos ≥1/4 o ≤1/3 de la longitud de la corola; erectos o recurvados, glabros, a veces glandulares | amarillas o blancas |
| Onoserideae | bisexuales | bilabiadas | fusionados 3+2, recurvados | rojizas o rosadas |
| Perityleae | bisexuales | tubular campanuladas, tetrámeras; a veces pentámeras | libres, divididos ≥1/4 de la longitud de la corola; erectos o recurvados; glabros, a veces glandulares | amarillas, a veces blancas |
| Senecioneae | bisexuales | tubulares o campanuladas, a veces cilíndricas o filiformes; pentámeras | libres, divididos ≥1/3 de la longitud de la corola; erectos o recurvados; glabros, a veces glandulares o pilosos | amarillas, a veces blanquecinas, rojizas o anaranjadas |
| Tageteae | bisexuales | tubulares, a veces cilíndricas; pentámeras | libres, divididos ≥1/3 de la longitud de la corola; erectos, a veces recurvados; con glándulas oleíferas o cavidades secretoras | amarillas, a veces rojizas moradas o verdosas |
| Vernonieae | bisexuales | infundibuliformes, pentámeras | libres, divididos ≥1/3 de la longitud de la corola; erectos o reflexos; glandulares o papilosos, a veces pilosos o glabros | moradas, a veces rosadas o blanquecinas |
Existen dos clases de cabezuelas homógamas, las cuales se diferencian por el tipo de flores que las conforman. En la primera están las discoides (Fig. 2A, C, E), que presentan flores bisexuales con corolas tubulares, infundibuliformes o en ocasiones campanuladas, pentámeras con lóbulos cortos o profundamente divididos, siendo las más comunes en la familia (Fig. 2A, B). A veces, pueden tener flores unisexuales, femeninas (Fig. 2C, D) o masculinas (Fig. 2E, F), con corolas tubulares o filiformes (Fig. 2D, F). Aunque no es muy común en especies mexicanas, algunos grupos presentan flores bisexuales con corolas bilabiadas, las cuales tienen tres lóbulos fusionados formando un labio superior y dos conformando el labio inferior. Son comunes en las tribus Nassauvieae (Fig. 1T) y Onoserideae (Fig. 1VV), y también llegan a presentarse en algunas Mutisieae (Fig. 1S), aunque esta tribu tiene cabezuelas heterógamas. La segunda clase de cabezuelas homógamas tiene flores bisexuales liguladas, cuyas corolas son aplanadas, con cinco dientes en el ápice y se encuentran exclusivamente en la tribu Cichorieae (Fig. 2G, H).
Las cabezuelas heterógamas también se clasifican en dos categorías, considerando la forma de la corola de las flores periféricas: pueden ser radiadas (Fig. 2I, K, M) o disciformes (Fig. 2O). Las primeras tienen corolas aplanadas cuyos lóbulos generalmente están fusionados aparentando ser uno solo, aunque a veces es posible diferenciar tres, como en algunas Anthemideae (Fig. 1A), Helenieae (Fig. 1M) o Millerieae (Fig. 1R) (Cuadro 2). Las flores radiadas se consideran neutras cuando carecen de gineceo funcional o es reminiscente (p. ej. en Heliantheae, Fig. 2J), pero si el gineceo está bien desarrollado y es funcional son femeninas (p. ej. en Mutisieae, Fig. 2L); por lo tanto, producen cipselas después de la fecundación, mientras que las neutras no.
En ocasiones las flores radiadas pueden tener corolas sub-bilabiadas, en las cuales el limbo está formado por tres lóbulos fusionados de igual longitud y dos más cortos, libres o fusionados, formando un labio generalmente inconspicuo; desarrollan gineceo funcional y por tanto son femeninas. Debido a las modificaciones y el arreglo de los lóbulos, se han denominado radiado-bilabiadas (Redonda-Martínez, 2022); se encuentran en algunas Mutisieae (Fig. 2M, N) y a veces presentan estaminodios (p. ej. Gerbera L. (Redonda-Martínez, 2018)). En tratamientos taxonómicos recientes (Rzedowski y Calderón de Rzedowski, 2008; Redonda-Martínez, 2018, 2019), las flores con corolas radiadas o radiado-bilabiadas han sido consideradas o citadas como “liguladas” únicamente porque tienen corolas aplanadas. Incluso McVaugh (1984) usó los términos radiada y ligulada de manera indistinta, a pesar de que reconoció las diferencias existentes entre ellas.
Por último, las cabezuelas disciformes (Fig. 2O) se diferencian de las anteriores porque en la periferia tienen flores con corolas filiformes (Fig. 2P), las cuales son similares a un tubo que cubre el estilo, ya que rara vez tienen lóbulos (Cuadro 2). Debido a su apariencia, es común que las personas no familiarizadas con la morfología de Asteraceae confundan este tipo de cabezuelas con las homógamas discoides. Sin embargo, también sucede con los especialistas. Por ejemplo, McVaugh (1984) las integró en la misma copla de una clave, a pesar de que las flores filiformes son femeninas y carecen de androceo, como ocurre en Gnaphalieae (Fig. 1K), Inuleae (Fig. 1Q) y algunos miembros de Astereae (Fig. 2O, P) o Mutisieae (Fig. 2L). Sin importar el tipo de flores periféricas que contengan las cabezuelas heterógamas, ya sean radiadas o disciformes, las flores centrales o discoides (ya sea que tengan corolas tubulares, infundibuliformes, campanuladas o bilabiadas), son bisexuales (Cuadro 3). Aunque a veces son funcionalmente masculinas; es decir, que desarrollan gineceo, pero este no es funcional; por tanto, solo las flores periféricas producirán cipselas (p. ej. Calenduleae y algunas Millerieae). Aun cuando Roque et al. (2009) consideraron que Baccharis L. tenía cabezuelas disciformes, aunque contienen únicamente flores femeninas o masculinas, en el presente trabajo se denominan homógamas; tal y como sugirieron Heiden y Bonifacino (2021).
Receptáculo y páleas
El receptáculo es el área basal de la cabezuela donde están insertos los verticilos que la integran; es decir, los filarios, las páleas y las flores (Roque et al., 2009). Su forma no es fácil de distinguir a menos que se haga un corte longitudinal de la cabezuela, o bien cuando las cipselas se han desprendido. Lo más común es que sea plano como en Helianthus annuus L. y Verbesina montanoifolia B.L. Rob. & Greenm. (Fig. 3A, B). Sin embargo, también puede ser cónico (p. ej. Millerieae, Melampodium divaricatum (Rich.) DC., Fig. 3C), convexo (Astereae, Erigeron L., Fig. 3D) o globoso (Helenieae, Helenium L. Fig. 1M). No obstante, las características más utilizadas para distinguir tribus, géneros o especies son la presencia o ausencia de páleas, alveolos e indumento. Si el receptáculo tiene páleas se denomina paleáceo (p. ej. Heliantheae, Fig. 3A, B, I, J o Coreopsideae, Fig. 3H) y epaleáceo cuando carece de ellas (p. ej. Astereae, Fig. 3D, F o Vernonieae Fig. 3E). Las cipselas que se desprenden del receptáculo dejan cicatrices semejantes a las celdas de un panal, estas se denominan alveolos y el receptáculo que las presenta se conoce como alveolado (p. ej. Vernonieae, Vernonia alamanii DC., Fig. 3E), escamoso si tiene escamas (p. ej. Astereae, Baccharis salicifolia (Ruiz & Pav.) Pers., Fig. 3F), piloso cuando presenta tricomas o cerdas reducidas; con cerdas al tener estructuras similares a las cerdas del vilano (p. ej. Cardueae, Cirsium velatum (S. Watson) Petr., Fig. 3G) y glabro si carece de indumento (Fig. 3D, E) (Cuadro 4).

Figura 3: Receptáculo en cabezuelas de Asteraceae. A. Heliantheae, Helianthus annuus L., receptáculo plano con páleas moradas en el ápice; B. Heliantheae, Verbesina montanoifolia B.L. Rob. & Greenm., receptáculo plano, paleáceo; C. Millerieae, Melampodium divaricatum (Rich.) DC., receptáculo cónico con páleas deciduas como unidad; D. Astereae, Erigeron L., receptáculo convexo, epaleáceo; E. Vernonieae, Vernonia alamanii DC., receptáculo alveolado, convexo, epaleáceo; F. Astereae, Baccharis salicifolia (Ruiz & Pav.) Pers., receptáculo plano, escamoso; G. Cardueae, Cirsium velatum (S. Watson) Petr., receptáculo plano con cerdas; H. Coreopsideae, Heterosperma pinnatum Cav., páleas laminares con estrías paralelas pardo-anaranjadas (*); I. Heliantheae, Tithonia rotundifolia (Mill.) S.F. Blake, cabezuela con páleas esclerificadas en el ápice (+); J. Heliantheae, Montanoa bipinnatifida (Kunth) K. Koch, cabezuelas con páleas conduplicadas acrescentes (pca); a=alveolo, c=cipselas, ce=cerdas, e=escamas, p=páleas, r=receptáculo. Fotos: Rosario Redonda-Martínez.
Cuadro 4: Características del receptáculo en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas.
| Tribus | Forma | Superficie | Páleas |
|---|---|---|---|
| Anthemideae | plano, hemisférico o cónico | glabro o piloso | laminares, a veces ausentes |
| Arctotideae* | plano | alveolado | ausentes |
| Astereae | plano o convexo | a veces alveolado, escamoso o glabro | ausentes |
| Bahieae | plano o ligeramente convexo | glabro | ausentes |
| Calenduleae* | plano | glabro | ausentes |
| Cardueae | plano | alveolado, con cerdas | ausentes |
| Chaenactideae | plano | glabro | ausentes |
| Cichorieae | plano | alveolado, a veces piloso o escamoso | ausentes |
| Coreopsideae | plano o cónico | glabro | laminares, con estrías paralelas pardo-anaranjadas, deciduas o persistentes |
| Eupatorieae | cónico, convexo o plano | alveolado, glabro o piloso | laminares, presentes en Ageratum L. |
| Gnaphalieae | plano o convexo, a veces cónico | piloso o con escamas | ausentes |
| Gochnatieae | plano | alveolado, glabro | ausentes |
| Helenieae | globoso o plano | alveolado, glabro | ausentes |
| Heliantheae | plano o convexo, a veces cónico o columnar | a veces alveolado, glabro | conduplicadas, a veces filiformes, persistentes, en ocasiones acrescentes, esclerificadas o coloridas en el ápice, generalmente envolviendo completamente a los ovarios o las cipselas, a veces formando complejos con los filarios y las cipselas |
| Inuleae | plano o convexo | glabro | ausentes |
| Liabeae | plano | alveolado, a veces piloso | ausentes |
| Madieae | plano | glabro | laminares, escariosas; únicamente en las flores periféricas |
| Millerieae | plano, convexo o cónico | liso o alveolado, glabro, a veces piloso | laminares o naviculares, a veces deciduas como unidad o formando un complejo con las cipselas y los filarios |
| Mutisieae | convexo | alveolado | ausentes |
| Nassauvieae | plano | alveolado, glabro, a veces piloso o setoso | laminares, presentes en Jungia L. f. |
| Neurolaeneae | plano o convexo | alveolado, glabro | conduplicadas, rara vez ausentes |
| Onoserideae | plano | alveolado | ausentes |
| Perityleae | plano o convexo | liso o alveolado | ausentes |
| Senecioneae | plano, convexo o cónico | alveolado, fimbriado, denticulado o piloso | ausentes |
| Tageteae | plano o convexo | alveolado o liso | ausentes |
| Vernonieae | plano o convexo | alveolado | laminares, presentes en Bolanosa A. Gray |
Las páleas son brácteas que están dispuestas sobre el receptáculo y rodean parcial o totalmente a las flores o a las cipselas (Roque et al., 2009). Son comunes en Anthemideae, Coreopsideae, Heliantheae, Madieae, Millerieae, Neurolaeneae y pueden encontrarse esporádicamente en algunos géneros de Eupatorieae, Nassauvieae y Vernonieae. Generalmente son laminares, aunque también hay conduplicadas y naviculares (Cuadro 4). En ocasiones son coloridas, dándole tonos oscuros a las cabezuelas (p. ej. H. annuus, Fig. 3A) o acentuando el color de las flores (Fig. 3B, I). A veces también presentan indumento similar al de los filarios (Fig. 3B) u otras ornamentaciones que pueden ser de importancia taxonómica. Por ejemplo, en Coreopsideae tienen estrías paralelas pardo-anaranjadas (Fig. 3H), siendo este un carácter diagnóstico para reconocer a sus miembros. En Heliantheae son conduplicadas; a veces tienen el ápice esclerificado (p. ej. Tithonia Desf. ex Juss., Fig. 3I) o son acrescentes (p. ej. Montanoa Cerv., Fig. 3J). Generalmente persisten en la cabezuela, aunque en varias Coreopsideae son deciduas e incluso caen antes de las cipselas (p. ej. Bidens L. o Cosmos Cav.) (Cuadro 4).
Involucro, calículo y filarios
El involucro es el conjunto de brácteas, llamadas filarios o brácteas involucrales, que rodean las flores en una cabezuela (Roque et al., 2009). Dependiendo del número de series que lo integren se reconocen tres tipos: uniseriado (p. ej. Senecioneae, Fig. 4A), biseriado (p. ej. Coreopsideae, Fig. 4B) y multiseriado (p. ej. Liabeae, Fig. 4C). Cuando es uniseriado, todos los filarios tienen la misma longitud, apariencia, forma e indumento (Fig. 4A, I). Si presentan dos o más series (Fig. 4B, C), las brácteas involucrales suelen diferir en longitud, forma, coloración, apariencia o indumento. El ejemplo más claro se observa en Coreopsideae, tribu en la que los filarios de la serie externa suelen ser crasos, verdes, fusionados en la base y con menor o mayor longitud en comparación con los internos, siendo este un carácter diagnóstico para reconocer a esta tribu (Fig. 4B) (Cuadro 5).

Figura 4: Involucro y filarios de Asteraceae. A. Senecioneae, Pseudogynoxys haenkei (DC.) Cabrera, involucro campanulado, uniseriado con calículo (*); B. Coreopsideae, Cosmos caudatus Kunth, involucro hemisférico, biseriado, la serie externa con filarios verdes, crasos y fusionados en la base, la serie interna con filarios rojizos; C. Eupatorieae, Chromolaena odorata (L.) R.M. King & H. Rob., involucro cilíndrico, multiseriado; D. Gnaphalieae, Gamochaeta americana (Mill.) Wedd., filario papiráceo con estereoma (e); E. Millerieae, Melampodium divaricatum (Rich.) DC., filario herbáceo; F. Calenduleae, Calendula officinalis L., filario con margen escarioso; G. Anthemideae, Achillea millefolium L., filario escarioso, con glándulas sésiles (+); H. Perityleae, Perityle emoryi Torr., filarios naviculares; I. Tageteae, Adenophyllum glandulosum (Cav.) Strother, involucro campanulado con filarios fusionados en toda su extensión, calículo (*) presente y pústulas (~) secretoras de aroma anisado. Fotos: Rosario Redonda-Martínez.
Cuadro 5: Características del involucro y los filarios en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas.
| Tribus | Involucro | Filarios | |||||
|---|---|---|---|---|---|---|---|
| Número de series | Forma | Calículo | Fusión | Disposición | Forma | Textura y modificaciones | |
| Anthemideae | multiseriado | campanulado | ausente | libres | imbricados, graduados | laminares | escariosos, con glándulas sésiles |
| Arctotideae* | multiseriado | campanulado a hemisférico | ausente | libres | imbricados, graduados | laminares | membranáceos |
| Astereae | multiseriado | cilíndrico, turbinado o campanulado | ausente | libres | imbricados, graduados o subiguales | laminares | membranáceos, a veces herbáceos, escariosos en el margen |
| Bahieae | multiseriado | cilíndrico o turbinado, a veces hemisférico | ausente | libres | imbricados, subiguales | laminares | membranáceos |
| Calenduleae* | multiseriado | campanulado o hemisférico | ausente | libres | imbricados, subiguales | laminares | herbáceos con margen escarioso |
| Cardueae | multiseriado | campanulado, a veces urceolado | ausente | libres | imbricados, graduados | laminares | membranáceos, con el ápice modificado en una espina |
| Chaenactideae | uni o biseriado | campanulado a hemisférico | ausente | libres | imbricados, subiguales | laminares | membranáceos |
| Cichorieae | uni o multiseriado | cilíndrico o campanulado | presente en algunos géneros | libres | imbricados, subiguales o graduados | laminares | herbáceos o membranáceos, con látex; frecuentemente con el ápice de un color distinto al de la lámina |
| Coreopsideae | biseriado | cilíndrico, campanulado o hemisférico | ausente | los externos fusionados en la base, los internos libres | graduados | laminares | los externos suculentos, los internos membranáceos o herbáceos |
| Eupatorieae | multiseriado, a veces uni o biseriado | cilíndrico, turbinado o campanulado | ausente | libres | imbricados, graduados o subiguales | laminares, a veces ligeramente cóncavos | membranáceos |
| Gnaphalieae | multiseriado | campanulado, cilíndrico o turbinado, a veces urceolado | ausente | libres | imbricados, graduados | laminares o ligeramente cóncavos | papiráceos generalmente con estereoma |
| Gochnatieae | multiseriado | cilíndrico, turbinado o campanulado | ausente | libres | imbricados, graduados | laminares | membranáceos |
| Helenieae | biseriado, a veces multiseriado | campanulado a hemisférico | ausente | libres | imbricados, subiguales | laminares | herbáceos |
| Heliantheae | uniseriado o multiseriado | campanulado a hemisférico | ausente | libres | imbricados, graduados o subiguales | laminares | membranáceos o herbáceos |
| Inuleae | multiseriado | cilíndrico, campanulado o hemisférico, a veces turbinado | ausente | libres | imbricados, graduados o subiguales | laminares | membranáceos |
| Liabeae | multiseriado | campanulado, a veces turbinado | ausente | libres | imbricados, graduados | laminares | membranáceos, con látex |
| Madieae | uni o biseriado, a veces multiseriado | cilíndrico o campanulado, a veces urceolado | ausente | libres | subiguales | laminares | herbáceos o membranáceos, con glándulas estipitadas |
| Millerieae | multiseriado, a veces uni o biseriado | obcónico, campanulado o hemisférico, a veces cupuliforme | ausente | libres | subiguales, imbricados, a veces graduados | laminares | herbáceos, a veces cartáceos o escariosos |
| Mutisieae | multiseriado | campanulado, turbinado o hemisférico | ausente | libres | imbricados, graduados | laminares | membranáceos |
| Nassauvieae | multiseriado | cilíndrico a campanulado | ausente | libres | imbricados, graduados | laminares | herbáceos o membranáceos |
| Neurolaeneae | multiseriado, a veces uniseriado | cilíndrico, campanulado o hemisférico | ausente | libres | subiguales o graduados, imbricados | laminares | membranáceos, escariosos en el margen |
| Onoserideae | multiseriado | cilíndrico o turbinado | ausente | libres | imbricados, graduados | laminares | membranáceos |
| Perityleae | uni o biseriado | cilíndrico a hemisférico | ausente | libres | subiguales, imbricados | naviculares | membranáceos |
| Senecioneae | uniseriado | cilíndrico, turbinado, campanulado o hemisférico | presente en algunos géneros | libres | iguales, imbricados | laminares o ligeramente cóncavos | membranáceos, a veces con resina |
| Tageteae | uniseriado | turbinado, cilíndrico, campanulado o hemisférico | presente en algunos géneros | fusionados en toda su extensión, a veces de la parte media hacia la base | iguales | laminares | membranáceos o herbáceos, con glándulas oleíferas, pústulas o cavidades secretoras de aroma anisado |
| Vernonieae | multiseriado | Campanulado o hemisférico, a veces cilíndrico o turbinado | reducido en algunos géneros | libres | imbricados, graduados | laminares | membranáceos |
En un involucro multiseriado (Fig. 4C), por lo general las series externas son comparativamente más cortas que las internas (Fig. 4C). Una de las características empleadas para diferenciar tribus relacionadas filogenéticamente es la consistencia de los filarios. Por ejemplo, en Gnaphalieae suelen ser papiráceos y generalmente tienen un esteroma (estructura herbácea o cartilaginosa que se diferencia del margen y ápice escarioso en los filarios de las Gnaphalieae, pudiendo dividirlos o no (Roque et al., 2009)) en la base (Fig. 4D), dándole a la cabezuela una apariencia seca. Mientras que en Inuleae son herbáceos o membranáceos, como en la mayoría de las Asteraceae (Fig. 4A, E, F, H, I). A veces son escariosos en el margen (p. ej. Calenduleae, Fig. 4F), o, en la mayor parte de la superficie (p. ej. Anthemideae, Fig. 4G), aunque la parte media sea herbácea o membranácea. En cuanto a la topología de las brácteas involucrales, es posible identificar dos tipos principales: laminares y naviculares. Los filarios laminares generalmente son aplanados (Fig. 4D-G), en ocasiones ligeramente cóncavos y son los más comunes en Asteraceae, mientras que los naviculares (Fig. 4H) son exclusivos de la tribu Perityleae. Cardueae también tiene un carácter diagnóstico en las brácteas involucrales: el ápice modificado en una espina (Fig. 1F) (Cuadro 5).
En algunos casos el involucro está rodeado por un conjunto de brácteas que difieren de los filarios en longitud, forma o indumento y se conocen como calículo (Roque et al., 2009); puede encontrarse en algunas Cichorieae, Senecioneae y Tageteae (Fig. 4I). La forma del involucro suele emplearse para diferenciar géneros y especies, ya que en las tribus es sumamente variable. Las formas más comunes son campanulado (Fig. 4A), hemisférico (Fig. 4B), cilíndrico (Fig. 4C) y tubinado. El grado de fusión de los filarios tiene importancia taxonómica. Por ejemplo, en las Coreopsideae los filarios de la serie externa están fusionados en la base (Fig. 4B), mientras que los involucros connados en toda su extensión, a veces de la parte media hacia la base, son exclusivos de la tribu Tageteae (Fig. 4I), al igual que la presencia de glándulas oleíferas, pústulas o cavidades secretoras de aroma anisado. Algo similar ocurre en Anthemideae, cuyos miembros presentan glándulas sésiles (Fig. 4G) (Cuadro 5).
Androceo
Las anteras de Asteraceae fusionadas (estambres sinantéreos) son el principal carácter diagnóstico de la familia. No obstante, presentan variaciones en la forma y longitud de los apéndices apicales y en la base, la coloración de las tecas y la presencia o ausencia de glándulas (Cuadro 6). El ápice de las anteras puede ser apiculado (común en Cardueae y Gochnatieae, Fig. 5A), deltado (p. ej. Coreopsideae, Fig. 5B), lanceolado (p. ej. Vernonieae, Fig. 5), oblongo (p. ej. Anthemideae, Fig. 5D), ovado (p. ej. Helenieae, Fig. 5E), o una combinación entre dos de las formas antes mencionadas. En algunos grupos están reducidos (p. ej. Eupatorieae, Fig. 5F) o ausentes (Cuadro 6). Aun cuando la forma de los apéndices apicales no suele emplearse para separar grupos, su presencia o ausencia ha sido utilizada para diferenciar géneros de una misma tribu, p. ej., Piqueria Cav. y Stevia Cav., en Eupatorieae (Villaseñor, 1987).
Cuadro 6: Características de las anteras en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas.
| Tribus | Coloración | Indumento | Apéndices apicales | Base | Collar |
|---|---|---|---|---|---|
| Anthemideae | blanquecinas | glabras | ovados o deltados | corto sagitada | balaustriforme |
| Arctotideae* | blanquecinas o amarillentas | glabras | deltados | corto sagitada | ausente |
| Astereae | blanquecinas | glabras | lanceolados | corto sagitada | ausente |
| Bahieae | blanquecinas | a veces glandulares en los apéndices apicales | ovados | sagitada | ausente |
| Calenduleae* | blanquecinas o amarillentas | glabras | deltados | caudada | cilíndrico |
| Cardueae | blanquecinas, a veces con tintes rosados o morados | glabras | apiculados, a veces lanceolados | caudada | cilíndrico, a veces ausente |
| Chaenactideae | blanquecinas, a veces con tintes rosados o pardos | glabras | ovados | corto sagitada | ausente |
| Cichorieae | blanquecinas o amarillentas, a veces azuladas o rosadas | glabras | lanceolados | caudada, a veces sagitada | ausente |
| Coreopsideae | negras | glabras | deltados u ovados, a veces carinados | corto sagitada | cilíndrico |
| Eupatorieae | blanquecinas | glabras | reducidos, a veces deltados, lanceolados, ovados u oblongos | reducida, a veces corto sagitada | cilíndrico, a veces inconspicuo |
| Gnaphalieae | blanquecinas | glabras | ovados o lanceolados | caudada | cilíndrico, a veces reducido o ausente |
| Gochnatieae | blanquecinas | a veces pilosas en la base | apiculados | caudada | ausente |
| Helenieae | negras | glabras | deltados u ovados, a veces carinados | corto sagitada | cilíndrico |
| Heliantheae | negras | glabras | ovados o deltados | sagitada | cilíndrico |
| Inuleae | blanquecinas | glabras | lanceolados, ovados o deltados | caudada | ausente |
| Liabeae | blanquecinas o amarillentas | glabras | lanceolados, ovados u oblongos | sagitada, a veces caudada | ausente |
| Madieae | moradas | glabras | ovados o deltados | sagitada, a veces caudada | cilíndrico o ausente |
| Millerieae | negras | glabras | lanceolados u ovados | corto sagitada | ausente |
| Mutisieae | blanquecinas, a veces con tintes rosados | glabras | oblongos o lanceolados | caudada o sagitada, a veces con papilas | ausente |
| Nassauvieae | blanquecinas | glabras | lanceolados | caudada | ausente |
| Neurolaeneae | negras | glabras | ovados o deltados | corto sagitada | cilíndrico |
| Onoserideae | blanquecinas | glabras | lanceolados | caudada | ausente |
| Perityleae | blanquecinas o amarillentas | glabras | ovados, a veces clavados | sagitada, a veces caudada | cilíndrico, a veces ausente |
| Senecioneae | blanquecinas | glabras | ovados, lanceolados u oblongos | sagitada, a veces caudada | cilíndrico o balaustriforme |
| Tageteae | blanquecinas o amarillas, a veces rojizas, moradas o negras | glabras | ovados, deltados o lanceolados | sagitada | cilíndrico |
| Vernonieae | blanquecinas, a veces con tintes rosados o morados | glandulares, a veces glabras | lanceolados, deltados u ovados | sagitada, a veces caudada | ausente |
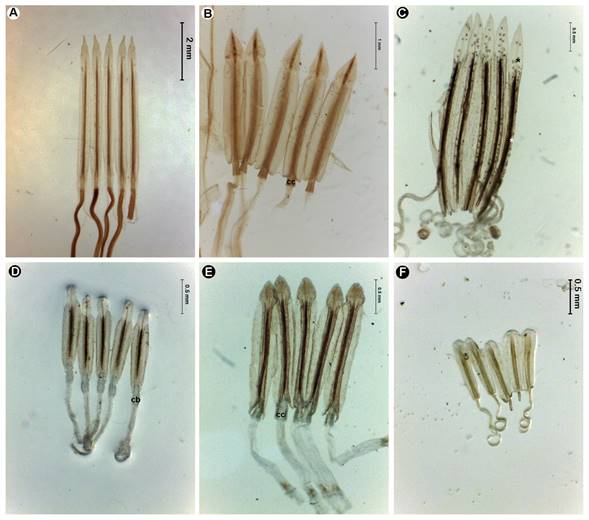
Figura 5: Androceo de Asteraceae. A. Gochnatieae, Tehuasca magna (M.C. Johnst. ex Cabrera) Panero, anteras con apéndices apicales apiculados y base caudada; B. Coreopsideae, Dahlia coccinea Cav., anteras con apéndices apicales deltados, base corto sagitada y collar cilíndrico; C. Vernonieae, Vernonia serratuloides Kunth, anteras con glándulas sésiles (*), apéndices apicales lanceolados y base sagitada; D. Anthemideae, Achillea millefolium L., anteras con apéndices apicales oblongos, base corto sagitada y collar balaustriforme; E. Helenieae, Helenium mexicanum Kunth, anteras con apéndices apicales ovados, base corto sagitada y collar cilíndrico; F. Eupatorieae, Koanophyllon longifolium (B.L. Rob.) R.M. King & H. Rob., anteras con apéndices apicales y base reducidos. cb=collar balaustriforme; cc=collar cilíndrico. Fotos: Rosario Redonda-Martínez.
La base puede ser caudada (Fig. 5A), corta (Fig. 5B, D-E) o largo sagitada (Fig. 5C), e incluso estar reducida como los apéndices (Fig. 5F) (Cuadro 6). En claves de identificación recientes la forma de la base de las anteras generalmente es referida como sagitada o caudada (Pruski, 2018; Redonda-Martínez, 2019), sin hacer mayor referencia a la longitud relativa (corta o larga). Ambos términos han sido retomados de Robinson (1983), quien aclaró que independientemente de la forma y longitud que tenga la base (medida a partir del punto de inserción de las tecas con el filamento), las anteras caudadas carecen de tejido fértil y por lo tanto de polen, mientras que las sagitadas presentan tejido fértil y producen polen en toda la teca. Previo a la publicación de ese trabajo, la base de las anteras era descrita como triangular, aflechada, cordada o hastada, y si su longitud superaba 1 mm, se consideraban caudadas. Por ello, Robinson (1983) propuso emplear el término calcarada o espolonada para referirse a las anteras cuyas bases fueran prominentes, pudiendo ser también caudadas. Esto comúnmente ocurre en varias tribus de las subfamilias Cichorioideae y Gochnatioideae (Funk et al., 2009), en las que incluso algunas especies tienen tricomas eglandulares en la base (p. ej. Gochnatieae (Roque et al., 2009; Redonda-Martínez, 2019)).
El collar de la antera corresponde a una extensión proximal del conectivo ubicado en el extremo distal del filamento (Roque et al., 2009). Esta área se caracteriza por el engrosamiento de las paredes celulares y la forma que adquieren, la cual contrasta con las células del filamento, aunque el collar está ausente en varios grupos (Fig. 5A, C, F). En aquellas tribus que sí presentan, los collares cilíndricos son los más comunes (p. ej. Coreopsideae (Fig. 5B), Eupatorieae, Heliantheae, Senecioneae y Tageteae), también hay balaustriformes (p. ej. Anthemideae (Fig. 5D) y algunas Senecioneae) (Cuadro 2), que se denominan así por tener apariencia similar a los balaustres del barandal de una escalera (Cuadro 6).
Gineceo
Asteraceae presenta ovario ínfero, estilo filiforme con ramas bífidas, las cuales varían en forma, longitud e indumento, características que guardan cierta relación entre sí. Por ejemplo, cuando un estilo presenta ramas largas, suelen ser lanceoladas, agudas y pilosas en toda su extensión (p. ej. Cichorieae, Fig. 6A), ya sea por arriba o debajo del punto de bifurcación. En Cardueae las ramas del estilo son glabras; no obstante, debajo del punto de bifurcación tienen un anillo colector. Los miembros de Eupatorieae presentan ramas clavadas, generalmente aplanadas, glabras o a veces papilosas, en toda su extensión. También es común que se diferencie claramente la región estigmática fértil, la cual comprende del punto de bifurcación hacia la parte media de las ramas (Fig. 6B), que incluso pueden ser más largas que el estilo (Fig. 6C) (Cuadro 7).
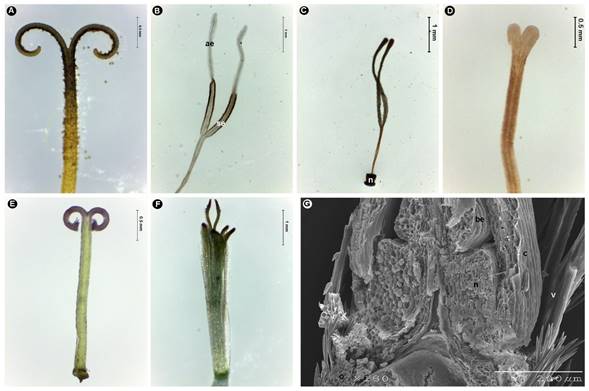
Figura 6: Estilo de Asteraceae. A. Cichorieae, Taraxacum officinale F.H. Wigg., ramas del estilo supinadoras, lanceoladas, agudas, pilosas en toda su extensión; B. Eupatorieae, Ageratina mairetiana (DC.) R.M. King & H. Rob., ramas del estilo supinadoras, lanceoladas, aplanadas, papilosas, con la superficie estigmática fértil (se) y apéndices apicales estériles (ae); C. Eupatorieae, Koanophyllon longifolium (B.L. Rob.) R.M. King & H. Rob., ramas del estilo supinadoras, clavadas, glabras, más largas que el estilo, mostrando el nectario cilíndrico (n) y la base del estilo completamente inmersa en él; D. Gochnatieae, Nahuatlea smithii (B.L. Rob. & Greenm.) V.A. Funk, ramas del estilo bifurcadas, cortas, obtusas, glabras; E. Anthemideae, Achillea millefolium L., ramas del estilo supinadoras, truncadas, pilosas en el ápice; F. Astereae, Symphyotrichum subulatum (Michx.) G.L. Nesom, ramas del estilo pronadoras, subuladas, pilosas por arriba del punto de bifurcación; G. Calenduleae, Calendula officinalis L., estilo de una flor central funcionalmente masculina, carente de ramas; H. Cardueae, Cirsium velatum (S. Watson) Petr., base del estilo dispuesta por encima del nectario (n). Fotos: Rosario Redonda-Martínez.
Cuadro 7: Características del estilo en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas.
| Tribus | Orientación de las ramas | Forma | Longitud relativa | Indumento |
|---|---|---|---|---|
| Anthemideae | supinadoras | truncadas | cortas o largas | pilosas solo en el ápice |
| Arctotideae* | supinadoras | subuladas | cortas | glabras |
| Astereae | bifurcadas o supinadoras en flores unisexuales; pronadoras en flores bisexuales | agudas en flores unisexuales; subuladas o triangulares y aplanadas en flores bisexuales | cortas o largas | glabras en flores unisexuales, pilosas por arriba del punto de bifurcación en flores bisexuales |
| Bahieae | supinadoras | lanceoladas, aplanadas o agudas, a veces truncadas | largas o cortas | papilosas por arriba del punto de bifurcación o solo en el ápice |
| Calenduleae* | supinadoras en las flores periféricas; ausentes en las flores centrales | aplanadas en las flores periféricas | largas | glabras, ápice del estilo piloso en las flores centrales |
| Cardueae | supinadoras | lineares o lanceoladas | largas | glabras, a veces papilosas; con un anillo colector por debajo del punto de bifurcación |
| Chaenactideae | supinadoras | lineares, agudas | cortas | pilosas o papilosas, por arriba del punto de bifurcación |
| Cichorieae | supinadoras | lanceoladas o agudas | largas | pilosas o papilosas desde antes del punto de bifurcación |
| Coreopsideae | supinadoras | truncadas o peniciladas | cortas o largas | pilosas o papilosas en el ápice |
| Eupatorieae | supinadoras | clavadas, obtusas | largas | glabras o papilosas por arriba del punto de bifurcación |
| Gnaphalieae | supinadoras | lineares, a veces truncadas | cortas o largas | pilosas en el ápice |
| Gochnatieae | bifurcadas o ligeramente erectas | teretes, obtusas | cortas | glabras |
| Helenieae | supinadoras | truncadas | cortas | pilosas en el ápice |
| Heliantheae | supinadoras | lineares, lanceoladas, agudas o redondeadas | largas o cortas | papilosas o pilosas por arriba del punto de bifurcación |
| Inuleae | supinadoras | filiformes, lanceoladas | largas | pilosas desde antes del punto de bifurcación |
| Liabeae | supinadoras | lanceoladas, agudas | largas | pilosas o papilosas desde antes del punto de bifurcación |
| Madieae | supinadoras | subuladas o truncadas | cortas | pilosas o papilosas, por arriba del punto de bifurcación |
| Millerieae | supinadoras, a veces erectas; reducidas o ausentes en flores funcionalmente masculinas | lanceoladas o peniciladas, agudas | cortas o largas | pilosas por arriba del punto de bifurcación, a veces glabras |
| Mutisieae | bifurcadas | ovadas, rómbicas, lanceoladas o truladas | cortas o largas | papilosas por arriba de la bifurcación, rara vez glabras |
| Nassauvieae | bifurcadas o supinadoras | obtusas o truncadas | cortas o largas | pilosas en el ápice |
| Neurolaeneae | supinadoras | lanceoladas, agudas | cortas o largas | glabras o papilosas por arriba del punto de bifurcación, a veces solo en el ápice |
| Onoserideae | erectas o ligeramente supinadoras | obtusas | cortas | papilosas en el ápice |
| Perityleae | supinadoras | ahusadas u obtusas | cortas | pilosas o papilosas por arriba del punto de bifurcación, a veces solo en el ápice |
| Senecioneae | supinadoras | lineares, agudas, truncadas o peniciladas | cortas o largas | papilosas por arriba del punto de bifurcación, a veces solo en el ápice o glabras |
| Tageteae | supinadoras | lineares o lanceoladas, agudas | cortas o largas | papilosas o pilosas por arriba del punto de bifurcación, a veces glabras |
| Vernonieae | supinadoras | lanceoladas, filiformes o agudas | largas | pilosas desde antes del punto de bifurcación |
Contrario a lo anterior, en Gochnatieae, Mutisieae, Nassauvieae y Onoserideae, las ramas son cortas, erectas o bifurcadas, obtusas o truncadas, glabras, papilosas o pilosas solo en el ápice (Fig. 6D). En otras tribus son truncadas y el indumento se encuentra únicamente en el área distal (p. ej. Anthemideae (Fig. 6E)). Los miembros de Astereae tienen ramas subuladas o triangulares en las flores bisexuales (Fig. 6F); otro carácter exclusivo de esta tribu es la orientación, ya que son pronadoras. Es decir, la superficie estigmática de una se ubica frente a la otra y los ápices pueden entrar en contacto o no, de tal manera que el espacio que hay al interior de las ramas forma un rombo o una elipse (Fig. 6F). Las 21 tribus restantes presentan ramas supinadoras, tanto en las flores femeninas como en las bisexuales, cuya principal diferencia con respecto a las pronadoras es que la superficie interna está orientada hacia arriba, quedando totalmente expuesta al medio (Fig. 6A, E). En las flores funcionalmente masculinas (p. ej. Calenduleae), las ramas del estilo pueden estar reducidas o ausentes (Fig. 6G; Cuadro 7).
A lo largo de la historia, las características estilares y en particular las diferencias en la forma, longitud e indumento de las ramas y estigmas han sido empleadas para delimitar tribus de Asteraceae (Cassini, 1819; Bentham, 1873; Small, 1919). Estos caracteres continúan vigentes y al incorporar la orientación que tienen las ramas en las flores bisexuales, el reconocimiento de algunos grupos resulta más sencillo.
La base del estilo también muestra diferencias en cuanto a nivel de inserción en el nectario. Cuando está completamente inmersa (p. ej. Eupatorieae (Fig. 6C)), se denomina anular, parcial si solo una parte está inserta en él, y apical cuando se encuentra arriba del nectario (Fig. 6H). El término estilopodio hace referencia a la asociación que ocurre entre la base del estilo y el nectario (Roque et al., 2009). Su uso no es muy común en las descripciones, salvo en Eupatorieae, tribu en la que es de importancia taxonómica para reconocer géneros y especies (King y Robinson, 1987).
Cipselas
Las cipselas son potencialmente útiles para delimitar grupos al igual que el vilano, por ello han sido empleadas desde las primeras clasificaciones tribales de Asteraceae (Cassini, 1819; Bentham, 1873; Small, 1919). Si bien no es común encontrar el término cipsela en las claves de identificación (p. ej. McVaugh, 1984; Villaseñor, 1987; Rzedowski y Calderón de Rzedowski, 2008; Bonifacino y Funk, 2018) o literatura especializada en Compositae (Funk et al., 2009; Roque et al., 2009), su empleo para referirse a los frutos de las asteráceas (p. ej. Pruski, 2018; Redonda-Martínez, 2018, 2019, 2020, 2022) ha ido ganando terreno. Aunque en algunos casos aquenio y cipsela se usan indistintamente (p. ej. Funk et al., 2009; Roque et al., 2009), lo ideal es utilizar el nombre correcto, pues, aunque ambos son frutos secos, tienen origen distinto, el aquenio deriva de un ovario súpero, mientras que la cipsela de un ovario ínfero (Marzinek et al., 2008). Los caracteres de las cipselas con mayor utilidad taxonómica para delimitar o identificar tribus, géneros y especies son la forma, el número de costillas, la presencia de estrías o pliegues, presencia o ausencia de vilano, fitomelanina, carpopodio e indumento (Cuadro 8).
Cuadro 8: Características de las cipselas en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas. NA = No aplica.
| Tribus | Morfismo | Fitomelanina | Forma | Número de costillas | Carpopodio | Rostro | Indumento |
|---|---|---|---|---|---|---|---|
| Anthemideae | homomórficas, a veces dimórficas | ausente | obovoides, columnares, elipsoides o prismáticas | 3-10 costilladas; a veces 3-5 anguladas o aladas | anular, a veces reducido | ausente | glándulas sésiles |
| Arctotideae* | homomórficas | ausente | elipsoides u obovoides | 3-5 costilladas; a veces 4-5 anguladas | ausente | ausente | seríceas |
| Astereae | homomórficas, a veces dimórficas | ausente | teretes o prismáticas; a veces obovoides, fusiformes o aplanadas u obcompresas | 3-5 costilladas; a veces 2-4 anguladas | anular, simétrico, reducido | rara vez presente | glabras, a veces pilosas o glandulares |
| Bahieae | homomórficas | presente | clavadas, obpiramidales o triquetras | 3-5 anguladas, estriadas | anular, simétrico, reducido cuando presente | ausente | pilosas o seríceas en las costillas |
| Calenduleae* | heteromórficas | ausente | uncinadas, a veces naviculares, arqueadas, incurvadas, columnares, comprimidas obcompresas o aplanadas | NA | ausente | reducido o ausente | glabras, superficie fenestrada; a veces tuberculada o rugulada |
| Cardueae | homomórficas | a veces presente | elipsoides o teretes | ausentes, a veces estriadas | anular, simétrico; a veces reducido o ausente | ausente | glabras |
| Chaenactideae | homomórficas | presente | fusiformes, s veces cilíndricas o compresas | 8-20 anguladas, a veces estriadas | ausente | ausente | setulosas o seríceas |
| Cichorieae | homomórficas | ausente | clavadas, columnares, elipsoides, fusiformes o prismáticas | 3-15 costilladas, a veces 4-5 anguladas; en ocasiones estriadas | anular, simétrico o asimétrico; a veces reducido o ausente | presente, a veces reducido o ausente | glabras; superficie tuberculada, muricada o espinulosa |
| Coreopsideae | homomórficas, a veces dimórficas o heteromórficas | presente | obcompresas, fusiformes o cilíndrico claviformes; a veces aplanadas, triquetras, aladas o suberificadas en las costillas | 2-5 costilladas o anguladas; a veces estriadas | anular, simétrico; a veces reducido, asimétrico o ausente | presente, a veces reducido o ausente | glabras, pulverulentas o esparcidamente pilosas o setosas; a veces solo en las costillas; en ocasiones la superficie tuberculada |
| Eupatorieae | homomórficas | presente | fusiformes o prismáticas, rara vez comprimidas | 2-10 costilladas; a veces 4-5 anguladas | anular, simétrico; a veces reducido, cuadrangular o asimétrico | ausente | pilosas, setosas o glandulares, a veces glabras |
| Gnaphalieae | homomórficas | ausente | elipsoidales, obovoides u ovoides | 2-5 costilladas | ausente | ausente | glabras, a veces papilosas, setosas, pilosas o glandulares |
| Gochnatieae | homomórficas | ausente | teretes o prismáticas | 4-5 costilladas | ausente | ausente | pilosas o seríceas |
| Helenieae | homomórficas | presente | prismáticas o claviformes | 4-5 costilladas, a veces 4 anguladas | ausente | ausente | seríceas, en ocasiones solo en las costillas |
| Heliantheae | homomórficas, a veces dimórficas | presente | obovoides, aplanadas o triquetras | 3-5 costilladas, a veces 8-10 costillas | anular, reducido, a veces ausente | ausente | glabras; superficie estriada |
| Inuleae | homomórficas | ausente | elipsoides, cilíndricas, fusiformes o claviformes | 6-8 costilladas, a veces 4 anguladas | anular, simétrico | ausente | pilosas o glandulares |
| Liabeae | homomórficas | ausente | teretes o prismáticas | 2-10 costilladas, a veces anguladas o estriadas | anular, simétrico o asimétrico; a veces reducido o ausente | ausente | pilosas o glabrescentes |
| Madieae | homomórficas | presente | compresas, obcompresas, teretes, obpiramidales, claviformes o fusiformes | 3 anguladas | ausente | ausente | glabras o pilosas |
| Millerieae | homomórficas | presente | prismáticas, obcónicas, obovoides u obpiramidales; a veces esféricas | 3-4 anguladas | anular, simétrico; a veces ausente | ausente | glabras, superficie estriada o tuberculada |
| Mutisieae | dimórficas, a veces heteromórficas | ausente | elipsoides o prismáticas, a veces subteretes o aplanadas | 4-10 costilladas | anular, simétrico; a veces reducido o ausente | presente, a veces reducido o ausente | pilosas o glabrescentes, a veces seríceas o glandulares |
| Nassauvieae | homomórficas | a veces presente | elipsoides, fusiformes o teretes | ausentes, generalmente estriadas | anular, simétrico | ausente, a veces reducido en Trixis P. Browne | pilosas o glabrescentes |
| Neurolaeneae | homomórficas, rara vez dimórficas | presente | teretes o prismáticas | 5 costilladas | anular, simétrico cuando presente | ausente | pilosas o glabrescentes |
| Onoserideae | homomórficas | ausente | elipsoides o fusiformes | 4-6 costilladas | ausente | ausente | esparcidamente pilosas o glabrescentes |
| Perityleae | homomórficas | presente | teretes, biconvexas u obpiramidales; a veces aladas | 2-4 anguladas, en ocasiones los ángulos suberificados (Galeana La Llave) | ausente | ausente | pilosas o seríceas; a veces solo en las costillas; superficie estriada |
| Senecioneae | homomórficas | ausente | oblongas, ovoides o claviformes, ocasionalmente aplanadas | 4-12 costilladas | ausente | ausente | glabras o pilosas; a veces seríceas o glandulares; superficie lisa o estriada |
| Tageteae | homomórficas | presente | claviformes o fusiformes; a veces prismáticas o aplanadas | 5-10 costilladas | anular, simétrico; a veces asimétrico o reducido | ausente | glabras o esparcidamente pilosas |
| Vernonieae | homomórficas | presente en Stramentopappus H. Rob. & V.A. Funk | prismáticas o tereres | 8-10 costilladas, a veces 4-5 anguladas | anular, simétrico o asimétrico; a veces reducido o ausente | ausente | pilosas, glandulares o seríceas, a veces glabras |
La forma suele describirse con base en figuras planas y tridimensionales. Así las cipselas pueden ser lineares (Fig. 7A-C), fusiformes (Fig. 7 D, E), clavadas (Fig. 7F-H), uncinadas (Fig. 7I), obovoides (Figs. 7J; 8A-C), elipsoides (Fig. 8D), teretes (Fig. 8E) o esféricas (Fig. 8F). Considerando el número de costillas pueden ser triquetas (Figs. 7F; 8G) o prismáticas, cuando tienen más de cuatro (Fig. 8H-J). En este último caso a veces es necesario hacer cortes transversales para cuantificarlas correctamente y corroborar que sean costillas, ya que en ocasiones tienen estrías (Fig. 8A) o pliegues (Fig. 8D).

Figura 7: Cipselas y vilano de Asteraceae. A. Tageteae, Tagetes lunulata Ortega, cipsela linear clavada glabrescente, con fitomelanina, vilano radial de escamas y una arista; B. Eupatorieae, Brickellia leonis Rzed. & Calderón, cipsela linear fusiforme pilosa, con fitomelanina, vilano radial de cerdas plumosas; C. Gochnatieae, Nahuatlea smithii (B.L. Rob. & Greenm.) V.A. Funk, cipsela linear fusiforme pilosa, sin fitomelanina, vilano radial multiseriado de cerdas capilares; D. Coreopsideae, Bidens L., cipselas fusiformes setulosas en el ápice, con fitomelanina; vilano radial de aristas retrorsamente barbeladas; E. Eupatorieae, Oxylobus arbutifolius (Kunth) A. Gray, cipsela prismática fusiforme setulosa, con fitomelanina, vilano radial de escamas laciniadas; F. Bahieae, Achyropappus anthemoides Kunth, cipsela clavada esparcidamente pilosa, con fitomelanina; vilano radial de escamas con una costilla media conspicua; G. Bahieae, Schkuhria pinnata (Lam.) Kuntze ex Thell., cipsela clavada serícea, con fitomelanina; vilano radial de escamas erosas y escamas aristadas; H. Anthemideae, Matricaria chamomilla L., cipsela prismática claviforme glabra, sin fitomelanina; vilano radial coroniforme; I. Calenduleae, Calendula officinalis L., cipselas uncinadas fenestradas, glabras, sin fitomelanina; vilano ausente; J. Coreopsideae, Coreopsideae, Heterosperma pinnatum Cav., cipselas obovoides aplanadas, a veces biconvexas, suberificadas en las costillas, con fitomelanina; vilano bilateral de aristas retrorsamente barbeladas. Los asteriscos (*) señalan el carpopodio y el signo más (+), la presencia de una costilla media en los elementos del vilano. Fotos: Rosario Redonda-Martínez.

Figura 8: Cipselas y vilano de Asteraceae. A. Heliantheae, Helianthus annuus L., cipsela obovoide estriada, con fitomelanina; vilano bilateral deciduo; B. Astereae, Heterotheca inuloides Cass., cipsela fusiforme obovoide, pilosa, con fitomelanina; vilano radial persistente, de cerdas capilares; C. Heliantheae, Delilia biflora (L.) Kuntze, cipsela obovoide formando un complejo con el involucro, vilano ausente; D. Cardueae, Cirsium velatum (S. Watson) Petr., cipsela elipsoide con pliegues, glabra, sin fitomelanina, con callo apical (ca); vilano ausente; E. Neurolaeneae, Neurolaena lobata (L.) Cass., cipsela terete esparcidamente pilosa, con fitomelanina; vilano radial persistente, de cerdas capilares; F. Millerieae, Smallanthus maculatus (Cav.) H. Rob., cipsela esférica glabra, estriada, con fitomelanina, formando un complejo con un filario y una pálea navicular; vilano ausente; G. Heliantheae, Lasianthaea fruticosa (L.) K.M. Becker var. michoacana (S.F. Blake) K.M. Becker, cipsela triquetra glabra, con fitomelanina; vilano bilateral persistente, de aristas glabras; H. Millerieae, Melampodium divaricatum (Rich.) DC., cipsela prismática tuberculada, con fitomelanina, vilano ausente; I. Senecioneae, Roldana hederifolia (Hemsl.) H. Rob. & Brettell, cipsela fusiforme prismática, costillada, sin fitomelanina; vilano radial deciduo, de cerdas capilares; J. Vernonieae, Vernonia alamanii DC., cipsela prismática costillada, con glándulas, sin fitomelanina; vilano radial persistente, biseriado de cerdas ensanchadas reducidas y cerdas capilares; K. Cichorieae, Taraxacum officinale F.H. Wigg., cipsela fusiforme, costillada, tuberculada en el ápice, sin fitomelanina, rostrada; vilano radial de cerdas capilares; L. Coreopsideae, Cosmos caudatus Kunth, cabezuela con cipselas heteromórficas fusiformes, rostradas; vilano radial de aristas retrorsamente barbeladas. Los asteriscos (*) señalan el carpopodio, ca=callo apical, f=filario, p=pálea, r=rostro. Fotos: Rosario Redonda-Martínez.
En las Asteraceae que tienen cabezuelas homógamas las cipselas son homomórficas, es decir que todos los frutos son similares (Cuadro 8). Mientras que en aquellas que presentan cabezuelas heterógamas suele haber dimorfismo; de tal manera que las cipselas producidas por las flores periféricas son diferentes de las que se originan de las flores centrales (p. ej. Mutisieae). Además, existe heteromorfia o heterocarpia; es decir, la producción de frutos completamente distintos en una misma cabezuela, como ocurre en Calenduleae (Fig. 7I) y en menor proporción en algunas Coreopsideae (p. ej. Heterosperma pinnatum Cav. (Fig. 7J) y Cosmos caudatus Kunth (Fig. 8L) (Cuadro 8)).
Cuadro 8: Características de las cipselas en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas.
| Tribus | Morfismo | Fitomelanina | Forma | Número de costillas | Carpopodio | Rostro | Indumento |
|---|---|---|---|---|---|---|---|
| Anthemideae | homomórficas, a veces dimórficas | ausente | obovoides, columnares, elipsoides o prismáticas | 3-10 costilladas; a veces 3-5 anguladas o aladas | anular, a veces reducido | ausente | glándulas sésiles |
| Arctotideae* | homomórficas | ausente | elipsoides u obovoides | 3-5 costilladas; a veces 4-5 anguladas | ausente | ausente | seríceas |
| Astereae | homomórficas, a veces dimórficas | ausente | teretes o prismáticas; a veces obovoides, fusiformes, aplanadas u obcompresas | 3-5 costilladas; a veces 2-4 anguladas | anular, simétrico, a veces reducido | rara vez presente | glabras, a veces pilosas o glandulares |
| Bahieae | homomórficas | presente | clavadas, obpiramidales o triquetras | 3-5 anguladas, estriadas | anular, simétrico, reducido cuando presente | ausente | pilosas o seríceas en las costillas |
| Calenduleae* | heteromórficas | ausente | uncinadas, a veces naviculares, arqueadas, incurvadas, columnares, comprimidas obcompresas o aplanadas | 2-4 costilladas, aladas | ausente | reducido o ausente | glabras, superficie fenestrada; a veces tuberculada, estriada, o rugulada |
| Cardueae | homomórficas | a veces presente | elipsoides o teretes | ausentes, a veces estriadas | anular, simétrico; a veces reducido o ausente | ausente | glabras |
| Chaenactideae | homomórficas | presente | fusiformes, a veces cilíndricas o compresas | 8-20 anguladas, a veces estriadas | ausente | ausente | setulosas o seríceas |
| Cichorieae | homomórficas | ausente | clavadas, columnares, elipsoides, fusiformes o prismáticas | 3-15 costilladas, a veces 4-5 anguladas; en ocasiones estriadas | anular, simétrico o asimétrico; a veces reducido o ausente | presente, a veces reducido o ausente | glabras; superficie tuberculada, muricada o espinulosa |
| Coreopsideae | homomórficas, a veces dimórficas o heteromórficas | presente | obcompresas, fusiformes o cilíndrico claviformes; a veces aplanadas, triquetras, aladas o suberificadas en las costillas | 2-5 costilladas o anguladas; a veces estriadas | anular, simétrico; a veces reducido, asimétrico o ausente | presente, a veces reducido o ausente | glabras, pulverulentas o esparcidamente pilosas o setosas; a veces solo en las costillas; en ocasiones la superficie tuberculada |
| Eupatorieae | homomórficas | presente | fusiformes o prismáticas, rara vez comprimidas | 2-10 costilladas; a veces 4-5 anguladas | anular, simétrico; a veces reducido, cuadrangular o asimétrico | ausente | pilosas, setosas o glandulares, a veces glabras |
| Gnaphalieae | homomórficas | ausente | elipsoidales, obovoides u ovoides | 2-5 costilladas | ausente | ausente | glabras, a veces papilosas, setosas, pilosas o glandulares |
| Gochnatieae | homomórficas | ausente | teretes o prismáticas | 4-5 costilladas | ausente | ausente | pilosas o seríceas |
| Helenieae | homomórficas | presente | prismáticas o claviformes | 4-5 costilladas, a veces 4 anguladas | ausente | ausente | seríceas, en ocasiones solo en las costillas |
| Heliantheae | homomórficas, a veces dimórficas | presente | obovoides, aplanadas o triquetras | 3-5 costilladas, a veces 8-10 costillas | anular, simétrico; a veces reducido o ausente | ausente | glabras; superficie estriada |
| Inuleae | homomórficas | ausente | elipsoides, cilíndricas, fusiformes o claviformes | 6-8 costilladas, a veces 4 anguladas | anular, simétrico | ausente | pilosas o glandulares |
| Liabeae | homomórficas | ausente | teretes o prismáticas | 2-10 costilladas, a veces anguladas o estriadas | anular, simétrico o asimétrico; a veces reducido o ausente | ausente | pilosas o glabrescentes |
| Madieae | homomórficas | presente | compresas, obcompresas, teretes, obpiramidales, claviformes o fusiformes | 3 anguladas | ausente | ausente | glabras o pilosas |
| Millerieae | homomórficas | presente | prismáticas, obcónicas, obovoides u obpiramidales; a veces esféricas | 3-4 anguladas | anular, simétrico; a veces ausente | ausente | glabras, superficie estriada o tuberculada |
| Mutisieae | dimórficas, a veces heteromórficas | ausente | elipsoides o prismáticas, a veces subteretes o aplanadas | 4-10 costilladas | anular, simétrico; a veces reducido o ausente | presente, a veces reducido o ausente | pilosas o glabrescentes, a veces seríceas o glandulares |
| Nassauvieae | homomórficas | a veces presente | elipsoides, fusiformes o teretes | ausentes, generalmente estriadas | anular, simétrico | ausente, a veces reducido en Trixis P. Browne | pilosas o glabrescentes |
| Neurolaeneae | homomórficas, rara vez dimórficas | presente | teretes o prismáticas | 5 costilladas | anular, simétrico cuando presente | ausente | pilosas o glabrescentes |
| Onoserideae | homomórficas | ausente | elipsoides o fusiformes | 4-6 costilladas | ausente | ausente | esparcidamente pilosas o glabrescentes |
| Perityleae | homomórficas | presente | teretes, biconvexas u obpiramidales; a veces aladas | 2-4 anguladas, en ocasiones los ángulos suberificados (Galeana La Llave) | ausente | ausente | pilosas o seríceas; a veces solo en las costillas; superficie estriada |
| Senecioneae | homomórficas | ausente | oblongas, ovoides o claviformes, ocasionalmente aplanadas | 4-12 costilladas | ausente | ausente | glabras o pilosas; a veces seríceas o glandulares; superficie lisa o estriada |
| Tageteae | homomórficas | presente | claviformes o fusiformes; a veces prismáticas o aplanadas | 5-10 costilladas | anular, simétrico; a veces asimétrico o reducido | ausente | glabras o esparcidamente pilosas |
| Vernonieae | homomórficas | presente en Stramentopappus H. Rob. & V.A. Funk | prismáticas o tereres | 8-10 costilladas, a veces 4-5 anguladas | anular, simétrico o asimétrico; a veces reducido o ausente | ausente | pilosas, glandulares o seríceas, a veces glabras |
En algunos grupos, las cipselas están fusionadas con el involucro constituyendo una estructura períptera (p. ej. Delilia biflora (L.) Kuntze, Fig. 8C), o bien, con las páleas y los filarios (como en algunas Millerieae (Fig. 8F) y Heliantheae), formando un fruto complejo (“complejo cipsela”, Roque et al., 2009). Su función principal es contribuir con su dispersión, ya que por lo general este tipo de cipselas carecen de vilano y su ausencia se compensa de otra manera. A veces, el ápice de la cipsela está alargado generando una estructura rígida y linear, el rostro (Fig. 8K, L), aunque en ocasiones es corto o inconspicuo (Fig. 7D).
El carpopodio es la zona de absición de la cipsela y está formado por uno o varios estratos celulares que difieren en forma de las células de la pared del fruto (Roque et al., 2009). Es simétrico cuando forma un anillo completo y asimétrico si está incompleto o dicha estructura es similar a una letra U. En ocasiones la simetría del carpopodio puede estar relacionada con la orientación; es decir, si es plano (Figs. 7C, E, F; 8E, I) u oblicuo (Figs. 7B, D, G; 8J). A veces puede proyectarse sobre las costillas o tener otras modificaciones que son de importancia taxonómica a nivel genérico (Roque et al., 2009).
Vilano
El cáliz modificado de las flores de Asteraceae recibe el nombre de vilano; dependiendo de su posición simétrica se reconocen dos tipos: radial cuando los elementos que lo integran están dispuestos alrededor de la flor o el ápice de la cipsela (Figs. 7A-H; 8B, E, I-K) y bilateral, que se caracteriza por tener un par de elementos ubicados en lados opuestos de la corola o el ápice de la cipsela (Figs. 7J; 8G), según sea el caso (Cuadro 9). Generalmente, las estructuras que forman el vilano están dispuestas en una sola serie, ya sea de igual o distinta longitud (Figs. 7A, B, D-H; 8A, E, K), pero en ocasiones es biseriado. En este caso, la serie externa está reducida y llega a ser inconspicua, mientras que la interna suele ser 10-12 veces más larga, característica común en Liabeae y Vernonieae (Fig. 8J). También existe el vilano multiseriado, el cual presenta tres o más series de elementos que varían principalmente en longitud, como ocurre en Gochnatieae (Fig. 7C) y algunas Senecioneae. Con respecto a su duración, puede ser persistente (Figs. 7A-H; 8B, E, G, J-L) o deciduo (Fig. 8I) y en ocasiones está ausente (Fig. 8C, D, F, H) (Cuadro 9).
Cuadro 9: Características del vilano en las 26 tribus de Asteraceae con distribución en México. Los asteriscos indican las tribus introducidas, NA=No Aplica.
| Tribus | Número de series | Simetría | Persistencia | Tipo de elementos |
|---|---|---|---|---|
| Anthemideae | uniseriado, cuando presente | radial, cuando presente | persistente, cuando presente | escuámulas o coroniforme, cuando presente |
| Arctotideae* | uni o biseriado | radial | persistente | escamas |
| Astereae | uni o multiseriado | radial | persistente | cerdas capilares barbeladas o plumosas; a veces escamas, escamas aristadas, aristas o una mezcla de dos tipos elementos diferentes; rara vez ausente |
| Bahieae | uniseriado | radial | persistente | escamas con una vena media engrosada a veces proyectándose en una arista; rara vez de aristas o ausente |
| Calenduleae* | ausente | NA | NA | NA |
| Cardueae | uni o multiseriado | radial | persistente, a veces deciduo | cerdas capilares barbeladas o plumosas; a veces de escamas o ausente |
| Chaenactideae | uni o multiseriado | radial | persistente, a veces deciduo como unidad | escamas erosas, libres o fusionadas formando una corona |
| Cichorieae | uni o multiseriado | radial | persistente, a veces deciduo | cerdas capilares glabras, barbeladas o plumosas; a veces escamas, aristas o escuámulas |
| Coreopsideae | uniseriado | radial, rara vez bilateral | persistente, a veces deciduo | aristas barbeladas o escuámulas, a veces coroniforme o ausente |
| Eupatorieae | uniseriado | radial | persistente, a veces algunos elementos deciduos en grupos o como unidad | cerdas capilares barbeladas o plumosas, escamas, aristas o escamas aristadas; a veces una combinación de elementos diferentes |
| Gnaphalieae | uniseriado | radial | persistente | cerdas capilares barbeladas, a veces plumosas; en ocasiones escamas o una mezcla de cerdas capilares y escamas; rara vez ausente |
| Gochnatieae | uni o multiseriado | radial | persistente | cerdas capilares barbeladas |
| Helenieae | uniseriado | radial | persistente | escamas o escamas aristadas |
| Heliantheae | uniseriado, rara vez multiseriado | bilateral | persistente, rara vez deciduo | escamas o aristas glabras; a veces escuámulas o ausente |
| Inuleae | uniseriado | radial | persistente | cerdas capilares lisas o barbeladas; a veces escamas o ausente |
| Liabeae | biseriado | radial | persistente | escamas o cerdas reducidas en la serie externa; cerdas capilares barbeladas en la serie interna |
| Madieae | uniseriado, a veces biseriado | radial | persistente | escamas o aristas; a veces ausente |
| Millerieae | uniseriado | radial | persistente, a veces deciduo | escamas o cerdas capilares barbeladas; a veces ausente |
| Mutisieae | uniseriado | radial | persistente, a veces algunos elementos deciduos | cerdas capilares barbeladas o plumosas, iguales o subiguales, libres, a veces fusionadas en la base |
| Nassauvieae | uni o multiseriado | radial | persistente | cerdas capilares barbeladas, a veces plumosas; rara vez ausente |
| Neurolaeneae | uniseriado | radial | persistente | escamas, aristas, cerdas capilares o coroniforme; a veces ausente |
| Onoserideae | multiseriado | radial | persistente | cerdas capilares |
| Perityleae | uniseriado | radial | persistente | 2 (1 o 4) cerdas y una corona de escamas cortas; rara vez escuámulas vestigiales o ausente |
| Senecioneae | uni o multiseriado | radial | persistente o deciduo | cerdas capilares glabras o barbeladas |
| Tageteae | uniseriado | radial | persistente | escamas, aristas, escamas aristadas o escamas divididas en cerdas; a veces dos tipos de elementos combinados, rara vez ausente |
| Vernonieae | biseriado | radial | persistente, a veces deciduo | escamas o cerdas reducidas en la serie externa; cerdas capilares barbeladas en la serie interna; a veces aristas (Trichospira Kunth) o coroniforme (Struchium P. Browne) |
Independientemente de la simetría del vilano (radial o bilateral), los elementos que lo integran son variables. Los más comunes son las cerdas, que son estructruras filiformes, flexibles, barbeladas o glabras. Las diferencias en la longitud de las barbas permiten distinguir entre cerdas plumosas (Fig. 7B) y capilares (Figs. 7C; 8B, E, J, K). A veces los elementos del vilano son aciculares rígidos o un tanto flexibles y se denominan aristas (Figs. 7A, D, J; 8G, L). Pueden ser barbeladas (Figs. 7D, J; 8L) o glabras (Fig. 8G). En el primer caso, la orientación de las barbas, ya sea antrorsa (hacia arriba) o retrorsa (hacia abajo), así como su disposición en la arista (toda la superficie o solo en el ápice), suele emplearse como carácter taxonómico para identificar géneros y especies en algunas tribus.
El vilano también puede estar conformado por escamas (Fig. 7A, E-G), estructuras aplanadas cortas, triangulares, ovadas u oblongas, libres o parcialmente fusionadas y a veces laciniadas en el ápice (Fig. 7E) o con el margen eroso (Fig. 7G). En ocasiones, las escamas tienen una proyección apical de la vena media similar a un mucrón, costilla o arista. Cuando esto ocurre, se conocen como escamas aristadas (Fig. 7G), las cuales son comunes en algunas Bahieae (Fig. 7F, G) y Helenieae. También llega a ocurrir que las escamas estén reducidas o sean vestigiales; se denominan escuámulas y pueden encontrarse esporádicamente en algunas Perityleae. Por último, el vilano también puede estar reducido a una estructura anular de apariencia cartilaginosa o en ocasiones papirácea, similar a una corona; razón por la cual se denomina coroniforme (Fig. 7H); este se encuentra en algunos miembros de Anthemideae, Coreopsideae, Neurolaeneae y Vernonieae (Cuadro 9).
Con base en el análisis morfológico de los caracteres antes descritos se generó una clave de identificación para las 26 tribus presentes en México, la cual se presenta a continuación, las introducidas están señalas con asteriscos.
Clave artificial para identificar tribus de Asteraceae presentes en México
1a Cabezuelas homógamas, con todas las flores iguales, tanto en la periferia como el centro, bisexuales, en ocasiones unisexuales …………………………………………………….… 2
1b Cabezuelas heterógamas, con dos o tres tipos de flores diferentes; las periféricas femeninas o neutras; las centrales, bisexuales, a veces funcionalmente masculinas ………………………………………………………………………..……...……….. 10
2a Filarios con látex; flores bisexuales con corolas liguladas, 5 dentadas en el ápice ………………………………….………………………………………….... Cichorieae
2b Filarios sin látex; flores bisexuales, a veces unisexuales, con corolas tubulares (4-)5 lobuladas en el ápice, a veces filiformes o bilabiadas ………………………………………………. 3
3a Plantas dioicas o poligamodioicas; cabezuelas con flores unisexuales, femeninas o masculinas, en ocasiones algunas flores bisexuales en el centro …….………... Astereae
3b Plantas monoicas; cabezuelas con flores bisexuales …………………………………... 4
4a Flores con corolas bilabiadas; anteras con base caudada ...…………………..………... 5
4b Flores con corolas tubulares, tubiformes o infundibuliformes; anteras con base sagitada, rara vez caudada o reducida ……………………………………………..…...……………….... 6
5a Flores rojizas o rosadas; ramas del estilo obtusas y papilosas en el ápice …...……………………………………………………………………..…. Onoserideae
5b Flores amarillas, a veces blanquecinas, rosadas o moradas; ramas del estilo truncadas y pilosas en el ápice ………………………………………………….…… Nassauvieae
6a Corolas con lóbulos profundamente partidos, ≥½ de su longitud; anteras con apéndices apicales apiculados, base caudada …………………………………...…………...…… 7
6b Corolas con lóbulos ligeramente partidos, ≤1/3 de su longitud; anteras con apéndices apicales lanceolados, deltados u ovados, a veces reducidas; base sagitada o reducida, rara vez caudada .………...……………………………………………………..………….. 8
7a Filarios con el ápice modificado en una espina, a veces dividido en apéndices subulados o filiformes; receptáculo plano, con cerdas; lóbulos de la corola erectos; estilo con un anillo colector por debajo del punto de bifurcación; ramas largas, lineares o lanceoladas, agudas, glabras a veces papilosas …….………………………...…..………. Cardueae
7b Filarios con el ápice agudo o mucronado; receptáculo plano, glabro, alveolado; lóbulos de la corola reflexos y enrollados; estilo sin anillo colector por debajo del punto de bifurcación; ramas cortas, obtusas, glabras ………………………………. Gochnatieae
8a Cipselas con fitomelanina; vilano uniseriado, persistente o deciduo como unidad, a veces con dos tipos de elementos distintos en la misma serie; ramas del estilo glabras desde el punto de bifurcación, en ocasiones papilosas; anteras glabras, con apéndices apicales ovados o reducidos, base reducida o corto sagitada ………………………………..… 9
8b Cipselas generalmente sin fitomelanina; vilano biseriado, a veces triseriado, persistente, en ocasiones los elementos deciduos individualmente; la serie externa de escamas o cerdas ensanchadas, la interna de cerdas capilares, rara vez de aristas o coroniforme; ramas del estilo pilosas desde antes del punto de bifurcación; anteras glandulares, a veces glabras, con apéndices apicales lanceolados, ovados o deltados, base sagitada, rara vez caudada ………...…………….…………………………………………….. Vernonieae
9a Involucro uni o biseriado, campanulado o hemisférico; ramas del estilo lineares, agudas, área estigmática fértil indiferenciada; anteras con apéndices apicales ovados, base corto-sagitada; cipselas estriadas, sin carpopodio; vilano persistente o deciduo como unidad …………………………………………………………………….……. Chaenactideae
9b Involucro multiseriado, a veces uni o biseriado; cilíndrico, turbinado o campanulado; ramas del estilo clavadas; glabras o papilosas, área estigmática fértil diferenciada; anteras con apéndices apicales y base generalmente reducidos; cipselas costilladas o anguladas, con carpopodio; vilano persistente ………….……………………………………………… Eupatorieae
10a Cabezuelas disciformes, flores periféricas femeninas, con corolas filiformes; flores centrales bisexuales, en ocasiones funcionalmente masculinas ………........................ 11
10b Cabezuelas radiadas, flores periféricas femeninas o neutras, rara vez reducidas o ausentes, con corolas radiadas, a veces radiado-bilabiadas; flores centrales bisexuales, a veces funcionalmente masculinas ……………………………………………..…….. 13
11a Involucro con filarios herbáceos o membranáceos, sin estereoma ……………………………………………………………………………………….. 12
11b Involucro con filarios papiráceos, generalmente con estereoma ……….... Gnaphalieae
12a Arbustos, a veces arborescentes, rara vez hierbas perennes o anuales; flores rosadas o moradas; ramas del estilo de las flores centrales lanceoladas, supinadoras, pilosas desde antes del punto de bifurcación ………………………………..……….………... Inuleae
12b Hierbas anuales o perennes, a veces arbustos; flores blanquecinas o verdosas, ramas del estilo de las flores centrales subuladas o triangulares, pronadoras, pilosas por arriba del punto de bifurcación ………....………………………………….……………. Astereae
13a Cabezuelas solitarias; flores periféricas femeninas, multiseriadas, isomórficas o dimórficas; flores centrales funcionalmente masculinas, a veces bisexuales; cipselas dimórficas o heteromórficas ......................................................................…………... 14
13b Cabezuelas agrupadas en inflorescencias, a veces solitarias; flores periféricas femeninas o neutras, uniseriadas, isomórficas, a veces reducidas o ausentes; flores centrales bisexuales, en ocasiones funcionalmente masculinas; cipselas homomórficas, a veces dimórficas .……………………………………………………………………...……. 15
14a Flores blanquecinas o rosadas; las periféricas dispuestas en dos series, la externa con corolas radiadas o radiado-bilabiadas, la interna con corolas radiado-bilabiadas, a veces reducidas o filiformes; flores centrales con corolas bilabiadas, rara vez tubulares, pentámeras; cipselas fusiformes, elipsoides o prismáticas; vilano de cerdas capilares ………………………………………………...……………………………… Mutisieae
14b Flores anaranjadas, a veces amarillas; las periféricas dispuestas en tres o más series, isomórficas, con corolas radiadas; flores centrales con corolas tubulares, pentámeras; cipselas uncinadas, fenestradas; vilano ausente ………………….……….. Calenduleae
15a Estilo de las flores centrales con ramas subuladas o triangulares, pronadoras ………………………………………………………………………….……… Astereae
15b Estilo de las flores centrales con ramas, lanceoladas, agudas, truncadas, ahusadas o lineares, supinadoras ……………………………….……………………………..…. 16
16a Cipselas con vilano bilateral, rara vez radial, con un reducido número de elementos, de escamas o aristas; a veces fusionadas con las costillas, reminiscentes o deciduas, rara vez ausente ………..……………………………………………………………………… 17
16b Cipselas con vilano radial, de cerdas capilares, escamas, aristas, escamas aristadas, o una combinación de dos tipos de elementos, libres, rara vez fusionados formando una corona, o ausente ……………………………………………………….…………… 18
17a Involucro multiseriado, a veces uniseriado; filarios libres, similares en todas las series; receptáculo con páleas conduplicadas, persistentes, a veces acrescentes, coloridas o esclerificadas en el ápice; cipselas dimórficas u homomórficas, aplanadas, obovoides o triquetras; vilano bilateral, de escamas o aristas glabras, a veces deciduo o ausente ……….............................................................................................. Heliantheae
17b Involucro biseriado, filarios diferenciados en ambas series; los externos fusionados en la base, verdosos, generalmente crasos; los internos, coloridos, a veces verdosos, herbáceos; receptáculo con páleas laminares, deciduas, no acrescentes, con estrías paralelas pardo-anaranjadas; cipselas homomórficas o heteromórficas, obcompresas o fusiformes, a veces aplanadas, en ocasiones rostradas; vilano radial, rara vez bilateral, de aristas barbeladas, a veces escamas reminiscentes o deciduas, ocasionalmente ausente …….…........................ Coreopsideae
18a Involucro connado en toda su extensión, a veces de la parte media hacia la base; filarios con glándulas, pústulas o cavidades secretoras de aroma anisado ................... Tageteae
18b Involucro libre en toda su extensión, ocasionalmente connado solo en la base; filarios glabros o con indumento constituido por tricomas, glándulas o una combinación de ambos, sin aroma anisado ...........................................................................………….. 19
19a Receptáculo epaleáceo ……………….………………………….………………..… 20
19b Receptáculo paleáceo ……………………………………………………………..… 26
20a Pedúnculos y filarios con látex; vilano biseriado, la serie externa reducida, de escamas; la serie interna de cerdas capilares …………………………………………… Liabeae
20b Pedúnculos y filarios sin látex, a veces con resina; vilano uniseriado, con uno o en ocasiones dos tipos de elementos, rara vez biseriado o multiseriado, pero con un solo tipo de elementos, a veces ausente ……….…………………..……..………...……….. 21
21a Cipselas sin fitomelanina, generalmente pardas o pajizas ………………….……….. 22
21b Cipselas con fitomelanina, negras ……...…………………………….………………. 23
22a Cabezuelas agrupadas en inflorescencias; involucro uniseriado, con o sin calículo; filarios iguales; anteras con collar cilíndrico o balaustriforme; vilano uni o multiseriado, persistente o deciduo, de cerdas capilares barbeladas o glabras ……………………………...………………………………….………….. Senecioneae
22b Cabezuelas solitarias; involucro multiseriado, sin calículo; filarios graduados; anteras sin collar; vilano uni o biseriado, persistente, de escamas ………………….. Arctotideae
23a Filarios naviculares; flores centrales con corolas 4(-5) lobuladas en el ápice; vilano de 2(1 o 4) cerdas, y una corona de escamas fusionadas, rara vez de escuámulas vestigiales o ausente ………………………………………….……………………….… Perityleae
23b Filarios laminares; flores centrales con corolas 5 lobuladas; vilano de escamas con o sin una vena media engrosada, a veces proyectándose en una arista, o escamas aristadas, libres.
25a Involucro campanulado a hemisférico; filarios herbáceos; receptáculo globoso, a veces plano; flores radiadas trilobuladas en el ápice; ramas del estilo truncadas, pilosas en el ápice; anteras con collar cilíndrico ……………………………….…………. Helenieae
25b Involucro cilíndrico o turbinado, a veces hemisférico; filarios membranáceos, receptáculo plano o ligeramente convexo; flores radiadas unilobuladas en el ápice; ramas del estilo lanceoladas, agudas, papilosas por arriba del punto de bifurcación; anteras sin collar …………………………………….……………………………………... Bahieae
26a Involucro uni o biseriado; filarios con glándulas estipitadas; páleas asociadas solo con las flores periféricas; anteras moradas …………………………………………. Madieae
26b Involucro multiseriado, rara vez uni o biseriado; filarios pilosos o con glándulas sésiles; páleas dispuestas en todo el receptáculo, a veces ausentes en las flores periféricas, anteras negras o blanquecinas ……………………………………………………………...…. 27
27a Filarios escariosos, excepto en la parte media, con glándulas sésiles; ramas del estilo truncadas, pilosas solo en el ápice; anteras blanquecinas con collar balaustriforme; cipselas con glándulas sésiles…………………………………………… Anthemideae
27b Filarios herbáceos o membranáceos, a veces escariosos en el margen, pilosos o glabros, rara vez con glándulas sésiles; ramas del estilo lanceoladas o peniciladas, agudas, pilosas o papilosas por arriba del punto de bifurcación, a veces glabras; anteras negras con collar cilíndrico o ausente; cipselas glabrescentes o pilosas ……..………………. 28
28a Filarios herbáceos, no escariosos; páleas laminares o naviculares, a veces deciduas como unidad o formando un complejo con las cipselas y los filarios; flores centrales bisexuales o funcionalmente masculinas, lóbulos papilosos; anteras sin collar; cipselas obcónicas, obpiramidales u obovoides, a veces esféricas, glabras, estriadas o tuberculadas ……………………………………………………………………...…….….. Millerieae
28b Filarios membranáceos, escariosos en el margen; páleas conduplicadas, persistentes, sin formar complejos con las cipselas, rara vez ausentes; flores centrales bisexuales, lóbulos glabros o glandulares; anteras con collar cilíndrico; cipselas teretes o prismáticas, costilladas, pilosas o glabrescentes, a veces estriadas …………………… Neurolaeneae
Conclusiones
Por primera vez se presentan las características de las cabezuelas para las 26 tribus de Asteraceae que se distribuyen en México, además de los órganos asociados a ellas, como son: involucro, filarios, receptáculo, páleas, flores periféricas, flores centrales, androceo, estilo, cipselas y vilano. Dichas estructuras están descritas de manera sencilla e ilustradas con fotografías de material in vivo, herborizado o conservado en espíritu, destacando los caracteres diagnósticos de cada tribu, la morfología más común, así como las excepciones que ocurren en algunos de sus miembros. Esto, con la finalidad de brindar al lector un panorama general de la morfología de las asteráceas y facilitar el uso de la clave de identificación de las tribus nativas e introducidas presentes en el país. Aunque Asteraceae es uno de los grupos más fáciles de identificar a nivel familia, en categorías taxonómicas inferiores la situación cambia completamente, ya sea porque las claves son antiguas y solo están disponibles en literatura especializada de difícil acceso; son poco conocidas o las personas que las consultas no están familiarizadas con las características morfológicas y terminología empleadas en ellas.
Adicionalmente, se incluye una actualización del número de especies y cambios nomenclaturales ocurridos después de la publicación del catálogo de plantas vasculares nativas de México (Villaseñor, 2016) y la estimación de la diversidad de asteráceas mexicanas (Villaseñor, 2018). A pesar de que Asteraceae es una de las familias más estudiadas en el mundo y México, uno de los principales centros de diversidad y endemismo, aún es necesario realizar mucho trabajo florístico y taxonómico relacionado con este grupo de angiospermas. Por lo anterior, esta contribución servirá de referencia a taxónomos, estudiantes de biología y áreas afines que requieran conocer y aprender la morfología básica de las compuestas e identificar especímenes a nivel tribal.

